










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkRevista de Biología Tropical
On-line version ISSN 0034-7744Print version ISSN 0034-7744
Rev. biol. trop vol.54 suppl.3 San José Dec. 2006
Temporal and spatial dynamics of Lutjanus cyanopterus (Pisces: Lutjanidae) and L. jocu spawning aggregations in the United States Virgin Islands
Elizabeth Kadison, Richard S. Nemeth, Steve Herzlieb & Jeremiah Blondeau
Center for Marine and Environmental Studies, University of the Virgin Islands, 2 John Brewers Bay, St. Thomas, US Virgin Islands, 00802-9990; ekadiso@uvi.edu
Received 01-VI-2006. Corrected 02-X-2006. Accepted 13-X-2006.
Abstract: The Grammanik bank is a deep reef (35-40 m) located approximately 14 km south of St. Thomas USVI, on the southern edge of the Puerto Rico shelf. The relatively narrow 1.5 km long reef annually hosts spawning aggregations of several species of groupers, snappers and jacks. Until February of 2005, when temporary seasonal protection was offered by the Caribbean Fisheries Management Council, the bank was subject to moderate or heavy exploitation from fishers. Cubera snapper (Lutjanus cyanopterus) were observed aggregating on the Grammanik bank in relatively large numbers from 2003 to 2005 and dog snapper (L. jocu) aggregations were observed in 2005. The arrival and departure of these aggregations appeared to be seasonal and to revolve around the moon phase. Cubera snapper spawning seasonality coincided with that elsewhere in the Caribbean and off south Florida. Schools of up to 1000 fish of each species were observed as well as pre-spawning behavior and milt release. Aggregations of both species showed high site fidelity within seasons but cubera snapper aggregations sites moved between seasons. The authors recommend year-round closure of the Grammanik bank based on its importance as critical fish habitat and a multi-species aggregation area. Rev. Biol. Trop. 54 (Suppl. 3): 69-78. Epub 2007 Jan. 15.
Key words: spawning aggregations, cubera snapper, dog snapper, Lutjanidae, MPA.
Many large reef fish including groupers (Serranidae) and snappers (Lutjanidae) aggregate in large numbers to spawn at specific times and locations each year. These aggregations are generally seasonal and revolve around the lunar cycle (Domeier and Colin 1997). The fidelity of many fish to specific spawning aggregation sites for decades or more has been documented both anecdotally and scientifically (Johannes 1978, Sadovy 1994, Colin 1996, Luckhurst 1998). A consequence of this phenomenon has been the over-exploitation of such sites worldwide by fishers, leading to significant declines or even the collapse of spawning populations in many areas (Olsen and LaPlace 1978, Sadovy 1997, Johannes et al. 1999).The importance of large spawning aggregations for the successful reproduction of participating species remains poorly understood, and even less clear is the part these aggregations play in the ultimate sustainability of regional fish stocks. However, the need to manage spawning aggregation sites and to incorporate them into marine protected areas (MPAs) is clear (Bohnsack 1998, Lindeman et al. 2000, Luckhurst 2003), as is the need for additional field assessments to collect biological and behavioral information to determine the current status of spawning populations.
In the Caribbean, aggregations of commercially important serranids including Nassau grouper, Epinephelus striatus (Bloch, 1792) and red hind, E. guttatus (Linnaeus, 1758) have been relatively well documented and studied (Colin et al. 1987, Colin 1992, Tucker et al. 1993, Shapiro et al. 1993a, b, Carter et al. 1994, Sadovy and Colin 1995, Aguilar-Perera and Aguilar-Davila 1996, Beets and Friedlander 1992), Whaylen et al. 2004, Nemeth 2005, Whiteman et al. 2005). In comparison, very little has been reported on lutjanid spawning aggregations in the region. Dog snapper, Lutjanus jocu (Bloch and Schneider, 1801) and cubera snapper, L. cyanopterus (Cuvier, 1828) are referred to as transient spawners, meaning fish travel long distances to aggregation areas and aggregations generally occur only one or two months a year but persist for several days during that period (Domeier and Colon 1997). Seasonality for both dog and cubera snapper is known primarily from traditional knowledge and from the collection of reproductive stage fish during specific periods of the year. Direct observational data on dog snapper spawning aggregations is extremely limited and comes only from Little Cayman (Whaylen et al. 2004) and the Mesoamerican reef off of Belize (Carter and Perrine 1994, Domeier and Colon 1997, Heyman et al. 2001, Heyman and Requena 2002). With the exception of the comprehensive account on Gladden Spit by Heyman et al. (2005) data is absent for cubera snapper spawning. Spawning aggregations of both species are known to occur off Cuba and the Florida Keys but evidence comes only from fishers and catch reports (Lindeman et al. 2000, Claro and Lindeman 2003).
The Grammanik bank is a deep water reef located 14 km south of St. Thomas on the southern edge of the insular platform known as the Puerto Rican shelf. The bank was discovered as a grouper, snapper and parrotfish aggregation area by fishermen in the mid-1950s but was fished very lightly until 1990 (Louis Blanchard, pers. comm.). In the 1990s, an area 8 km west of the Grammanik bank was closed seasonally and then year-round to fishing by the Caribbean Fisheries Management Council (Federal Registers 55(213) November 2, 1990 and 64(213) November 4, 1999 respectively). This area, now known as the Marine Conservation District (MCD) included both a red hind grouper and a traditional Nassau grouper aggregation site. With the closure of the MCD for fishing purposes, pressure shifted to the Grammanik bank. Grouper landings from the bank were reported in excess of 20 000 lbs annually from 1999-2001 (Turbe, pers. comm.). Grouper were targeted from February through April each year but in May grouper catches declined and the fish were believed to leave the bank, replaced by large cubera snapper (Bobby Vante, pers. comm.). Although exploited as a food fish in Puerto Rico (Olsen, pers. comm.), Belize (Anon 2000), Honduras (Gobert et al. 2005), Jamaica (Thompson and Munro 1983), Cuba (Claro and Lindeman 2003) and the continental United States (Lindeman et al. 2000), cubera snapper and dog snapper are believed to be extremely ciguatoxic in the Virgin Islands and little or no fishery exists for them in most of the lesser Antilles. The cubera snapper is listed as a vulnerable species by the International Union for the Conservation of Natural Resources (IUCN), but the condition of regional stocks for cubera as well as the dog snapper is unknown, due primarily to lack of fisheries data.
Cubera snappers, generally solitary reef dwellers (Allen 1985) and usually absent from the Grammanik bank, were first observed in large aggregations in 2003. Dog snapper, also normally rare on the bank, were observed aggregating in 2005. Because relatively little is known about spawning behavior of cubera snapper and dog snapper in the eastern Caribbean, this paper presents a preliminary description of the temporal and spatial dynamics of aggregations of these species on the Grammanik bank.
Materials and methods
Site identification and description: The historic grouper and snapper spawning area known as the Grammanik bank was located through informal interviews with local St. Thomas fishers in 2002. The bank sits on the southern edge of the insular Puerto Rican shelf approximately 14 km south of St. Thomas, USVI at 18º11 N latitude, and 64º57 W longitude. It runs in a roughly east/west direction in 35-40 m of water and is approximately 1.5 km long and 100 m wide at the widest point. The benthic substrate is dominated by plated Montastrea corals at relatively high densities (Herzlieb et al., in press). The western end of the reef is hard-bottom colonized by sponges and gorgonians. On the south edge the bathymetry drops rapidly to 200 m and on the north side, separated by a 20-30 m wide sand channel, another Montastrea dominated reef runs parallel to the main bank (Fig. 1).

Field observations: The Grammanik bank was surveyed by divers swimming alone or with Apollo av-1 underwater scooters from March through August of 2003 and 2004, and February through June 2005. Field work was conducted opportunistically around the full moon over these three years, but was most intensive during the months of March and April. Dives were made throughout the day but were primarily in the late afternoon, from 15:00 to 18:00 hr. This pattern was established based on observations early in the study of increased aggregating and pre-spawning behavior by several species of fish late in the afternoon (Nemeth et al. 2004). The eastern and western ends of the bank were marked with a Garmin GPS72 global positioning satellite (GPS) receiver and the reef was divided into six sections, all approximately 160 m in longitudinal distance. These segments were marked on the GPS and were delineated underwater by numbered 20 cm polystyrene trap floats attached to the bottom on a 3 m long polypropylene line. Visual fish surveys were made along the bank by divers using Nitrox. In 2004 and 2005 additional opportunistic surveys were made on the northern reef running parallel to the main bank. Generally two divers swam or used scooters to move along the bank, 25 m apart and 5 m off of the bottom, recording all grouper and snapper by species and number within a 20 m wide belt transect beneath them. Transects were swam along the reef in an east or west direction depending on the current. Horizontal visibility, based on dive logs, ranged from 20-335 m. The total effective search area per survey was approximately 60 000 m2 or 40% of the bank. Data was recorded on an underwater slate separately for each buoyed section. If a large school of snapper was encountered, the number of individuals in the entire school was estimated, as well as the maximum, minimum and median size (TL) of the fish. Any unusual behavior or color patterns were recorded. The location of the large snapper schools were recorded on the GPS and subsequent dives were made on those locations to collect additional aggregation population size and behavioral data. The transient and ephemeral nature of the snapper aggregations precluded using transects or point counts to estimate fish density on these dives. Instead, observations and total population estimates were made haphazardly focusing on the area of highest fish density and the most dramatic fish behavior. Additional size estimates of cubera snapper were made by divers using a 1 m wide t-shaped bar constructed of 1.27 cm PVC pipe marked with 5 cm increments. Total length estimates were divided into 10 cm size classes. Digital video was taken with a Sony TRV900 in a Light in Motion underwater housing and digital still photos were taken with a Canon PowerShot S45. During surveys in June 2004, two Sony 532 nm wavelength green laser pointers were mounted to an underwater video housing and experimentally used to measure a series of fish. This method of estimating size was unsuccessful and was discontinued because large fish showed an extreme aversion to the laser beams.
Measurements of temperature were made once every hour from 3 April 2003 to 16 April 2005 by an Aanderaa RCM 9 MkII data recorder. The recorder was mounted approximately 8 km west of the Grammanik bank close to the shelf edge in 38 m water depth. Data for the period from 03 November 2004 through 24 January 2005 was lost due to flooding of the data recorder housing. Data from this period was supplemented with digital temps from dive computers. Monthly mean temperatures were calculated for all other months and compared to the annual arrival and departure of cubera snapper aggregations to and from the bank.
Results
Lutjanus cyanopterus: The initial sighting of an unusually large group of cubera snapper was 15 May 2003, the day of the full moon. An aggregation of 350- 400 fish was observed on the eastern end of the bank at 18º11.40 N latitude, 64º57.02 W longitude. A month prior to this, on 17 April 2003, over 50 cubera snapper had been seen dispersed fairly evenly across the reef from this point to the western edge of the bank (18º11.26 N, 64º57.60 W). The size of the fish in the aggregation ranged from 40 cm to 130 cm TL with a median size of 80 cm TL (Fig. 2). The fish circled the divers several times, moving in a clockwise direction. They then ascended in the water column approximately 20 m as a school, with three or four very large individuals in the lead. After approximately 2 min the school streamed back down to the reef. At the reef groups of 3-6 fish oriented themselves head down and tail up together, their heads sticking down into the coral and bodies twitching or shaking vigorously. This activity lasted from 30 to 60 s and was observed three times, each by a separate group of fish, during one 17 min dive. This behavior was followed by more circling and another slow ascent as a group toward the surface. Activities were repeated until the divers left the water. The following two days a similar number of fish was found at the same location, repeating the behavior observed the previous day. The school surrounded the divers and circled them closely, ascending up into the water column and streaming back down to the reef, but always staying within sight of the divers. The fish appeared silver-gray, as they do outside the aggregation season, and no distinctive markings or aggressive behavior was observed. Several fish had distended abdomens but these fish were similar in size and color to the other fish. More tightly packed groups of 4-7 individuals within the larger aggregation were noted. Fish in these small groups were occasionally seen twitching and touching, at times head down and tail up. Milt release was observed four times on 16 May 2003 as fish ascended toward the surface. In June and July, on four dives made during the week following the full moon, divers again found cubera snapper on the aggregation site. The fish were in slightly smaller numbers but except for milt release, the same type of activity was observed.
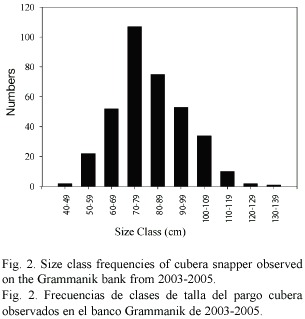
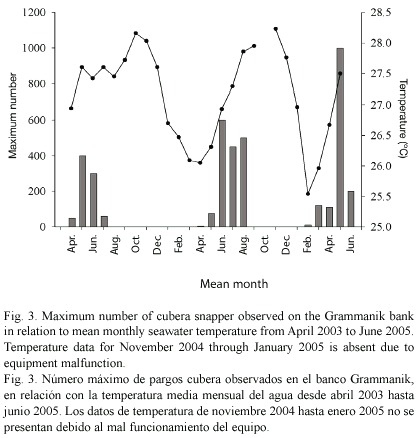
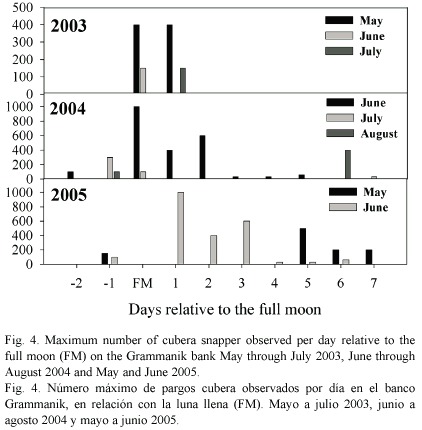
In May of 2004 and March and April of 2005, up to 70 cubera snapper were again seen dispersed across the reef in groups of 1-6 fish. Relatively large aggregations of up to 1000 fish were observed between June and August of 2004 and in May and June of 2005 (Fig. 3). Seawater temperature (at 35 m) during these two years showed seasonal fluctuations ranging from 25.6ºC in February 2005 to 28.2ºC in October 2004. The appearance of cubera snapper on the bank coincided with rising sea temperatures (Fig. 3). Aggregation formation was observed during months with mean temperatures above 26.9ºC. In addition to seawater temperature, cubera aggregations appeared to form around the lunar cycle. We observed spawning aggregations on the Grammanik bank within 10 days following the full moon each month (Fig. 4) but observed less than 10 individuals per day on nine separate dives conducted within four days after the new moon in May and June of 2005.
The cubera aggregation was consistent in location from month to month each year, however the primary aggregation shifted along the reef from one year to the next. In 2003 the aggregation was on the east end of the bank but in 2004 and 2005 the site shifted approximately 350 m and 600 m respectively to the west (Fig. 1). During all three years the aggregation was found primarily on the top of the reef and over the southern edge. In 2004 and 2005 the schools were more transient and the fish seemed much less attracted to divers. There was usually some initial circling followed by ascending and descending as a school, but this was generally followed by the aggregation splitting into smaller groups which dispersed out onto the bank. Although more dives were made in 2004 and 2005 and the aggregation size on several days was greater than 400 fish, less pre-spawning behavior in the form of fish interaction or movement (twisting, shaking or vertical orientation) was observed and no milt release was seen.
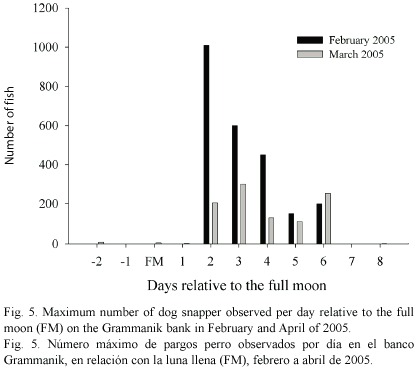
Lutjanus jocu : An aggregation of dog snapper was first observed on 26 February 2005, two days after the full moon. The fish were aggregated over the sand channel between the two Montastrea banks at 18º11.34 N, 64º57.37 W (Fig. 1). Approximately 1000 fish were seen in the aggregation. Size ranged from 45 cm to 75 cm TL with a median size of 55 cm TL. No dramatic or unusual color changes were noted except striping of some fish. The fish swam up into the water column as a school and then streamed back down to the reef. Otherwise they exhibited no dramatic or unusual behavior. Occasionally the large school would split into smaller ones, but these would merge after several minutes. Dog snapper aggregations were observed over the next four days at the same location and to the northwest over the northern reef (Fig. 1). Although most of the snapper stayed in con-specific groups, some mixed into the yellowfin grouper aggregation that was also present on the northern bank. No dog snapper were observed 12 days past the February full moon and an aggregation was not seen again until 28 March 2005, two days after the following full moon (Fig. 5). The dog snapper aggregation was again observed over the sand channel directly adjacent to the Montastrea reef. Behavior was similar to that observed in February. On 30 March 2005 we observed some milt release. The aggregation was seen for five consecutive days, but disappeared by 2 April 2005 (Fig. 5).
Discussion
Cubera snapper: Large aggregations of cubera snapper were observed on the Grammanik bank each year from 2003 to 2005 although no actual spawning was observed. The majority of dives were made in the late afternoon or early evening when fish were most abundant and the time that cubera snapper spawn on Gladden Spit off Belize (Heyman et al. 2001, 2005). Based on number, behavior, and timing of fish as well as the distended abdomens and milt release observed, we are confident that spawning was imminent. Many of the behaviors described by Heyman et al. (2005) were recorded on the Grammanik bank including twitching, nuzzling and the vertical orientation of fish in small groups close to the reef with their heads in the substrate. Unlike the description by Heyman, no obvious color morphs were distinguished and we did not observe fish spiraling tightly up into the water column. The color changes and the spiral formation may occur only immediately before fish spawn.
The observation of actual snapper spawning has proven elusive to many scientists (Carter and Perrine 1994, Domeier and Colon 1997, Claro and Lindeman 2003, Burton et al. 2005) and may occur at night on the Grammanik bank as it is believed to in other areas (Domeier and Colin 1997, Claro and Lindeman 2003). While diving the bank during the grouper and snapper spawning seasons we regularly observed many sharks including the great hammerhead, Sphyrna mokarran (Ruppell, 1835), bull shark, Carcharhinus leucas (Valenciennes, 1841), Caribbean reef shark, C. perezii (Poey, 1876), and lemon shark, Negaprion brevirostris (Poey, 1868). Although we did not observe any attacks on aggregating fish, large individuals were encountered and at times shark densities were relatively high. Sharks were not observed on the Grammanik bank during any dives from 2003-2005 that occurred outside of the spawning season. Larval predation occurs at Gladden spit (Heyman et al. 2001) and undoubtedly large piscivores are also present, however the shear number of fish (c. 2000) attending those aggregations may facilitate daytime spawning by minimizing the dangers of adult and larval predation.
Cubera snapper aggregation formation at Gladden Spit is hypothesized to correlate to annual rising water temperature and increasing photoperiod (Heyman et al. 2005). In Belize this generally begins in March and lasts through September. Aggregation sizes peak in May and June. Off Cuba, aggregations are reported to occur from June through September (Claro and Lindeman 2003) and in the Florida Keys from May through August (Lindeman et al. 2000). On the Grammanik bank aggregations of up to 1000 cubera snapper were observed between May and August from 2003 to 2005 and smaller numbers of scattered fish (< 150) were seen during the months of March, April and/or May. The water temperature data collected lend support to Heymans rising water temperature hypothesis. In 2004 and 2005 cubera snapper began appearing on the bank when the monthly mean water temperature began increasing and aggregation formation occurred from 2003- 2005 when mean water temperature rose higher than 26.9°C. The slightly protracted spawning season in Belize may be a function of warmer, more favorable water temperature earlier in the year (mean monthly water temperature in March on Gladden Spit was higher than 26.5ºC every year for the six year study period) or again may be because of the large number of cubera snapper spawning in that region of the Mesoamerican reef. The scattered smaller groups of fish that we observed in the months just prior to aggregation formation may have been prepared to spawn but numbers may have been too low to form aggregations.
Similar to cubera spawning periodicity in Belize (Heyman et al. 2005), Cuba (Claro and Lindeman 2003) and the Florida Keys (Lindeman et al. 2000), aggregation formation on the Grammanik bank appears to be in accordance to the lunar phase and occurs on and after the full moon (Fig. 3). Aggregations were observed fairly consistently on all dives made on the full moon and up to one day after the third quarter moon but on multiple dives made within 4 days around the new moon in May and June of 2005 were generally absent from the bank.
Spatial dynamics: Cubera snapper studied on Gladden Spit from 1998-2003 exhibited extreme site fidelity with aggregations observed consistently in an area of < 1000 m2 (Heyman et al. 2005). In contrast the aggregations on the Grammanik bank moved west from 2003 to 2005, utilizing 600 linear meters or more of the 100 m wide bank in an area > 60 000 m2. In addition to the top of the reef, the aggregation was seen on one occasion south of the bank in greater than 45 m of water. The Grammanik bank lies on a shelf edge which is fairly uniform along its east-west axis and the benefits derived from spawning on the promontory formation described at Gladden Spit may be lacking. Without the obvious bathymetric landmarks present on the reef in Belize, the snapper on the Grammanik bank may employ a more nomadic behavior, moving up and down the reef to collect aggregation participants. In support of this hypothesis is the initial reaction we got from snapper during most of our dives. Cubera snapper nearly always met divers descending in the water column, approaching aggressively and spending a considerable amount of time circling closely. During many dives we observed fish moving swiftly across the reef in seemingly random directions, merging into larger aggregations or splitting into smaller groups. Although the main aggregations appeared consistently during a season on one area of the reef, that area was variable between years. Although this may have been a function of prevailing currents, the bathymetry and bank orientation are fairly consist across the Grammanik bank. We believe that the area utilized by the cubera snapper may be fairly random on the reef and more dependent on immediate fish densities and social interactions. These factors may play as vital a role in reproduction as habitat and physical oceanographic parameters (Coleman et al. 1996, Vincent and Sadovy 1998).
Dog snapper: The dog snapper aggregation on the Grammanik bank was not observed the first two seasons of sampling. It is unlikely that it formed for the first time in February of 2005 but instead was probably missed on prior surveys due to the compact nature of the aggregation or it s location outside the immediate survey area. The dog snapper aggregation was found over the sand channel north of the fore-reef and on the edge and over the adjacent northern reef running parallel to the southern bank. The site is less than 300 m from the grouper aggregation area, but was not surveyed completely in 2003 and 2004. In addition, annual surveys started in the month of March in 2003 and 2004 while the largest dog snapper aggregations were observed in February of 2005.
We found a relation between the presence of dog snapper aggregations and lunar phase on the Grammanik bank with densities peaking between the full and third quarter moon. Regionally the timing appears inconsistent both in season and lunar phase. At Gladden Spit dog snapper aggregations peak around the full and third quarter moon in April and May (Heyman et al. 2001), while observations on probable dog snapper aggregations off English Cay, Belize were just after the full moon in July (Domeier and Colin 1997), off Cay Glory three days after the new moon in January (Carter and Perrine 1994) and in the Cayman Islands between the full and third quarter moon in January (Whaylen et al. 2004). Fisheries data supports spawning in summer off of Rileys Hump in the Dry Tortugas (Lindeman et al. 2000) and year round on the full moon off much of the coast of Cuba (Claro and Lindeman 2003). The only published account of observed dog snapper spawning is from Gladden Spit in Belize (Heyman et al. 2001). Observations from English Cay (Domeier and Colin 1997) and Cay Glory (Carter and Perrine 1994) culminated in tight balls of fish within aggregations made of up to 1000 individuals, all rising toward the surface together and then slowly descending. This is very similar to the behavior we observed in the afternoon. On dives made earlier in the day fish were less active, swimming across the bottom in smaller groups with little vertical movement. The change in behavior later in the day, with no actual spawning observed, indicated that these fish may also be spawning after sunset.
Management and Conservation: Grouper and snapper spawning aggregations are highly vulnerable to extirpation and should be managed and protected. Beginning in 2005 the Grammanik bank was closed seasonally from February through May to fishing (Federal Registry Vol. 70 No. 2 and 208) due to data collected by Nemeth et al. (2004) confirming the presence of spawning aggregations of yellowfin grouper, Mycteroperca venenosa (Linnaeus, 1758), tiger grouper, M. tigris (Valenciennes 1833) and Nassau grouper during those months. With the discovery of large snappers utilizing the bank for spawning as well, its overall importance as critical fish habitat was further highlighted. Although the current status of both cubera and dog snapper is unknown in the U.S. Virgin Islands due to lack of fishery data, their populations and aggregations are functionally protected from over-exploitation by the believed presence of ciguatoxins in the tissues. There is controversy over whether or not additional legal protection is warranted. In support of a permanent year-round closure of the Grammanik bank is the protection the coral reef habitat would receive from destructive fishing gear and anchors (Appeldorn et al. 2000, Quant 1999) that are now used on the bank. In addition, a precautionary measure to protect the cubera snapper from potential exploitive threats such as new non-consumptive markets would be in place.
Resumen
El banco Grammanik es un arrecife profundo (35-40 m) ubicado aproximadamente 14 km al Sur de St. Thomas (Islas Vírgenes EUA), en el límite sur de la plataforma de Puerto Rico. Este arrecife relativamente angosto y de 1.5 km de longitud, recibe anualmente agrupaciones de varias especies de meros, pargos y jureles, durante sus desoves. Fue objeto de explotación de moderada a alta por parte de los pescadores hasta febrero del 2005, cuando el Consejo de Pesca del Caribe le ofreció protección temporal. El pargo cubera (Lutjanus cyanopterus) fue observado en el banco Grammanik en agrupaciones numerosas entre el año 2003 y el 2005, a su vez, agrupaciones del pargo perro o pargo caucha (L. jocu) se observaron en el 2005. La llegada y partida de estos grupos parece ser estacional y estar relacionada con la fase lunar. El desove estacional del pargo cubera coincide con informes de otros sitios del Caribe y al Sur de la Florida, donde se observaron cardúmenes de hasta 1000 peces de cada especie, así como comportamientos previos al desove y liberación de esperma. Agrupaciones de ambas especies mostraron alta fidelidad al sitio en una misma estación, pero los sitios de agrupación del pargo cubera cambiaron entre estaciones. Los autores recomiendan el cierre del banco Gammanik durante todo el año, basados en su importancia como hábitat crítico para los peces y por ser un área de agrupación para múltiples especies.
Palabras clave: agrupaciones de desove, pargo cubera, pargo perro o caucha, Lutjanidae, Áreas Marinas Protegidas (AMP).
References
Aguilar-Perera, A. & W. Aguilar-Davila. 1996. A spawning aggregation of Nassau grouper Epinephalus striatus in the Mexican Caribbean. Env. Biol. Fish. 45: 351-361. [ Links ]
Allen, G. R. 1985. FAO Species Catalogue. Vol. 6. An annotated and illustrated catalogue of lutjanid species known to date. FAO Fish. Synop. 125. 208 p. [ Links ]
Appeldorn, M., R. S. Nemeth, J. Vasslides & M. Scharer. 2000. The effect of fish traps on benthic habitats off La Parguera, Puerto Rico. Dept. Mar. Sci., Univ. Puerto Rico, Mayaguez, Puerto Rico. 29 p. [ Links ]
Beets, J. & A. Friedlander. 1992. Stock analysis and management strategies for the red hind, Epinephelus guttatus, in the US Virgin Islands. Proc. Gulf and Carib. Fish. Inst. 42: 66-80. [ Links ]
Bohnsack, J. A. 1998. Protection of grouper spawning aggregations. Nat. Mar. Fish. Serv., Coastal Resource Div., Contrib. No. CRD-88/89-06. 8 p. [ Links ]
Burton, M. L., K. J. Brennan, R. C. Munoz & R. O. Parker Jr. 2005. Preliminary evidence of increased spawning aggregations of mutton snapper (Lutjanus analis) at Rileys hump two years after the establishment of the Tortugas South Ecological Reserve. Fish. Bull. 103: 404-406. [ Links ]
Carter, J. & D. Perrine. 1994. A spawning aggregation of dog snapper, Lutjanus jocu (Pisces: Lutjanidae) in Belize, Central America. Bull. Mar. Sci. 55: 228- 234. [ Links ]
Carter, J., G. J. Marrow & V. Pryor. 1994. Aspects of the ecology and reproduction of the Nassau grouper, Epinephalus striatus, off the coast of Belize, Central America. Gulf. Carib. Fish. Inst. 43: 64-11 [ Links ]
Claro, R. & K. C. Lindeman. 2003. Spawning aggregation sites of snapper and grouper (Lutjanidae and Serranidae) species on the insular shelf of Cuba. Gulf Carib. Res. 14: 91-106. [ Links ]
Coleman, F., C. Koenig & A. Collins. 1996. Reproductive styles of shallow-water groupers (Pisces: Serranidae) in the eastern Gulf of Mexico and the consequences of fishing spawning aggregations. Env. Biol. Fish. 47: 129-141. [ Links ]
Colin, P. L. 1992. Reproduction of the Nassau grouper, Epinephalus striatus (Pisces: Serranidae) and its relationship to environmental conditions. Env. Biol. Fish. 34: 357-377. [ Links ]
Colin, P. L. 1996. Longevity of some coral reef fish spawning aggregations. Copeia 1996: 189-191. [ Links ]
Colin, P. L., D.Y. Shapiro & D. Weiler. 1987. Aspects of the reproduction of two groupers, Epinephalus guttatus and E. striatus in the West Indies. Bull. Mar. Sci. 40: 220-230. [ Links ]
Domeier, M. L. & P. L. Colin. 1997. Tropical fish spawning aggregations: defined and reviewed. Bull. Mar. Sci. 60: 698-726. [ Links ]
Gobert, B., P. Berthou, E. Lopez, P. Lespagnol, M. D. O. Turcios, C. Macabiau & P. Portillo. 2005. Early stages of snapper-grouper exploitation in the Caribbean (Bay Islands, Honduras). Fish. Res. 73: 159-169. [ Links ]
Heyman, W. D. & N. Requena. 2002. Status of Multi-Species Spawning Aggregations in Belize. The Nature Conservancy. Punta Gorda, Belize. 27 p. [ Links ]
Heyman, W. D., B. Kjerfve, R. T. Graham, K. L. Rhodes & L. Garbutt. 2005. Spawning period. J. Fish Biol. 67: 83-101. [ Links ]
Heyman, W. D., R. T. Graham, B. Kjerfve & R. E. Johannes. 2001. Whale sharks Rhincodon typus aggregate to feed on fish spawn in Belize. Mar. Ecol. Prog. Ser. 215: 275-282. [ Links ]
Johannes, R. E. 1978. Traditional marine conservation methods in Oceania and their demise. Ann. Rev. Ecol. Syst. 9: 349-364. [ Links ]
Johannes, R. E., L. Squire, T. Graham, Y. Sadovy & H. Renguul. 1999. Spawning aggregations of groupers (Serranidae) in Palau. The Nat. Conserv. Mar. Ser. Publ. No. 1. 144 p. [ Links ]
Lindeman, K. C., R. Pugliese, G. T. Walsh & J. Ault. 2000. Developmental patterns within a multispecies reef fishery: management applications for essential fish habitats and protected areas. Bull. Mar. Sci. 66: 929-956. [ Links ]
Luckhurst B. E. 1998. Site fidelity and return migration of tagged red hind (Epinephelus guttatus) to a spawning aggregation site in Bermuda. Proc. Gulf Carib Fish Inst. 50: 750-763. [ Links ]
Luckhurst, B. E. 2003. Development of a Caribbean Regional Conservation Strategy for reef fish spawning aggregations. Proc. Gulf Carib. Fish. Inst. 54: 668-679. [ Links ]
Nemeth, R. S. 2005. Population characteristics of a recovering US Virgin Islands red hind spawning aggregation following protection. Mar. Ecol. Prog. Ser. 286: 81-97. [ Links ]
Nemeth, R. S., E. Kadison, S. Herzlieb, J. Blondeau & E. Whiteman. 2004. Status of yellowfin (Mycteroperca venenosa) and Nassau (Epinephalus striatus) grouper spawning aggregations in the US Virgin Islands with notes on other species. Proc. 57 th GCFI, St. Petersburg, Florida, USA. [ Links ]
Olsen, D. A. & J. A. LaPlace. 1978. A study of a Virgin Islands fishery based on a breeding aggregation. Proc. Gulf Carib. Fish. Inst. 31: 130-144. [ Links ]
Quant, A. 1999. Assessment of trap damage on coral reefs around St. Thomas USVI. University of the Virgin Islands. St. Thomas, USVI. 14 p. [ Links ]
Sadovy, Y. 1994. Grouper stocks of the western central Atlantic: the need for management and management needs. Proc. Gulf Carib. Fish. Inst. 43: 43-65. [ Links ]
Sadovy, Y. 1997. The case of the disappearing grouper: Epinephelus striatus, the Nassau grouper in the Caribbean and western Atlantic. Proc. Gulf Carib. Fish. Inst. 43: 43-65. [ Links ]
Sadovy, Y & P. L. Colin. 1995. Sexual development and sexuality in the Nassau grouper. J. Fish Biol. 46: 961-976. [ Links ]
Shapiro, D. Y., Y. Sadovy & M. A. Mcgehee. 1993a. Size, composition and spatial structure of the annual spawning aggregation of the red hind, Epinephelus guttatus (Pisces: Serranidae). Copeia 1993: 399-406. [ Links ]
Shapiro, D. Y., Y. Sadovy & M. A. Mcgehee. 1993b. Periodicity of sex change and reproduction in the red hind (Epinephelus guttatus) a protogynous grouper. Bull. Mar. Sci. 53: 399-406. [ Links ]
Thompson, R. & J. L. Munro. 1983. The biology, ecology and bionomics of the snappers, Lutjanidae, p. 94-109. In J. L. Munro (ed.). Caribbean Coral Reef Fisheries Resources. ICLARM, Manila, Philippines. [ Links ]
Tucker, J. W., P. G. Bush & S. T. Slaybaugh. 1993. Reproductive patterns of Cayman Islands Nassau grouper (Epinephelus striatus) populations. Bull. Mar. Sci. 52: 961-969. [ Links ]
Vincent, A. & Y. Sadovy. 1998. Reproductive ecology in the conservation and management of fishes, p. 209- 245 In T. Claro (ed.). Behavior ecology and conservation biology. Oxford Univ. Press, New York. [ Links ]
Whiteman, E., C. A. Jennings & R. S. Nemeth. 2005. Sex structure and potential female fecundity in an Epinephalus guttatus spawning aggregation: applying ultrasound imaging. J. Fish Biol. 66: 983-995. [ Links ]
Whaylen, L., C. V. Pattengill-Semmens, B. G. Bush & M. R. Boardman. 2004. Observations of a Nassau grouper (Epinephelus striatus) spawning aggregation site in Little Cayman, including multi-species spawning information. Env. Bio. Fish. 70: 305-313. [ Links ]

