










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.61 n.3 San José Sep. 2013
Crouania pumila sp. nov. (Callithamniaceae: Rhodophyta), a new species of marine red algae from the Seaflower International Biosphere Reserve, Caribbean Colombia
Crouania pumila sp. nov. (Callithamniaceae: Rhodophyta), una nueva especie de alga roja marina de la Reserva Internacional de la Biosfera Seaflower, Caribe colombiano
Crouania pumila sp. nov. (Callithamniaceae: Rhodophyta), una nueva especie de alga roja marina de la Reserva Internacional de la Biosfera Seaflower, Caribe colombiano
*Dirección para correspondencia:
Abstract
In the Colombian Caribbean, the marine macroalgal flora of the Seaflower International Biosphere Reserve has been little studied, despite its ecological importance. Historical records have reported only 201 macroalgae species within its area of almost 350 000km2. However, recent surveys have shown a diversity of small algae previously overlooked. With the aim to determine the macroalgal diversity in the Reserve, we undertook field surveys in different ecosystems: coral reefs, seagrass beds, and rocky and sandy substrates, at different depths, from intertidal to 37m. During these field surveys, we collected a small described species belonging to the genus Crouania (Callithamniaceae, Rhodophyta), Crouania pumila sp. nov. that is decribed in this paper. This new species was distinguished from other species of the genus by a distinctive suite of traits including its diminutive size (to only 3.5mm in length), its decumbent, slightly calcified habit (epiphytic on other algae), its ramisympodial branching, the ecorticate main axes, and the elongate shape of the terminal cells of the cortical filaments. The observations were provided for both female (cystocarpic) and tetrasporangiate thalli; however, male thalli were not seen. Further studies have to be undertaken in this Reserve in order to carry out other macroalgal analysis and descriptions.
Key words: Caribbean, Colombia, Crouania pumila, new species, Old Providence Island, red algae.
Resumen
Crouania pumila sp. nov. (Ceramiaceae, Rhodophyta) es descrita para la isla de Providencia, en el Caribe colombiano. La nueva especie se destaca de las otras especies del género por características como su tamaño diminuto (hasta 3.5mm de largo), su hábito decumbente y poco calcificado (epífita sobre otras algas), su ramificación ramisimpodial, la ausencia de corticación y la forma alargada de las células terminales de los filamentos corticales. Se proveen observaciones sobre talos femeninos (cistocárpicos) y tetraspóricos. No se observaron talos masculinos.
Palabras clave: Caribe, Colombia, Crouania pumila, nueva especie, isla de Providencia.
Seaweeds are widely distributed and support most oceanic ecosystems, and generally have small sizes (Norton et al. 1996). They are at the base of the food web and, along with phytoplankton, constitute the primary producers in the ocean.
Crouania J. Agardh is a genus of filamentous terete red algae characterized by a uniaxial organization of its indeterminate axes, each axial cell bearing a whorl of three determinate branchlets, and new indeterminate axes (branches) arising from an axial cell or from the basal cell of a whorl-branchlet (Wollaston & Womersley 1998). The primordium of a new indeterminate branch appears as a chain of 10-20 cells before the formation of its own whorl-branchlets. Tetrasporangia are borne on the basal cells of whorl-branchlets, and gland cells are absent. Characters used to distinguish the species include habit type, thallus size, presence or absence of cortication on the main axis, the branching pattern and origin, shape of terminal cortical cells, length/diameter ratio of axial cells and position of the tetrasporangia (Saenger & Wollaston 1982, Norris et al. 1984, Wollaston & Womersley 1998, Mateo-Cid et al. 2002, Schneider 2004).
With the re-organization of the family Ceramiaceae, as proposed by Choi et al. (2008) on the basis of gene-sequence data, Crouania is now assigned to the family Callithamniaceae. The genus is widely distributed in warm temperate and tropical seas (Norris et al. 1984), with 16 species currently recognized (Guiry & Guiry 2012), four of which are reported for the Western Atlantic (Wynne 2011): C. attenuata (C. Agardh) J. Agardh, C. elisiae C.W. Schneid., C. mayae Mateo-Cid, Mendoza-González & Searles and C. pleonospora W.R. Taylor. The status of a fifth species, C. capricornica Saenger & E.M. Woll., in the flora of the Western Atlantic, is uncertain. That species, which has a type locality of Port Curtis, Queensland, Australia (Saenger & Wollaston 1982), was reported to occur in Martinique, Lesser Antilles, by Bucher & Norris (1995), which was its first report from the Atlantic Ocean. Schneider (2004), however, pointed out that Bucher & Norris (1995) depicted elongated outer cortical cells and more than one sporangium per whorl-branchlet in their material, features that are in conflict with Saenger & Wollaston’s (1982) original description of this species. This led Wynne (2005) to dismiss the record from his checklist. But a second record of this species was missed, namely, the report of C. capricornica by Díaz-Pulido & Díaz-Ruiz (2003) from Caribbean Colombia. In addition to their record of this Australian-based species for the Atlantic coast of Colombia, Díaz-Pulido & Díaz-Ruiz (2003) reported the occurrence of C. attenuata and C. pleonospora.
In the present paper we describe the new species Crouania pumila, collected subtidally from Old Providence Island, Seaflower Biosphere Reserve, Caribbean Colombia.
Materials and Methods
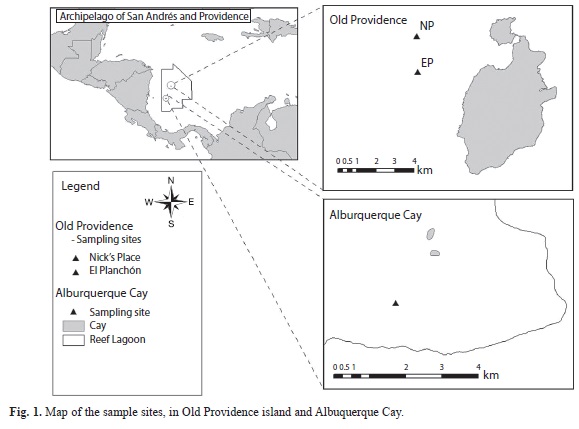
The Archipelago of San Andres, Old Providence and Santa Catalina islands are located in the western portion of the Caribbean Sea, between 10-18° N -78-82° W, with a marine extension of more than 250 000km2, of which 5 000km2 are shallow water. Three main islands (San Andres, Old Providence and Santa Catalina) and several cays, shoals and banks forming the archipelago, were declared as the Seaflower International Biosphere Reserve in 2000. Old Providence is a small (17km2) volcanic island, 228km off the coast of Nicaragua (Fig. 1). The island harbors the second largest barrier reef of the Caribbean Sea (32km long), situated on the east side of the island. Among the southern cays of the Archipelago, Albuquerque lies about 137km southwest of Old Providence, 190km2 off the Nicaraguan coast. The key is made up of two islands that were formed by the accumulation of sediments on the reef lagoon (Fig. 1).
During our field surveys, specimens were collected by SCUBA at a depth of 16-37m, in October 2009, in Old Providence, at two diving sites. The first one is El Planchón, a ship wreck lying at about 20m depth, covered by a rich coral community and associated macroalgae. The second site is Nick´s Place, a leeward cliff on the platform of the island, with a terrace at about 16m and a slope descending abruptly, where the specimen was collected at 37m.; the specimens were epiphytic on larger red algae (Chondria sp.) or as part of turf. Additional specimens were collected in Albuquerque Cay, by SCUBA, at a depth of 1-2m, in October 2010, also epiphytic on larger algae. The alga was fixed in 5% formalin in seawater and mounted on microscope slides, unstained, in 50% Karo corn syrup in a distilled water solution to which few phenol crystals were added to prevent fungal growth. The slides for herbarium specimens were sealed with nail polish. Cross-sections were made by hand with a razorblade. The specimens were observed and photographed with an Olympus BX52 light microscope, connected to a digital camera Moticam 2 300.
Results
Crouania pumila
B. Gavio, V. Reyes-Gómez et M.J. Wynne sp. nov.
Plantae epiphyticae, pusillae, usque 3.5mm altae, roseolae, leviter calcareae. Axis ramique cylindrici, articulati, gelatinosi, ecorticati. Plantae sistema ramisympodiale formantes. Cellulae axiales 30-50µm diametro et 70-100µm altae, gradatim decrescentes usque cellula apicalis tholiformis, 7.0-7.5µm diámetro et 7.0-10µm alta. Tres rami breves verticillati atque determinati in omni nodo, ramulis verticillorum di-trichotome ramosis. Prima cellula ramorum magis parva, isodiametra, 15-25µm diametro et 15-25µm alta. Secunda cellula ramorum diametro similare sed magis alta, 20-50µm longa. Cellulae terminales ramorum elongatae, 10-17.5µm altae, 5µm latae.
Rhizoidea multicellulares cum cellulis 10-23µm diametro et 75-120mm longis. Gonimoblasta cum carposporas isodiametras, 25-30µm diametro. Tetrasporangia ovales 37-50µm diámetro et 50-60µm longa, uno vel duo in quo verticillo. Plantae masculae ignotae.
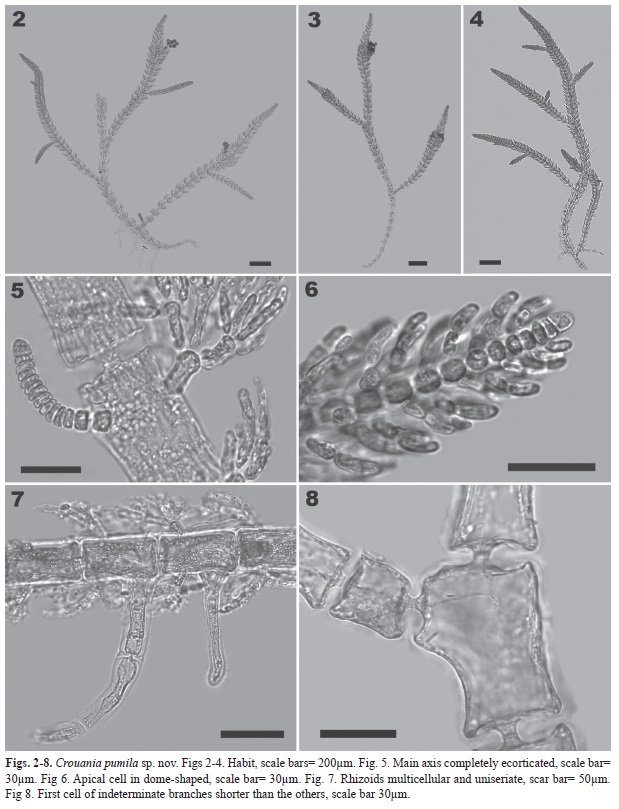
Plants epiphytic, small, to 3.5mm long, light pink, lightly calcified. Thallus decumbent with prostrate main axes attached to other algae, ramisympodially branched (Figs. 2-4). Main axis completely ecorticate (Figs. 5-8), with primary axial cells 30-50µm in width and 70-100µm in length (length/width ratio: 1.5-2.5), gradually tapering distally to a dome-shaped apical cell 7-7.5µm wide and 7-10µm long (Fig. 6). Rhizoids multicellular, uniseriate, composed of cells 10-23µm wide and 75-120µm long, arising proximally from axial cells (Fig. 7). First cell of indeterminate branches shorter than the others, almost isodiametric, 15-25µm wide (Figs. 5, 8-9). Second cell of indeterminate branches longer, up to 20-25µm long (Fig. 9). Indeterminate branches arising distally from axial cells (Figs. 5, 8-9). Determinate branchlets in whorls of three (Fig. 10), tri-tetrachotomously divided, with first cell of the branchlet elongate, 15-17.5µm long, 7.5-10µm wide, terminal cells elongate, 10-17.5µm long, 5µm wide. Carposporophyte a compact rounded mass of carpospores (Figs. 11-12); individual carpospores 25-30µm in diameter (Fig. 13). Tetrasporangia ovoid, 37-50µm in diameter, 50-60µm long, one or two per whorl (Figs 14-17, Table 1). Spermatangial thalli not seen.
HOLOTYPE: VR 191, collected by B. Gavio and J.F. Ortiz, 18 October, 2009, at a depth of 16m COL 550268 (Herbario Nacional de Colombia, Bogotá Colombia), a female plant.
ISOTYPES: COL 550269, female; COL 550270 vegetative plant.
TYPE LOCALITY: The type specimens were epiphytic on Chondria sp., at El Planchón, Old Providence Island, Colombia. 13° 22’17” N, 81° 25’05” W
ADDITIONAL COLLECTIONS EXAMINED [PARATYPES]:
LRP 78, collected by M. Albis in Albuquerque Cay, 22 September, 2010, 1-2m depth. COL 553429, tetrasporophyte, epiphyte on Chondria sp.
VR 185, collected by V. Reyes and H. Velasquez, 18 October, 2009, at Nick´s Place, Old Providence Island, Colombia, 13° 23’ 18.5” N-81° 25’ 07.3” W, at a depth of 37m, turf with Centroceras sp.
DISTRIBUTION: Old Providence Island and Albuquerque Cay, Caribbean Colombia.
Discussion
In all of its characters, C. pumila conforms well to the genus. Its most distinctive character is the diminutive size of the thalli, which, in all the specimens examined, do not exceed 0.5cm in length. Considering that we found both cystocarpic and tetrasporic thali, we can state that the alga reaches full maturity at this size, and probably does not exceed the size that we observed. Of the 16 currently accepted (Guiry & Guiry 2012) only three other species of Crouania have a comparable size: C. destriana Wollaston from D’Estree Bay, Kangaroo Island, South Australia (Wollaston 1968), C. ischiana (Funk) Boudouresque & M. Perret-Boudouresque from Isla Ischia, Italy, Mediterranean Sea (Funk 1955), and C. minutissima Yamada from the Atoll of Ant, Caroline Islands, South Pacific (Yamada 1944). However, it is easily distinguishable from C. destriana by the complete absence of cortication and the elongate terminal cells of the determinate branches. Such terminal cells in C. destriana are ovoid (Wollaston & Womersley 1998). C. minutissima has relatively large axial cells, 200-250µm long and 64µm wide, with a L/W ratio of 3-4: 1 (Yamada 1944), whereas in our species the axial cells are 70-100µm long and 30-50µm wide (L/W ratio: 1.5-2.5:1). C. ischiana has a known distribution limited to the Mediterranean (Gómez-Garreta et al. 2001), ranging from the Andalucian coast of Spain in the West (Conde et al. 1996) to the Gulf of Saronikos of Greece in the East (Diapoulis & Verlaque 1981). Its size is more similar to our specimens; however, it has a monopodial branching pattern in contrast to the ramisympodial branching present in the new species from Colombia (Funk 1955). Furthermore, in our species the main axes are widest in their middle part and taper towards the apex, while in C. ischiana the diameter of the axes is more even (Coppejans 1983). The presence of hyaline hairs at the apex, reported in C. ischiana (Coppejans 1983), has not been observed in the new species.
In the Western Atlantic, the four other species of Crouania reported to date can be distinguished from our specimens because these other species are much larger in size.
It seems prudent to include C. capricornica in the discussion on the basis that there have been reports of that species from the Western Atlantic. Similar to the new species, C. capricornica has a prostrate habit, and the axial cells in that species have a size range closer to C. pumila. However, C. capricornica has sub-spherical terminal cells of the determinate branchlets, whereas in C. pumila they are very elongate. The Atlantic occurrence of C. capricornica remains uncertain. Material of C. capricornica from Martinique, described by Bucher & Norris (1995), should be examined carefully to determine its taxonomic status. Schneider (2004) questioned the identity of the Martinique material based on the presence of more than one sporangium per whorl-branch and the elongated outer cortical cells, which, according to him, are not in agreement with the original description of C. capricornica. Saenger & Wollaston (1982), however, found slightly elongate terminal cells in the type specimen of the taxon and stated that “in the limited material available, no more than one tetrasporangium has been observed on each fertile whorl-branchlet” (page 81) suggesting that they had limited access to tetrasporic material of the species, and they could not dismiss a priori the presence of several tetrasporangia per whorl. We consider that the shape of the terminal cells of the determinate branches is not a good taxonomic character because in the same plant, spherical, slightly elongate to elongate cells may be observed (pers. obs.). Thus, species differentiation based solely or mainly on this morphological character should be reconsidered.
We are able to dismiss the record of C. capricornica for the Atlantic coast of Colombia, reported by Díaz-Pulido & Díaz-Ruiz (2003). We analyzed the specimen (INV-RDH 398) collected by G. Díaz-Pulido, which is deposited in the Museum of Marine Natural History of Colombia MHNMC, Santa Marta, and could determine that the material does not correspond to the genus Crouania because the axial cells bear a whorl of four determinate branchlets, not three as in the genus Crouania. The identity of the specimen could not yet be resolved, but at this time we can exclude C. capricornica from the Colombian Caribbean marine flora.
There are many examples where size has been used as one of the characters to distinguish species of red algae from their congeners, for example: Jania pumila J.V. Lamouroux, Amphiroa minutissima W.R. Taylor, Predaea pusilla (Berthold) Feldmann, Grateloupia minima P.L. Crouan & H.M. Crouan, Symphiocladia pumila (Yendo) Uwai & Masuda, and Scagelothamnion pusillum (Ruprecht) Athanasiadis. These species are currently recognized (Guiry & Guiry 2012).
Acknowledgments
The authors wish to thank Juan Felipe Ortiz, Hernán Velasquez and Felipe Cabeza´s diving center for logistic support in collecting field trip. Douglas Rivera collaborated making the map. Lanie Ruiz made the pictures of the tetrasporic specimen. Craig Schneider provided useful literature and information. We acknowledge the Museum of Marine Natural History of Colombia (MHNMC), INVEMAR, for the loan of the specimen INV-RDH 398 identified as “Crouania capricornica”. The present research was funded by the Universidad Nacional de Colombia, Caribbean branch, through the projects # 20501003060, 20101003400 and 20101003960.
References
Bucher, K.E. & J.N. Norris. 1995. Marine algae new to the Lesser Antilles, including Mazoyerella kraftii sp. nov. (Ceramiales, Rhodophyta). Carib. J. Sci. 31: 1-24. [ Links ]
Choi, H.G., G.T. Kraft, H.S. Kim, M.D. Guiry & G.W. Saunders. 2008. Phylogenetic relationships among lineages of the Ceramiaceae (Ceramiales, Rhodophyta) based on nuclear small subunit rDNA sequence data. J. Phycol. 44: 1033-1048. [ Links ]
Conde, F., A. Flores-Moya, J. Soto, M. Altimirano & A. Sánchez. 1996. Check-list of Andalusia (S. Spain) seaweeds. III Rhodophyceae. Acta Bot. Malacitana 21: 7-33. [ Links ]
Coppejans, E. 1983. Iconographie d’algues Méditerranéennes: Chlorophyta, Phaeophyta, Rhodophyta, J. Cramer, Vaduz, Germany. [ Links ]
Diapoulis, A. & M. Verlaque. 1981. Contribution a la flore des algues marines de la Grèce. Thalassographica 1: 99-102. [ Links ]
Díaz-Pulido, G. & M. Díaz-Ruiz. 2003. Diversity of benthic marine algae of the Colombian Atlantic. Biota Colombiana 4: 203-246. [ Links ]
Funk, G. 1955. Beiträge zur Kenntnis der Meeresalgen von Neapel zugleich mikrophotographischer Atlas. Pubblicazioni della Stazione Zoologica di Napoli 25:1-178. [ Links ]
Gómez-Garreta, A., T. Gallardo, M.A. Ribera, M. Cormaci, G. Furnari, G. Giaccone & C.F. Boudouresque. 2001. Check-list of the Mediterranean seaweeds. III. Rhodophyceae Rabenh. 1. Ceramiales Oltm. Bot. Mar. 44: 425-460. [ Links ]
Guiry, M.D. & W.M. Guiry. 2012. AlgaeBase, National University of Ireland, Galway. (http://www.algaebase.org). [ Links ]
Maggs, C.A. & M.H. Hommersand. 1993. Seaweeds of the British Isles. Volume 1 Rhodophyta. Part 3A Ceramiales, The Natural History Museum, London, England. [ Links ]
Mateo-Cid, L.E., A.C. Mendoza-Gonzalez & R.B. Searles. 2002. New Mexican records of marine algae including Crouania mayae sp. nov. (Ceramiaceae, Rhodophyta). Carib. J. Sci. 38: 205-221. [ Links ]
Norris, R.E., E.M. Wollaston & M.J. Parsons. 1984. New terminology for sympodial growth in the Ceramiales (Rhodophyta). Phycologia 23: 233-237. [ Links ]
Norton, T.A., M. Melkonian & R.A. Andersen. 1996. Algal biodiversity. Phycologia 35: 308-326. [ Links ]
Saenger, P. & E.M. Wollaston. 1982. A new species of Crouania (Rhodophyta: Ceramiaceae) from Port Curtis, Queensland. Proc. R. Soc. Qd. 93: 79-82. [ Links ]
Schneider, C.W. 2004. Notes on the marine algae of the Bermudas. 6. Some rare or newly informed Ceramiales (Rhodophyta), including Crouania elisiae sp. nov. Phycologia 43: 563-578. [ Links ]
Taylor, W.R. 1960. Marine algae of the eastern tropical and subtropical coasts of Americas, University of Michigan, Michigan, USA. [ Links ]
Wollaston, E.M. 1968. Morphology and taxonomy of Southern Australian genera of Crouanieae Schmitz (Ceramiaceae, Rhodophyta). Austr. J. Bot. 16: 217-417. [ Links ]
Wollaston, E.M. & H.B.S. Womersley. 1998. Tribe Crouanieae, p. 43-67. In H.B.S. Womersley (ed.). The marine benthic flora of southern Australia. Part IIIC. State Herbarium of South Australia, South Australia, Australia.
Wynne, M.J. 2005. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: second revision, Nova Hedwigia, Beihefte 129, Schweizerbart Science Publishers, Stuttgart, Germany. [ Links ]
Wynne, M.J. 2011. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: third revision, Nova Hedwigia, Beihefte 140, Schweizerbart Science Publishers, Stuttgart, Germany. [ Links ]
Yamada, Y. 1944. A list of the marine algae from the Atoll of Ant. Sci. Pap. Inst. Algol. Res., Fac. Sci., Hokkaido Imp. Univ. 3: 31-45. [ Links ]
Choi, H.G., G.T. Kraft, H.S. Kim, M.D. Guiry & G.W. Saunders. 2008. Phylogenetic relationships among lineages of the Ceramiaceae (Ceramiales, Rhodophyta) based on nuclear small subunit rDNA sequence data. J. Phycol. 44: 1033-1048. [ Links ]
Conde, F., A. Flores-Moya, J. Soto, M. Altimirano & A. Sánchez. 1996. Check-list of Andalusia (S. Spain) seaweeds. III Rhodophyceae. Acta Bot. Malacitana 21: 7-33. [ Links ]
Coppejans, E. 1983. Iconographie d’algues Méditerranéennes: Chlorophyta, Phaeophyta, Rhodophyta, J. Cramer, Vaduz, Germany. [ Links ]
Diapoulis, A. & M. Verlaque. 1981. Contribution a la flore des algues marines de la Grèce. Thalassographica 1: 99-102. [ Links ]
Díaz-Pulido, G. & M. Díaz-Ruiz. 2003. Diversity of benthic marine algae of the Colombian Atlantic. Biota Colombiana 4: 203-246. [ Links ]
Funk, G. 1955. Beiträge zur Kenntnis der Meeresalgen von Neapel zugleich mikrophotographischer Atlas. Pubblicazioni della Stazione Zoologica di Napoli 25:1-178. [ Links ]
Gómez-Garreta, A., T. Gallardo, M.A. Ribera, M. Cormaci, G. Furnari, G. Giaccone & C.F. Boudouresque. 2001. Check-list of the Mediterranean seaweeds. III. Rhodophyceae Rabenh. 1. Ceramiales Oltm. Bot. Mar. 44: 425-460. [ Links ]
Guiry, M.D. & W.M. Guiry. 2012. AlgaeBase, National University of Ireland, Galway. (http://www.algaebase.org). [ Links ]
Maggs, C.A. & M.H. Hommersand. 1993. Seaweeds of the British Isles. Volume 1 Rhodophyta. Part 3A Ceramiales, The Natural History Museum, London, England. [ Links ]
Mateo-Cid, L.E., A.C. Mendoza-Gonzalez & R.B. Searles. 2002. New Mexican records of marine algae including Crouania mayae sp. nov. (Ceramiaceae, Rhodophyta). Carib. J. Sci. 38: 205-221. [ Links ]
Norris, R.E., E.M. Wollaston & M.J. Parsons. 1984. New terminology for sympodial growth in the Ceramiales (Rhodophyta). Phycologia 23: 233-237. [ Links ]
Norton, T.A., M. Melkonian & R.A. Andersen. 1996. Algal biodiversity. Phycologia 35: 308-326. [ Links ]
Saenger, P. & E.M. Wollaston. 1982. A new species of Crouania (Rhodophyta: Ceramiaceae) from Port Curtis, Queensland. Proc. R. Soc. Qd. 93: 79-82. [ Links ]
Schneider, C.W. 2004. Notes on the marine algae of the Bermudas. 6. Some rare or newly informed Ceramiales (Rhodophyta), including Crouania elisiae sp. nov. Phycologia 43: 563-578. [ Links ]
Taylor, W.R. 1960. Marine algae of the eastern tropical and subtropical coasts of Americas, University of Michigan, Michigan, USA. [ Links ]
Wollaston, E.M. 1968. Morphology and taxonomy of Southern Australian genera of Crouanieae Schmitz (Ceramiaceae, Rhodophyta). Austr. J. Bot. 16: 217-417. [ Links ]
Wollaston, E.M. & H.B.S. Womersley. 1998. Tribe Crouanieae, p. 43-67. In H.B.S. Womersley (ed.). The marine benthic flora of southern Australia. Part IIIC. State Herbarium of South Australia, South Australia, Australia.
Wynne, M.J. 2005. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: second revision, Nova Hedwigia, Beihefte 129, Schweizerbart Science Publishers, Stuttgart, Germany. [ Links ]
Wynne, M.J. 2011. A checklist of benthic marine algae of the tropical and subtropical western Atlantic: third revision, Nova Hedwigia, Beihefte 140, Schweizerbart Science Publishers, Stuttgart, Germany. [ Links ]
Yamada, Y. 1944. A list of the marine algae from the Atoll of Ant. Sci. Pap. Inst. Algol. Res., Fac. Sci., Hokkaido Imp. Univ. 3: 31-45. [ Links ]
*Correspondencia a:
Brigitte Gavio. Universidad Nacional de Colombia, Departamento de Biología, Ciudad Universidad, Bogotá, Colombia; bgavio@unal.edu.co. Universidad Nacional de Colombia, sede Caribe, San Luis Free Town # 52-44, San Andrés Isla, Colombia; bgavio@unal.edu.co
Viviana P. Reyes-Gómez. Universidad Nacional de Colombia, Departamento de Biología, Ciudad Universidad, Bogotá, Colombia; vpreyesg@unal.edu.co.
Michael J. Wynne. University of Michigan Herbarium, Ann Arbor, MI 48108, USA; mwynne@umich.edu
1. Universidad Nacional de Colombia, Departamento de Biología, Ciudad Universidad, Bogotá, Colombia; bgavio@unal.edu.co, vpreyesg@unal.edu.co
2. Universidad Nacional de Colombia, sede Caribe, San Luis Free Town # 52-44, San Andrés Isla, Colombia; bgavio@unal.edu.co
3. University of Michigan Herbarium, Ann Arbor, MI 48108, USA; mwynne@umich.edu
Received 09-VIII-2012. Corrected 12-XII-2012. Accepted 22-I-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}