










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Successional stage, fragmentation and exposure to extraction influence the population structure of Euterpe precatoria (Arecaeae)
Influencia de la etapa de sucesión, fragmentación y exposición a la extracción en la estructura poblacional de Euterpe precatoria (Arecaeae)
Influencia de la etapa de sucesión, fragmentación y exposición a la extracción en la estructura poblacional de Euterpe precatoria (Arecaeae)
*Dirección para correspondencia:
Abstract
The neotropical palm Euterpe precatoria is subject to extraction for its valuable palm heart. The development of management and conservation practices for this species requires understanding of its population structure, dynamics, and traditional use across the range of environments it inhabits, from different successional stages in continuous forest to forest fragments. Here, we analyzed how the population structure of E. precatoria varies with successional stage, fragmentation, and exposure to extraction. Since E. precatoria recruitment increases with disturbance, we expected seedling density to be higher in secondary forests and fragments relative to primary forests. The study was conducted from 2007-2008 in the Caribbean Slope of Costa Rica at Braulio Carrillo National Park (BCNP), La Selva Biological Station (LSBS), Manú Center, and Finca El Progreso (FEP). The first two sites had continuous primary and secondary forests (BCNP had one extracted primary forest); the last two consisted of primary forest fragments. Population structure was variable, with greater densities in the extracted primary forest, and in the secondary forests, as compared to primary forests and fragments. Palms <5m across all sites represented 50-90% of the total number of individuals. In sites that suffered historical over-extraction, local communities have lost the tradition of consuming this species. Understanding how population dynamics is affected by extraction and succession is essential to the design of sustainable management programs rooted in community participation.
Key words: Euterpe precatoria, non-timber forest products, palm conservation, palm ecology, palm management.
Resumen
La palma neotropical Euterpe precatoria sufre un proceso de extracción ilegal debido al sabor y calidad de su palmito. El desarrollo de prácticas de manejo y conservación de esta especie requiere de la comprensión de su estructura y dinámica poblacional en los diferentes ambientes que coloniza. Analizamos cómo la estructura poblacional de E. precatoria varió con el estadio sucesional, la fragmentación y la exposición a la extracción. Esperábamos que la densidad de plántulas fuera mayor en bosques secundarios y fragmentos en relación con los bosques primarios. El estudio se realizó entre 2007-2008 en la vertiente del Caribe de Costa Rica en el Parque Nacional Braulio Carrillo, la Estación Biológica La Selva, el Centro Manú y la Finca El Progreso. Los dos primeros sitios tenían bosques primarios y secundarios continuos (Braulio Carrillo tenía además un bosque primario extraído), mientras que los dos últimos representaban fragmentos de bosques primarios con una historia previa de extracción. La mayor densidad de plántulas se encontró en el bosque primario extraído y en los bosques secundarios, mientras que las palmas reproductivas fueron más comunes en los bosques primarios y en los fragmentos. Las palmas <5m en todos los sitios representaron el 50-90% del número total de individuos. En los sitios que históricamente sufrieron sobre-extracción, las comunidades perdieron la tradición de consumir esta especie. Comprender los factores que afectan la estructura poblacional es esencial para el diseño de programas de gestión sostenible basados en la participación comunitaria.
Palabras clave: Euterpe precatoria, productos forestales no maderables, conservación de palmas, ecología de palmas, palmito, manejo de palmas.
The intensification of the use of non-timber forest products over the last half century has increased the extent of human impacts on tropical ecosystems, thereby threatening the viability of many wild populations (Cunningham & Milton 1987, Orlande et al. 1995). In many cases, resources are over-extracted surpassing the recovery capacity of the target species (Hall & Bawa 1993, Ticktin 2004). This is particularly true for palms, which represent one of the most economically important groups of Angiosperms in tropical forests (Johnson 1996, Campos & Ehringhaus 2003, Dransfield et al. 2008). Arecaceae has been a traditional source of construction materials, food, and fiber for human groups (Macía et al. 2011). Palm use is intrinsically linked to local knowledge, passed from generation to generation, but this age-old human-palm relationship is changing dramatically due to large scale commercial trade (Kahn 1991, Morcote-Ríos & Bernal 2001, Campos & Ehringhaus 2003, Manzi & Coomes 2008, Weinstein & Moegenburg 2004), which is influencing the population structure and geographic distribution of many species through cultivation, extraction, and fragmentation (Bonesso-Sampaio et al. 2007, Endress et al. 2004, Flores & Ashton 2000).
Intensive palm extraction is linked to habitat fragmentation; the combination of both factors cause significant changes in population structure and genetic diversity. Sezen et al. (2007) showed that fragmentation and changes in land uses significantly reduced the genetic diversity of seedlings and saplings of Iriartea deltoidea in second growth forests in the vicinity of La Selva, Costa Rica. Harvesting of palm heart from I. deltoidea and similar species usually targets the largest individuals, which produce the most palm heart. The harvesting of specific size classes significantly determines population growth and tolerance to extraction (Freckleton et al. 2003).
In Costa Rica, the core of illegal palm heart harvesting is concentrated on Euterpe precatoria Mart. Park rangers estimate that around 2 000 palms per year are illegally extracted from Braulio Carrillo National Park (Avalos 2007). A typical extraction bout lasting three days could remove more than 300 individuals close to 20m in height (Avalos 2007). This practice started as a cultural Easter tradition involving the extraction of a handful of stems per household, but now has evolved into an important underground commercial activity feeding local black markets year round (Sylvester & Avalos 2009). The extraction of palm heart (notwithstanding its low nutritional value, Mora-Urpí et al. 1997) has continued throughout much of the region (even though it has been banned by local laws, and consumers have the alternative choice of using commercially-produced palm heart from Bactris gasipaes). The consumption of E. precatoria is centered on Costa Rican Holy Week celebrations, corresponding with intensified extraction activities. As the land was divided into pastures and fields, access to standing forest became more limited, and wild palms (whose populations had been depleted by extraction and deforestation) became more difficult to find. Thus, people began replacing wild palm heart with B. gasipaes. Today the commercial and domestic cultivation of B. gasipaes is ubiquitous in many rural communities while the tradition of using wild palm heart has been abandoned. A decline in traditional extraction has been observed in lowland rainforest communities on the Caribbean slope of Costa Rica. In communities at higher elevations, another culturally significant palm, Geonoma undata subsp. edulis, has become exceedingly inaccessible as its palm heart is now provided by local poachers (Sylvester & Avalos 2009). Poachers, as opposed to families, now exert the majority of extraction pressure on wild palms, and the overexploitation caused by this activity threatens forest diversity and function, as well as the survival of inherently valuable traditions.
Here, we pursue to answer the question: how does the population structure of E. precatoria change with successional stage, forest fragmentation, and exposure to extraction? We predict that seedling density increases with disturbance, being higher in secondary forests, extracted forests, and forest fragments relative to primary forests, where adults are expected to be proportionally more abundant. Understanding the factors that influence population structure is critical to secure the long-term viability of this species, improve its current conservation status and management practices, and maintain its cultural significance.
Materials and Methods
Study sites: This study was conducted during April, August and November from 2007 to 2008 at four sites along the Caribbean Slope of Costa Rica: Braulio Carrillo National Park (BCNP), La Selva Biological Station (LSBS), Manú Center (Manú) and Finca El Progreso (FEP). The first two sites have continuous primary and secondary forests, whereas the latter two consisted of fragments maintaining primary forests. In BCNP, data were collected at Quebrada González Station (48 000Ha, 10º09’44’’ N - 83º56’15’’ W, 400-500m), a tropical wet premontane forest. Here, we selected three areas representing different successional stages and disturbance regimes: BCNP primary (a primary forest without extraction), BCNP secondary (a secondary forest without extraction), and BCNP extracted (a primary forest that suffered recent extraction). At LSBS (1 600Ha, 10º25’58’’ N - 84º00’06’’ W, 30-150m) data were obtained from primary (LSBS primary) and secondary forests (LSBS secondary), both without contemporary records of palm heart extraction. The fragmented forest of Manú (10º09’29’’ N - 83º46’57’’ W, 450m, Buenos Aires de Guápiles) consisted of 12ha of tropical premontane rainforest receiving 4 475mm of annual precipitation (IICA 1971). The surrounding area is a mosaic of lightly disturbed old-growth forests, and selectively logged old-growth scrub and pastures, as well as residential lots. The fragment at FEP (10º30’35’’ N - 83º44’39’’ W, Cariari de Guápiles) is located at 45m with an annual precipitation of 4 000-5 000mm. This 30ha tropical humid rainforest fragment is found within a similar landscape of lightly disturbed old-growth forests, selectively logged old-growth forests, timber plantations, pastures, and agricultural land.
Manú and FEP were previously utilized for selective logging, but have been left undisturbed since 1996 and 1987, respectively. Historically, palm heart extraction took place with high intensity at both sites but has decreased in the last decade, likely due to local reduction in abundance of E. precatoria.
Study species: Euterpe precatoria Mart. belongs to the subfamily Arecoideae and the tribe Euterpeinae along with five other genera (Prestoea, Neonicholsonia, Hyospathe, Oenocarpus and Jessenia). Euterpe precatoria is divided into two varieties by Henderson (1995), longevaginata (stems solitary or cespitose, low and high elevations in the Andes and Central America) and precatoria (stems solitary, low elevations in the Amazon). In Costa Rica, E. precatoria var longevaginata is commonly described as a single-stemmed, solitary subcanopy palm distributed from sea level to 1 150m of elevation from Belize to Bolivia (Grayum 2003, Henderson et al. 1995). The vertical stem may reach 26m in height with a diameter at breast height (DBH) of up to 15cm, and a cone of stilt roots that in extreme cases may reach over 1.5m in height. This monoecious species has protandrous inflorescences, is pollinated by beetles and bees (Küchmeister et al. 1997), and is dispersed by birds (Galetti & Aleixo 1998, Henderson 2002), and by Cebus monkeys in Colombia (Zona & Henderson 1979). Across its latitudinal range, E. precatoria is found in inundated forests (where it reaches high densities), as well as in drained terra firme forests (Henderson 1995). In Costa Rica, the overall density of E. precatoria tends to be lower relative to what has been recorded in the Western Amazon, occurring in highly diverse lowland as well as premontane forests (Clark et al. 1995, Homeier et al. 2002, Vormisto et al. 2004). It colonizes a variety of substrata including fallen logs and the buttresses of dead canopy trees. Because it is a monopodial species, the extraction of its palm heart results in the death of the individual. The disturbance created by the extraction process favors seedling and juvenile growth, which increases the overall abundance of E. precatoria in extracted sites (Avalos 2007). This is consistent with our observations, which show that this species benefits from moderate forest disturbance and intermediate light levels.
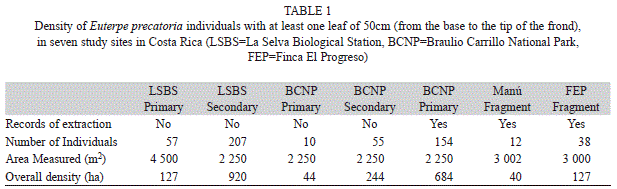
Population structure determination: We used square plots to quantify the population structure of E. precatoria at each site. Plot establishment was as follows: - In BCNP we made 30 permanent plots of 15x15m (10 plots in primary, secondary and extracted forest, respectively). Plots were separated by at least 50m. - At LSBS we made 20 plots (15x15m) in primary forest, and 10 plots (15x15m) in secondary forest. - Within the forest fragments of Manú we made 10 plots (5x5m), and 43 plots (8x8m) located at intervals of 15m. - Finally, at FEP, we established 40 plots (5x5m), and 20 plots (10x10m). Because of the need to avoid edge effects, the shape and size of the forest fragments prevented the implementation of an identical sampling protocol relative to the continuous forests. Plot size varied in fragments in order to include a representative sample of the habitat heterogeneity prevalent at each site, while simultaneously maintaining a distance of at least 50m from the nearest forest edge. However, sampled areas across sites were comparable and representative of population abundance and habitat heterogeneity at each location (Table 1).
Within each plot, all E. precatoria individuals with at least one leaf of 50cm (from the base to the tip of the frond) were located and measured. At Manú and FEP we found very low abundances of palms with at least one leaf ≥50m. At these sites, we also counted the number of seedlings with leaves <50cm to determine the regeneration potential of E. precatoria in fragmented forest conditions. We measured stem height from the ground to the first frond (excluding the crown shaft) using basic trigonometry rules, and a Suunto PM-5 clinometer in palms higher than 8m. Stem height was corrected by subtracting the height of the cone of stilt roots.
Palm-use history: To establish the history of palm use as a forest resource, we conducted semi-structured interviews with members of the Buenos Aires (Manú) and Colinas (FEP) communities, with each household representing a sample unit. The purpose of these interviews was to gather information on previous land-use of these sites and the surrounding areas as well as the history of palm extraction. Questions gathered information on demographics (gender, age, family composition, time residing in area), history of farm and forest fragments in the area (land use history, forest use history), and palm use and extraction habits (what palm products were extracted and from what species, where and how were they extracting, what were the main motivation for extraction). We identified 10 people per site who were key community members with sufficient knowledge on the history of land use at each site (Buenos Aires community -Manú- and FEP and vicinity).
Palm population structure was compared among sites by determining the distribution of individuals in each height class. We used correspondence analysis to compare the distribution of E. precatoria across sites in terms of the similarity in abundance of different ontogenetic stages. E. precatoria undergoes an ontogenetic transition at about 1m in stem height and reaches maturity at 10m stem height (Avalos & Fernández Otárola 2010). Thus, we considered individuals <1m as seedlings, those between 1 and 10m as juveniles, and palms >10m as adults.
Results
Palm density: Considerable differences in absolute abundance of E. precatoria were found among sites. Palm density varied almost two orders of magnitude among studied sites (920 individuals per Ha at LSBS secondary forest compared with 36 individuals per Ha in the primary forest of BCNP, Table 1). Greater concentrations of E. precatoria were found in primary and secondary continuous forests compared to the primary forest fragments of Manú and FEP. Palm concentration in sites exposed to frequent extraction (BCNP extracted forest) showed the highest density.
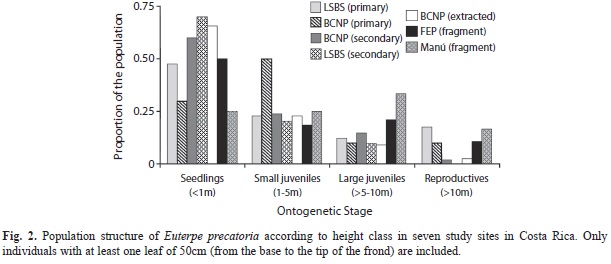
Population structure: The population structure of E. precatoria varied among sites. There was a lack of tall palms (>10m) in secondary forests (LSBS and BCNP) as well as in the extracted forest of BCNP. The primary forest fragments of Manú and FEP exhibited similar trends as those of primary forests without extraction at LSBS and BCNP, both having a lower proportion of seedlings and more reproductive individuals. This pattern is reflected in the ordination diagram generated by the correspondence analysis, where disturbed areas formed a group clustered around the abundance of seedlings and small juveniles. Primary forests and fragments show similar compositions related to the abundances of large individuals (Fig. 1). Euterpe precatoria was predominantly represented by individuals <5m across all study sites. Combined proportions of individuals <5m varied between 50-90% of the total number of individuals (Fig. 2). The inclusion of very short individuals (leaf length <50cm) in the populations of Manú and FEP accounted for 92 and 87% of these populations respectively.
Palm-use: Ninety percent of participants had lived in their respective communities for more than 10 years. An equal number of men and women were interviewed, ranging from 17-64 years old with an average age of 44. The average family size varied between three and five members.
Interviews revealed a tendency to consume commercially-produced palm heart from Bactris gasipaes (locally known as “pejibaye”) over wild palm heart harvested from the surrounding forests. Families who had consumed E. precatoria (locally known as “palmito mantequilla”) referred to past experiences (>10 years) when palm heart was not commercially available and the forest was closer to their homes. In contrast to areas of Costa Rica where wild populations of E. precatoria are relatively available (see Sylvester & Avalos 2009), the traditional use of palm heart from this species has been abandoned. No recent accounts of E. precatoria harvest/consumption were provided in the interviews. Nevertheless, selected interviewees in both sites referenced poaching activity in their communities between 2005-2010 (but of unknown species).
Discussion
Habitat disturbance increases E. precatoria abundance: Palm recruitment and establishment is affected by increased light and nutrient availability (Chazdon 1986, Everham et al. 1996), as well as by the palm dispersion strategy (Cintra & Horna 1997, Fragoso et al. 2003), previous history of land use practices (Clark et al. 1995, Sezen et al. 2007) and human intervention (Wright & Duber 2001). For many palm species, increased disturbance favoring recruitment and palm growth is associated with changes in canopy structure (Cintra & Horna 1997, De Steven 1989, Svenning 1999, Terborgh & Davenport 2001), even if light increases are small (Svenning 2000). In this study, E. precatoria preferentially recruited in slightly disturbed environments such as secondary and extracted forests. The density of this species increased with the magnitude of disturbance, as reflected by the steep rise in seedling number with increasing disturbance regime. However, E. precatoria was also prevalent in primary forests, showing that this species is tolerant enough to survive under shade and could regenerate with progressing forest succession and canopy closure. Poorter (1999) has shown that E. precatoria grows best at intermediate light levels (50% of canopy cover), which is consistent with our observations of increased growth and abundance with increasing canopy openings, such as those brought about by extraction and early successional environments. The regeneration behavior of E. precatoria falls within the concept of cryptic pioneer, since it regenerates under moderately disturbed conditions but can survive under moderate shade (Kitajima & Poorter 2008). The extraction of E. precatoria creates low intensity canopy and understory disturbance. The canopy opening created by a palm fall is small, as the crown and trunk are compact and most of the disturbance is restricted to the forest understory (Avalos 2007). While small, the increase in canopy aperture and the removal of adult palms favors an increase in the abundance of juveniles and seedlings. A dynamic disturbance regime could facilitate germination and early E. precatoria establishment.
Aguilar-Barquero & Jiménez-Hernández (2009) analyzed changes in diversity and abundance of the palm assemblage at FEP using three fragments with different degrees of human intervention (primary forest, logged primary forests left to natural regeneration, and secondary forest). Their sampling protocol spanned a smaller area for palms <5m (429m2), compared to our study. Among the 15 species of palms recorded in that study, E. precatoria was found (but with low frequency) only in primary forest fragments, showed higher density in the primary forest with some degree of human disturbance, and was intermediate in abundance relative to other palms. The authors concluded that E. precatoria could be an indicator of primary forest conditions. In our study, the preference for disturbed habitats was clear at the seedling and juvenile stages, but did not translate consistently to adult palms. Avalos (2007) found that extraction processes increase the number of individuals in lower size classes, similar to what was seen at extracted and secondary forests in BCNP. Svenning (1998) reported an analogous preference for disturbed forest in the stilt-rooted palm, Iriartea deltoidea, in lowland Amazonian Ecuador.
Forests under pressure of extraction, as well as secondary forests, did not support high densities of individuals >10m. The lack of adult palms in extracted forests is a consequence of selective extraction by poachers, who maximize the yield of palm heart by removing the tallest, reproductive individuals (Avalos 2007). This may limit the reproductive capacity of the population, and as observed, alter population structure with negative implications for population viability (Sedrez dos Reis et al. 2000). Freckleton et al. (2003) modeled the effects of a variety of harvest regimes on E. edulis and predicted that palm populations may recover following low or moderate harvesting, but that sustained high levels of harvesting targeting adults and pre-adults (so that reproductive individuals take longer to be replaced) may lead to local population extinction. Because E. precatoria has a large minimum reproductive size of 10m, and a slow growth rate compared to commercial palm heart species, its population size may require more time to recover after extensive extraction events. In areas of secondary forests, the scarcity of adult palms may represent a time-lag effect as a result of E. precatoria’s slow growth; there may not have been sufficient time post-disturbance to allow establishment of adult palms. This is why adults are more associated with primary forests.
Resilience to fragmentation effects: Euterpe precatoria is a long lived forest palm, which makes it more resilient to the extinction and thinning processes associated with fragmentation. Palm populations within recently fragmented forests, such as Manú and FEP (12 years and 21 years respectively), may not demonstrate the full effects of fragmentation -i.e., decrease in adult density-. The adults at FEP denote a time-lag effect since this area has not yet begun to express fragmentation effects. This could be related to their larger size compared to Manú. Continued monitoring of the population structure within these forest fragments would be necessary to determine how long it may take for the effects of fragmentation to be expressed as the adults begin to die.
The apparent abundance of seedlings in relation to adult palms in the studied fragments suggests that E. precatoria seeds are being dispersed into these fragments from neighboring populations. The landscape matrices in which these forest fragments exist still maintain a fair number of adults (pers. obs.), so that bird dispersal could be sufficiently effective to maintain isolated populations (Guindon 1996, Sezen et al. 2007). Beyond seedling input, preliminary processes related to fragmentation may occur. The low density and higher proportion of large juveniles and adults in forest fragments suggests that seedling recruitment rates have declined, and that mortality rates have probably increased. Preliminary data monitoring changes in canopy cover in our study fragments (using hemispherical photographs) indicate that E. precatoria seedlings under deep canopy cover show higher mortality rates and slower growth than those in forest fragments with greater canopy aperture. An increase in seedling predation or a decline in the number and quality of safe habitats (a characteristic of fragmented forests) could contribute to the reduced number of recruits. Without incoming seeds from outside sources, the effects of fragmentation and dense canopy conditions could be much more pronounced. The effects of forest fragments, such as the ones studied here, are combined with a previous history of extraction. Fragmentation is a complex process whose effects cannot be analyzed independently of previous human intervention.
Changing traditional uses of E. precatoria: The preservation and incorporation of traditional resource use in conservation strategies is important for natural resource management. Traditional activities, such as palm heart extraction, link local communities with the surrounding forests. An understanding of natural resource availability and distribution and how it may be affected by traditional use and human disturbance is essential to the design of sustainable resource management programs rooted in community participation. The situation of E. precatoria extraction has arrived at a critical juncture. With the increased remoteness of palm populations, traditional practices (i.e., low intensity extraction done sparingly by communities themselves to satisfy religious purposes) are either replaced by illegal poaching (i.e., high intensity extraction from protected areas done by professional poachers to satisfy a black market and provide palm heart all year round) or are abandoned altogether. The future of E. precatoria and the survival of the important cultural activity of palm harvesting are interdependent. Outside of national parks and protected areas, few alternatives exist to absorb extraction pressures. Conservation policies must consider programs to promote to moderate intensity extraction, such as cultivation in fragments owned by local communities, or even the introduction of alternative uses that might be more profitable than short-term, destructive harvesting, such as the utilization of E. precatoria fruits, which have significant antioxidant capacity (Kang et al. 2012) and represent a major source of income for human communities in the Amazon basin (Stoian 2004, Macía et al. 2011).
Acknowledgments
The authors thank the Alpízar family at Finca El Progreso, and the staff of Manú Center for facilitating field work within these forest fragments. Students and staff of The School for Field Studies helped during fieldwork. The park rangers of Quebrada González (C. Mora, P. Ezeta-Salicetti and S. Barquero) and the staff of La Selva provided significant logistical support. M.F.O. was supported by the REU Scholarship Program of the Organization for Tropical Studies (OTS) with CRUSA Foundation for Cooperation financial support and by an OTS Research Fellowship (Don and Beverly Stone Fund). Comments by Henrik Balslev and an anonymous reviewer significantly improved the manuscript. This research was supported by the Directorate of Research of the University of Costa Rica project number 111-A3-129 and The School for Field Studies. Ideawild provided part of the field equipment.
References
Aguilar-Barquero, V. & F. Jiménez-Hernández. 2009. Diversidad y distribución de palmas (Arecaceae) en tres fragmentos de bosque muy húmedo en Costa Rica. Rev. Biol. Trop. 57: 83-92. [ Links ]
Avalos, G. 2007. Changes in size preference of illegally extracted heart of palm Euterpe precatoria (Arecaceae) in Braulio Carrillo National Park Costa Rica. Econ. Bot. 61: 96-98. [ Links ]
Avalos, G. & M. Fernández Otárola. 2010. Allometry and stilt root structure of the neotropical palm Euterpe precatoria (Arecaceae) across sites and successional stages. Am. J. Bot. 97: 388-394. [ Links ]
Bonesso-Sampaio, M., I.B. Schmidt & I.B. Figueiredo. 2007. Harvesting effects and population ecology of the Buriti Palm (Mauritia flexuosa L. f. Arecaceae) in the Jalapão Region Central Brazil. Econ. Bot. 62: 171-181. [ Links ]
Campos, M.T. & C. Ehringhaus. 2003. Plant virtues are in the eyes of the beholders: a comparison of known palm use among indigenous and folk communities of southwestern Amazonia. Econ. Bot. 57: 324-344. [ Links ]
Chazdon, R.L. 1986. Light variation and carbon gain in rain forest understory palms. J. Ecol. 74: 995-1012. [ Links ]
Cintra, R. & V. Horna. 1997. Seed and seedling survival of the palm Astrocaryum murumuru and the legume tree Dipteryx micrantha in gaps in Amazonian forest. J. Trop. Ecol. 13: 257-277. [ Links ]
Clark, D.A., D.B. Clark, R.M. Sandoval & M.V.C. Castro. 1995. Edaphic and human effects on landscape-scale distributions of tropical rain forest palms. Ecology 76: 1581-1594. [ Links ]
Cunningham, A.B. & S.J. Milton. 1987. Effects of basket-weaving industry on Mokola Palm and dye plants in Northwestern Botswana. Econ. Bot. 41: 386-402. [ Links ]
Dransfield, J.N., W. Uhl, C.B. Amussen, W.H.J. Baker, M.M. Harley & C.E. Lewis. 2008. Genera Palmarum: The Evolution and Classification of Palms. Kew Publishing Royal Botanic Gardens, London, United Kingdom. [ Links ]
De Steven, D. 1989. Genet and ramet demography of Oenocarpus mapora ssp. mapora a clonal palm of Panamanian tropical moist forest. J. Ecol. 77: 579-596. [ Links ]
Endress, B.A., D.L. Gorchov & R.B. Noble. 2004. Non-timber forest product extraction: effects of harvest and browsing on an understory palm. Ecol. Appl. 14: 1139-1153. [ Links ]
Everham, E.M., R.W. Myster & E. Vandegenachte. 1996. Effects of light moisture temperature and litter on the regeneration of five tree species in the tropical montane wet forest of Puerto Rico. Am. J. Bot. 83: 1063-1068. [ Links ]
Flores, C.F. & P.M.S. Ashton. 2000. Harvesting impact and economic value of Geonoma deversa Arecaceae an understory palm. Econ. Bot. 54: 267-277. [ Links ]
Fragoso, J.M.V., K.M. Silvius & J.A. Correa. 2003. Long-distance seed dispersal by tapirs increases seed survival and aggregates tropical trees. Ecology 84: 1998-2006. [ Links ]
Freckleton, R.P., D.M. Silva-Matos, M.L.A. Bovi & A.R. Watkinson. 2003. Predicting the impacts of harvesting using structured populations models: the importance of density-dependence and timing of harvest for a tropical palm. J. Appl. Ecol. 40: 846-858. [ Links ]
Galetti, M. & A. Aleixo. 1998. Effects of palm heart harvesting on avian frugivores in the Atlantic rain forest of Brazil. J. Appl. Ecol. 35: 286-293. [ Links ]
Grayum, M.H. 2003. Arecaceae, p. 201-293. In B.E. Hammel, M.H. Grayum, C. Herrera & N. Zamora (eds.). Manual de Plantas de Costa Rica. Vol. III. Missouri Botanical Garden, St. Louis, Missouri, USA. [ Links ]
Guindon, C.F. 1996. The importance of forest fragments to the maintenance of regional biodiversity in Costa Rica, p. 168-186. In J. Schenlhas & R. Greenberg (eds.). Forest Patches in Tropical Landscapes. Island, Washington D.C., USA. [ Links ]
Hall, P. & K. Bawa. 1993. Methods to assess the impact of extraction of non-timber tropical forest products on plant populations. Econ. Bot. 47: 234-247. [ Links ]
Henderson, A. 1995. The Palms of the Amazon. Oxford University, New York, USA. [ Links ]
Henderson, A., G. Galeano & R. Bernal. 1995. Field guide to the palms of the Americas. Princeton University, New Jersey, USA. [ Links ]
Henderson, A.G. 2002. Evolution and ecology of palms. New York Botanical Garden, New York, USA. [ Links ]
Homeier, J., S.W. Breckle, H. Dalitz, C. Levers & R. Ortiz. 2002. Demography, spatial distribution, and growth of three arborescent palm species in a tropical premontane rain forest in Costa Rica. Ecotropica 8: 239-247. [ Links ]
IICA. 1971. Catálogo de Estaciones Agrícolas Experimentales del Istmo Centroamericano. Instituto de Investigaciones en Ciencias Agropecuarias. Publicación Miscelánea No. 83 Turrialba, Costa Rica. [ Links ]
Johnson, D. (ed.). 1996. Palms: Their Conservation and Sustained Utilization. Status Survey and Conservation Action Plan, IUCN, Gland, Switzerland and Cambridge, United Kingdom. [ Links ]
Kahn, F. 1991. Palms as key swamp forest resources in Amazonia. For. Ecol. Manag. 38: 133-142. [ Links ]
Kang, J., K.M. Thakali, X. Chenghui, M. Kondo, Y. Tong, B. Ou, G. Jensen, M.B. Medina, A.G. Schauss & X. Wu. 2012. Bioactivities of açaí (Euterpe precatoria Mart.) fruit pulp, superior antioxidant and anti-inflammatory properties to Euterpe oleracea Mart. Food Chemistry 133: 671-677. [ Links ]
Kitajima, K. & L. Poorter. 2008. Functional basis for resource niche partitioning by tropical trees, p. 160-181. In W.P. Carson & S.A. Schnitzer (eds.). Tropical Forest Community Ecology. Wiley-Blackwell, New Jersey, USA. [ Links ]
Küchmeister, H., I. Silberbauer-Gottsberger & G. Gottsberger. 1997. Flowering pollination nectar standing crop and nectaries of Euterpe precatoria (Arecaceae) an Amazonian rain forest palm. Plant Syst. Evol. 206: 71-97. [ Links ]
Macía, M.J., P.J. Armesilla, R. Cámara-Leret, N. Paniagua-Zambrana, S. Villalba, H. Balslev & M. Pardo-deSantayana. 2011. Palm Uses in Northwestern South America: A Quantitative Review. Bot. Rev. 77: 462-570. [ Links ]
Manzi, M. & O.T. Coomes. 2008. Managing Amazonian palms for community use: a case of aguaje palm (Mauritia flexuosa) in Peru. For. Ecol. Manag. 257: 510-517. [ Links ]
Mora-Urpí, J.C., J.C. Weber & C.R. Clement. 1997. Peach palm, Bactris gasipaes Kunth. Promoting the Conservation and Use of Underutilized and Neglected Crops. Institute of Plant Genetics and Crop Plant Research, Rome, Italy. [ Links ]
Morcote-Ríos, G. & R. Bernal. 2001. Remains of palms (Palmae) at archaeological sites in the New World: a Review. Bot. Rev. 67: 309-350. [ Links ]
Orlande, T., J. Laarman & J. Mortimer. 1995. Palmito sustainability and economics in Brazil’s Atlantic coastal forest. For. Ecol. Manag. 80: 257-265. [ Links ]
Poorter, L. 1999. Growth responses of 15 rainforest tree species to a light gradient: the relative importance of morphological and physiological traits. Funct. Ecol. 13: 396-410. [ Links ]
Sedrez-dos Reis, M., A.C. Fantini, R.O. Nodari, A. Reis, M.P. Guerra & A. Mantovani. 2000. Management and conservation of natural populations in Atlantic Rain Forest: the case study of palm heart (Euterpe edulis Martius). Biotropica 32: 894-902. [ Links ]
Sezen, U.U., R.L. Chazdon & K.E. Holsinger. 2007. Multigenerational genetic analysis of tropical secondary regeneration in a canopy palm. Ecology 88: 3065-3075. [ Links ]
Stoian, D. 2004. Todo lo que sube tiene que bajar: La economía del palmito (Euterpe precatoria) en el norte amazónico de Bolivia, p. 117-140. In N. Alexiades & P. Shanley (eds.). Productos Forestales, Medios de Subsistencia y Conservación. CIFOR, Bogor, Indonesia. [ Links ]
Svenning, J.C. 1998. The effects of land-use on the local distribution of palm species in an Andean rain forest fragment in northwestern Ecuador. Biodivers. Conserv. 7: 1529-1537. [ Links ]
Svenning, J.C. 1999. Recruitment of tall arborescent palms in the Yasuní National Park Amazonian Ecuador: are large treefall gaps important? J. Trop. Ecol. 15: 355-366. [ Links ]
Svenning, J.C. 2000. Small canopy gaps influence plant distributions in the rain forest understory. Biotropica 32: 252-261. [ Links ]
Sylvester, O. & G. Avalos. 2009. Illegal palm heart (Geonoma edulis) harvest in Costa Rican national parks: patterns of consumption and extraction. Econ. Bot. 63: 179-189. [ Links ]
Terborgh, J. & l. Davenport. 2001. Endogenous and exogenous control of leaf morphology in Iriartea deltoidea (Palmae). J. Trop. Ecol. 17: 695-703. [ Links ]
Ticktin, T. 2004. The ecological implications of harvesting non-timber forest products. J. Appl. Ecol. 41: 11-21. [ Links ]
Vormisto, J., J.C. Svenning, P. Hall & H. Balslev. 2004. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. J. Ecol. 92: 577-588. [ Links ]
Weinstein, S. & s. Moegenburg. 2004. Açaí palm management in the Amazon Estuary: curse for conservation or passage to plantations? Conserv. Soc. 2: 317-346. [ Links ]
Wright, S.J. & H.C. Duber. 2001. Poachers and forest fragmentation alter seed dispersal seed survival and seedling recruitment in the palm Attalea butyraceae with implications for tropical tree diversity. Biotropica 33: 583-595. [ Links ]
Zona, S. & A. Henderson. 1979. A review of animal-media-ted dispersal in palms. Selbyana 11: 6-21. [ Links ]
Avalos, G. 2007. Changes in size preference of illegally extracted heart of palm Euterpe precatoria (Arecaceae) in Braulio Carrillo National Park Costa Rica. Econ. Bot. 61: 96-98. [ Links ]
Avalos, G. & M. Fernández Otárola. 2010. Allometry and stilt root structure of the neotropical palm Euterpe precatoria (Arecaceae) across sites and successional stages. Am. J. Bot. 97: 388-394. [ Links ]
Bonesso-Sampaio, M., I.B. Schmidt & I.B. Figueiredo. 2007. Harvesting effects and population ecology of the Buriti Palm (Mauritia flexuosa L. f. Arecaceae) in the Jalapão Region Central Brazil. Econ. Bot. 62: 171-181. [ Links ]
Campos, M.T. & C. Ehringhaus. 2003. Plant virtues are in the eyes of the beholders: a comparison of known palm use among indigenous and folk communities of southwestern Amazonia. Econ. Bot. 57: 324-344. [ Links ]
Chazdon, R.L. 1986. Light variation and carbon gain in rain forest understory palms. J. Ecol. 74: 995-1012. [ Links ]
Cintra, R. & V. Horna. 1997. Seed and seedling survival of the palm Astrocaryum murumuru and the legume tree Dipteryx micrantha in gaps in Amazonian forest. J. Trop. Ecol. 13: 257-277. [ Links ]
Clark, D.A., D.B. Clark, R.M. Sandoval & M.V.C. Castro. 1995. Edaphic and human effects on landscape-scale distributions of tropical rain forest palms. Ecology 76: 1581-1594. [ Links ]
Cunningham, A.B. & S.J. Milton. 1987. Effects of basket-weaving industry on Mokola Palm and dye plants in Northwestern Botswana. Econ. Bot. 41: 386-402. [ Links ]
Dransfield, J.N., W. Uhl, C.B. Amussen, W.H.J. Baker, M.M. Harley & C.E. Lewis. 2008. Genera Palmarum: The Evolution and Classification of Palms. Kew Publishing Royal Botanic Gardens, London, United Kingdom. [ Links ]
De Steven, D. 1989. Genet and ramet demography of Oenocarpus mapora ssp. mapora a clonal palm of Panamanian tropical moist forest. J. Ecol. 77: 579-596. [ Links ]
Endress, B.A., D.L. Gorchov & R.B. Noble. 2004. Non-timber forest product extraction: effects of harvest and browsing on an understory palm. Ecol. Appl. 14: 1139-1153. [ Links ]
Everham, E.M., R.W. Myster & E. Vandegenachte. 1996. Effects of light moisture temperature and litter on the regeneration of five tree species in the tropical montane wet forest of Puerto Rico. Am. J. Bot. 83: 1063-1068. [ Links ]
Flores, C.F. & P.M.S. Ashton. 2000. Harvesting impact and economic value of Geonoma deversa Arecaceae an understory palm. Econ. Bot. 54: 267-277. [ Links ]
Fragoso, J.M.V., K.M. Silvius & J.A. Correa. 2003. Long-distance seed dispersal by tapirs increases seed survival and aggregates tropical trees. Ecology 84: 1998-2006. [ Links ]
Freckleton, R.P., D.M. Silva-Matos, M.L.A. Bovi & A.R. Watkinson. 2003. Predicting the impacts of harvesting using structured populations models: the importance of density-dependence and timing of harvest for a tropical palm. J. Appl. Ecol. 40: 846-858. [ Links ]
Galetti, M. & A. Aleixo. 1998. Effects of palm heart harvesting on avian frugivores in the Atlantic rain forest of Brazil. J. Appl. Ecol. 35: 286-293. [ Links ]
Grayum, M.H. 2003. Arecaceae, p. 201-293. In B.E. Hammel, M.H. Grayum, C. Herrera & N. Zamora (eds.). Manual de Plantas de Costa Rica. Vol. III. Missouri Botanical Garden, St. Louis, Missouri, USA. [ Links ]
Guindon, C.F. 1996. The importance of forest fragments to the maintenance of regional biodiversity in Costa Rica, p. 168-186. In J. Schenlhas & R. Greenberg (eds.). Forest Patches in Tropical Landscapes. Island, Washington D.C., USA. [ Links ]
Hall, P. & K. Bawa. 1993. Methods to assess the impact of extraction of non-timber tropical forest products on plant populations. Econ. Bot. 47: 234-247. [ Links ]
Henderson, A. 1995. The Palms of the Amazon. Oxford University, New York, USA. [ Links ]
Henderson, A., G. Galeano & R. Bernal. 1995. Field guide to the palms of the Americas. Princeton University, New Jersey, USA. [ Links ]
Henderson, A.G. 2002. Evolution and ecology of palms. New York Botanical Garden, New York, USA. [ Links ]
Homeier, J., S.W. Breckle, H. Dalitz, C. Levers & R. Ortiz. 2002. Demography, spatial distribution, and growth of three arborescent palm species in a tropical premontane rain forest in Costa Rica. Ecotropica 8: 239-247. [ Links ]
IICA. 1971. Catálogo de Estaciones Agrícolas Experimentales del Istmo Centroamericano. Instituto de Investigaciones en Ciencias Agropecuarias. Publicación Miscelánea No. 83 Turrialba, Costa Rica. [ Links ]
Johnson, D. (ed.). 1996. Palms: Their Conservation and Sustained Utilization. Status Survey and Conservation Action Plan, IUCN, Gland, Switzerland and Cambridge, United Kingdom. [ Links ]
Kahn, F. 1991. Palms as key swamp forest resources in Amazonia. For. Ecol. Manag. 38: 133-142. [ Links ]
Kang, J., K.M. Thakali, X. Chenghui, M. Kondo, Y. Tong, B. Ou, G. Jensen, M.B. Medina, A.G. Schauss & X. Wu. 2012. Bioactivities of açaí (Euterpe precatoria Mart.) fruit pulp, superior antioxidant and anti-inflammatory properties to Euterpe oleracea Mart. Food Chemistry 133: 671-677. [ Links ]
Kitajima, K. & L. Poorter. 2008. Functional basis for resource niche partitioning by tropical trees, p. 160-181. In W.P. Carson & S.A. Schnitzer (eds.). Tropical Forest Community Ecology. Wiley-Blackwell, New Jersey, USA. [ Links ]
Küchmeister, H., I. Silberbauer-Gottsberger & G. Gottsberger. 1997. Flowering pollination nectar standing crop and nectaries of Euterpe precatoria (Arecaceae) an Amazonian rain forest palm. Plant Syst. Evol. 206: 71-97. [ Links ]
Macía, M.J., P.J. Armesilla, R. Cámara-Leret, N. Paniagua-Zambrana, S. Villalba, H. Balslev & M. Pardo-deSantayana. 2011. Palm Uses in Northwestern South America: A Quantitative Review. Bot. Rev. 77: 462-570. [ Links ]
Manzi, M. & O.T. Coomes. 2008. Managing Amazonian palms for community use: a case of aguaje palm (Mauritia flexuosa) in Peru. For. Ecol. Manag. 257: 510-517. [ Links ]
Mora-Urpí, J.C., J.C. Weber & C.R. Clement. 1997. Peach palm, Bactris gasipaes Kunth. Promoting the Conservation and Use of Underutilized and Neglected Crops. Institute of Plant Genetics and Crop Plant Research, Rome, Italy. [ Links ]
Morcote-Ríos, G. & R. Bernal. 2001. Remains of palms (Palmae) at archaeological sites in the New World: a Review. Bot. Rev. 67: 309-350. [ Links ]
Orlande, T., J. Laarman & J. Mortimer. 1995. Palmito sustainability and economics in Brazil’s Atlantic coastal forest. For. Ecol. Manag. 80: 257-265. [ Links ]
Poorter, L. 1999. Growth responses of 15 rainforest tree species to a light gradient: the relative importance of morphological and physiological traits. Funct. Ecol. 13: 396-410. [ Links ]
Sedrez-dos Reis, M., A.C. Fantini, R.O. Nodari, A. Reis, M.P. Guerra & A. Mantovani. 2000. Management and conservation of natural populations in Atlantic Rain Forest: the case study of palm heart (Euterpe edulis Martius). Biotropica 32: 894-902. [ Links ]
Sezen, U.U., R.L. Chazdon & K.E. Holsinger. 2007. Multigenerational genetic analysis of tropical secondary regeneration in a canopy palm. Ecology 88: 3065-3075. [ Links ]
Stoian, D. 2004. Todo lo que sube tiene que bajar: La economía del palmito (Euterpe precatoria) en el norte amazónico de Bolivia, p. 117-140. In N. Alexiades & P. Shanley (eds.). Productos Forestales, Medios de Subsistencia y Conservación. CIFOR, Bogor, Indonesia. [ Links ]
Svenning, J.C. 1998. The effects of land-use on the local distribution of palm species in an Andean rain forest fragment in northwestern Ecuador. Biodivers. Conserv. 7: 1529-1537. [ Links ]
Svenning, J.C. 1999. Recruitment of tall arborescent palms in the Yasuní National Park Amazonian Ecuador: are large treefall gaps important? J. Trop. Ecol. 15: 355-366. [ Links ]
Svenning, J.C. 2000. Small canopy gaps influence plant distributions in the rain forest understory. Biotropica 32: 252-261. [ Links ]
Sylvester, O. & G. Avalos. 2009. Illegal palm heart (Geonoma edulis) harvest in Costa Rican national parks: patterns of consumption and extraction. Econ. Bot. 63: 179-189. [ Links ]
Terborgh, J. & l. Davenport. 2001. Endogenous and exogenous control of leaf morphology in Iriartea deltoidea (Palmae). J. Trop. Ecol. 17: 695-703. [ Links ]
Ticktin, T. 2004. The ecological implications of harvesting non-timber forest products. J. Appl. Ecol. 41: 11-21. [ Links ]
Vormisto, J., J.C. Svenning, P. Hall & H. Balslev. 2004. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. J. Ecol. 92: 577-588. [ Links ]
Weinstein, S. & s. Moegenburg. 2004. Açaí palm management in the Amazon Estuary: curse for conservation or passage to plantations? Conserv. Soc. 2: 317-346. [ Links ]
Wright, S.J. & H.C. Duber. 2001. Poachers and forest fragmentation alter seed dispersal seed survival and seedling recruitment in the palm Attalea butyraceae with implications for tropical tree diversity. Biotropica 33: 583-595. [ Links ]
Zona, S. & A. Henderson. 1979. A review of animal-media-ted dispersal in palms. Selbyana 11: 6-21. [ Links ]
*Correspondencia a:
Gerardo Avalos. Escuela de Biología, Universidad de Costa Rica, 2060 San Pedro, San José, Costa Rica. The School for Field Studies, Center for Sustainable Development Studies, 100 Cummings Center Suite 534-G, Beverly, Massachusetts 01915-6239; avalos@fieldstudies.org
Mauricio Fernández Otárola. Escuela de Biología, Universidad de Costa Rica, 2060 San Pedro, San José, Costa Rica. Programa de Pós-graduação em Ecologia, Instituto de Biologia, Universidade Estadual de Campinas (Unicamp) 13083-970, CP 6109, Campinas, São Paulo, Brazil; maufero@gmail.com
James Theodore Engeln. Macalester College, Saint Paul, MN 55105 USA; james.engeln@gmail.com
1. Escuela de Biología, Universidad de Costa Rica, 2060 San Pedro, San José, Costa Rica.
2. The School for Field Studies, Center for Sustainable Development Studies, 100 Cummings Center Suite 534-G, Beverly, Massachusetts 01915-6239; avalos@fieldstudies.org
3. Programa de Pós-graduação em Ecologia, Instituto de Biologia, Universidade Estadual de Campinas (Unicamp) 13083-970, CP 6109, Campinas, São Paulo, Brazil; Correspondence author: maufero@gmail.com
4. Macalester College, Saint Paul, MN 55105 USA; james.engeln@gmail.com
Received 04-VI-2012. Corrected 20-X-2012. Accepted 15-XI-2012.


{kind=link}
{kind=link}