











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.50 no.2 San José jun. 2002
to adjacent pastures on the western slope of Volcán Barva, Costa Rica
Gilbert Barrantes and Ana Pereira
Museo de Zoología, Escuela de Biología, Universidad de Costa Rica, San José, Costa Rica.
Received 28-III-2002. Corrected 30-X-2002. Accepted 27-II-2002.
Abstract
Logging, cattle raising, and agricultural activities have caused the destruction of most forested areas in Costa Rica. In some middle and highlands the abrupt topography delayed the complete destruction of montane forest. Consequently, some fragments of almost pristine forest remain along streams that run in deep canyons. Frequently, these remnants serve as corridors between larger forested areas and as routes for movement of frugivorous birds. Eighteen bird species, e.g., Turdus plebejus, Elaenia frantzii and Ptilogonys caudatus are common dwellers of forest patches throughout the Pacific slope of the Volcán Barva. These species fly frequently from forest fragments to adjacent pastures. They defecated and regurgitated seeds of 28 plant species on stumps scattered on pasture areas. Isolated trees and specially the stumps are suitable microhabitats for germination of seeds and establishment of seedlings.
Key words: Deforestation, ornithocory, natural regeneration.
Deforestation in Costa Rica has increased steadily during the last half century as result of agricultural activities, cattle raising, and specially, logging activities (Sader and Joyce 1988, Rosero-Bixby and Palloni 1998). These human activities have devastated a large proportion of formerly forested areas of this country. Consequently, pristine forests have quickly been converted into islands contained in a matrix of agricultural systems (Ewel 1983). Many areas dedicated to agriculture or cattle raising turned out to be inappropriate for such activities due to local climate and soil conditions. In some cases, such areas are allowed to revert to natural conditions as it is occurring with pastures in some neotropical areas (Ewel 1983).
Several practices have been implemented to expedite the natural processes of regeneration in tropical ecosystems (Uhl and Jordan 1984, Janzen 1988a, b, Uhl 1988, Virgílio et al.1997). One approach that has been frequently proposed is the natural regeneration that involves protection of the deforested area from further disturbances to allow natural successional processes to restore forest communities (Fournier 1977, Lamb et al. 1997). The natural recovery of an ecosystem usually takes very long. One of the causes of such a delay is the low availability of propagules and non-availability of suitable microhabitats for plant establishment (Janzen 1988a, b, Uhl 1988).
Restoration time of natural forests may be reduced by introducing seeds or propagules of certain species that modify the environment and facilitate establishment of other species (Sun et al. 1995). Planting particular trees is another technique that usually accelerates natural regeneration. There are several plant species that tolerate and ameliorate the adverse conditions prevailing in degraded ecosystems (Lamb et al. 1997). Other species not only tolerate such conditions but accelerate the colonization process by attracting seed-dispersing animals (Lugo 1988, Lugo et al. 1993, Parrota 1992). In this study we evaluated the role of stumps in pastures surrounded by forest remnants as collectors of bird-dispersed seeds and appropriate microhabitats for seedling establishment.
Materials and methods
We conducted this research from February through December 1986, on the western slope of the Volcán Barva in an area of ca. 400 ha (84º07|W, 10º09|N). The study site, located at 1|800|m, consisted of patches of remnant montane forest (Holdridge 1967), mostly along small streams, immersed into a pasture matrix.
We gathered bird feces from 11 stumps scattered throughout the pasture area every two weeks. We counted and identified (if possible) the seeds present in the feces. We also censused frugivorous birds and recorded number and species of fruits eaten by these birds along a 2|km transect at forest remnant edges. Changes in abundance of some seed species in fecal samples were analyzed using one sample Kolmogorov-Smirnov test. We also evaluated the association between amount of seed species in fecal samples and abundance of some bird species with Spearman correlation tests.
Results
We registered 18 frugivorous birds in the study site. Based on their habitat use, we divided these species into three categories (Table 1). First, a group of birds that consisted of those species, such as Pharomachrus mocinno, Aulacorhynchus pracinus, and Myadestes melanops, whose activity concentrated in forest fragments. These species were not observed on pastures and seldom at the edge of forest fragments. Other species, e.g., Catharus ustulatus and Chlorospingus ophthalmicus, that exploited intensely forest edges and often flew between forest patches and isolated trees, constituted the second group. The last group is formed by birds that concentrated their activities either permanently or temporarily in pastures (scattered trees) and forest edges. Some of these birds, e.g., Thraupis episcopus and Turdus grayi, foraged commonly in pastures over all the study period, whereas other birds, e.g., Turdus plebejus and Ptilogonys caudatus, used intensely the open areas only when some isolated trees (e.g., Cornus disciflora) produced large fruit crops.
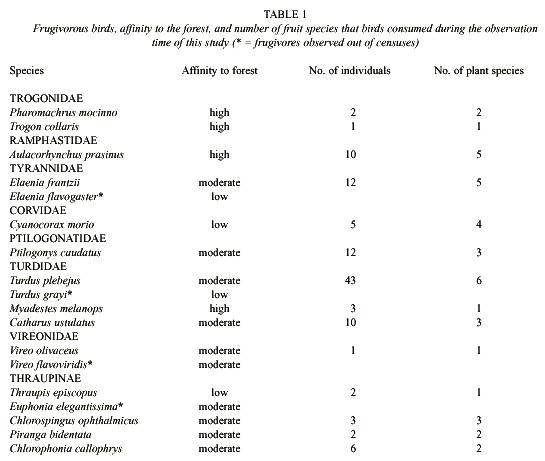
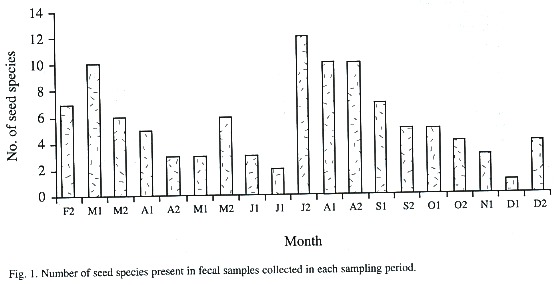
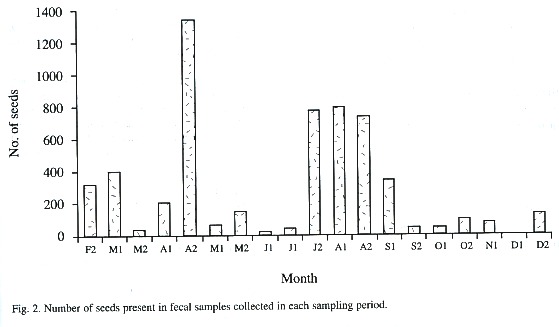
We collected 133 feces from which we identified 28 seed species (Table 2). The number of species of seeds disseminated from forest patches to pastures did not vary significantly along the study period (Dmax = 0.1, p > 0.20, n = 19, Fig. 1). Ficus sp., followed by the unknown species 3, Cornus disciflora, and Rubus sp., appeared in most of the surveys (Table 2). Yet, the amount of these seed species varied greatly among samplings. Ficus sp. occurred in a very low proportion in February, April, and July, whilst, in May, August, and October, the proportion of this species increased significantly (Dmax = 0.54, p = 0.05, n = 13). Cornus disciflora was proportionally greater in September and October (Dmax = 0.88, P < 0.001, n = 8), and Rubus sp. in March and September (Dmax = 0.88, p < 0.001, n = 8). This intraspecific fluctuation affected the total amount of seeds counted biweekly from fecal samples (Fig. 2).
We registered 14 birds during the study period feeding on fruits of 19 plant species in forest fragments (Table 3). Only seven out of these 19 seed species appeared in the fecal samples. Even some species on which birds fed heavily, e.g., Trema micrantha, and Oreopanax xalapense were not present on the feces collected. This showed either that some of the birds may not fly from fragments to nearby pastures or they did not defecate on stumps. Overall, the number of feces gathered on stumps was not directly related with the number of frugivorous birds counted on each sampling (rs = 0.3, p = 0.2, n = 19 ). We detected, however, a highly significant association between the number of fecal samples and abundance of Turdus plebejus during this study (rs = 0.61, p = 0.0005, n = 11). The importance of this species as possible disperser is supported by the fact that 38% of the birds observed feeding upon any type of fruit belong to this species (Table 1).

We also found several seedling species already established around the stumps. These seedlings mostly corresponded to the same species of plants dispersed by birds (Table 2). The establishment of seedlings around stumps supports the idea that these structures not only function as seed collectors but also proportionate an adequate microhabitat for seedling establishment.
Discussion
The number of seeds that each fruit contain varies enormously among plant species. For instance, each Cornus disciflora and Trichilia glabra fruit possesses few seeds (one and three respectively), whereas fruits of Blakea gracilis and Conostegia oerstediana contain a large number of seeds (more than 100). This difference in number of seeds per fruit determines, to a large extent, the fluctuation in seed species and seed number over the study period (Fig. 2). Other factors, such as crop size, fruiting period, and synchrony in fruit production, also affect availability of this resource and, therefore, its presence in the fecal samples (Howe and Primak 1975, Herrera 1981, Moermond and Denslow 1983, Levey et al. 1984).
Species-specific behavior and pattern of habitat use of some birds affect seed dispersion from forests to open areas (Herrera 1984, Wheelwright 1985, Moermond and Denslow 1985). Turdus plebejus, an altitudinal migrant, likely plays an important role in the mobilization of seeds from forest patches to adjacent pastures at middle-high elevations of the western slope of Costa Rican cordilleras. This species frequently flies back and forth between forest and isolated trees in pastures where it feeds on fruits. It is also common that T. plebejus perches and defecates on stumps (pers. obs.). Seeds are washed out by rainfall and deposited around the stumps where they germinate, relatively well protected from cattle grazing. By using a large variety of fruits, T. plebejus may be an excellent disperser in this environment. The variety of fruits that T. plebejus consumes makes it an important element in the natural regeneration of forests at middle and high elevations in Costa Rica. It feeds upon fruits of herbs (e.g., Rubus sp. and Phytolacca rugosa), bushes (e.g., Parathesis storki and Solanum sp.), and trees (e.g., Cornus disciflora and Ficus sp.). Ptilogonys caudatus, another altitudinal migrant, also moves frequently between forest fragments and isolated trees in pastures. Yet, this species does not perch on stumps in open areas, so that most of the seeds are likely dropped beneath the crown of trees. Other bird species may play a much less important role on seed dispersion at the west slope of the Volcán Barva.
Acknowledgments
We thank J.M. Mora and anonymous reviewers for valuable comments on the manuscript.
Resumen
La deforestación, la ganadería y las actividades agrícolas han causado la destrucción de la mayoría de los bosques en Costa Rica. Sin embargo, la abrupta topografía de las zonas medias y altas del país ha retardado este proceso de deforestación en el bosque montano. Es así como aún algunos fragmentos de bosques poco alterados estan todavía en pie a lo largo de riachuelos que corren en cañones profundos. Estos fragmentos sirven como corredores entre áreas más grandes de bosque y como rutas para el movimiento altitudinal de aves frugívoras. Dieciocho aves, e.g., Turdus plebejus, Elaenia frantzii y Ptilogonys caudatus son habitantes comunes de los parches de bosque presentes en la vertiente Pacífica del Volcán Barva. Estas especies vuelan frecuentemente entre los fragmentos de bosque y potreros adyacentes. Estas aves defecaron y regurgitaron semillas de 28 especies de plantas en troncos distribuidos en los potreros. Los árboles aislados y troncos son micro-hábitats adecuados para la germinación de semillas y el establecimiento de plántulas.
References
Ewel, J. 1983. Succession, p. 217-223. In F.B. Golley (ed.). Tropical rain forest ecosystems. Structure and function. Elsevier, New York. [ Links ]
Fournier, L.A. 1977. La sucesión ecológica como un método eficaz para la recuperación del bosque en Costa Rica. Agron. Costarricense 2: 53-55. [ Links ]
Janzen, D.H. 1988a. Management of habitat fragments in a tropical dry forest: Growth. Ann. Miss. Bot. Garden 75: 105-116. [ Links ]
Janzen, D.H. 1988b. Tropical dry forests. The most endangered major tropical ecosystem, p. 130-137. In W.O. Wilson (ed.). Biodiversity. National Academy, Washington, D. C. [ Links ]
Lamb, D., J. Parrota, R. Keenan & N. Tucker. 1997. Rejoining habitat remnants: Restoring degradated rainforest lands, p. 366-385. In W.F. Laurence & R.O. Bierregaard, Jr. (eds.). Tropical forest remnants. Ecology, management, and conservation of fragmented communities. University of Chicago, Chicago. [ Links ]
Lugo, A.E. 1988. The future of the forest: Ecosystem rehabilitation in the tropics. Environment 30: 16-20. [ Links ]
Lugo, A.E., J.A. Parrota & S. Brown. 1993. Loss in species caused by tropical deforestation and their recovery through management. Ambio 22: 106-109. [ Links ]
Herrera, C.M. 1981. Fruit variation and competition for dispersers in natural populations of Smilax aspera. Oikos 36: 51-58. [ Links ]
Herrera, C.M. 1984. A study of avian frugivores, bird-dispersed plants, and their interaction in Mediterranean scrublands. Ecol. Monogr. 54: 1-23. [ Links ]
Holdridge, L.R. 1967. Life zone ecology. Tropical Science Center, San José, Costa Rica. 216 p. [ Links ]
Howe, H.F. & R.B. Primak. 1975. Differential seed dispersal by birds of the tree Casearia nitida (Flacourtiaceae). Biotropica 7: 278-283. [ Links ]
Levey, D.J., T.C. Moermond & J.S. Denslow. 1984. Fruit choice in neotropical birds: The effect of distance between fruits on preference patterns. Ecology 65: 844-850. [ Links ]
Moermond, T.C. & J.S. Denslow. 1983. Fruit choice in netropical birds: Effects of fruit type and accessibility on selectivity. J. Anim. Ecol. 52: 407-420. [ Links ]
Moermond, T.C. & J.S. Denslow. 1985. Neotropical avian frugivores: Patterns of behavior, morphology, and nutrition with consequences for fruit selection. Ornithol. Mongr. 36: 865-897. [ Links ]
Rosero-Bixby, L. & A. Palloni. 1998. Población y deforestación en Costa Rica, p. 131-150. In Conservación del bosque en Costa Rica. Academia Nacional de Ciencias, Grafos S. A., San José, Costa Rica. [ Links ]
Parrota, J.A. 1992. The role of plantation forests in rehabiltating degraded ecosystems. Ag. Ecosyst. Environ. 41: 115-133. [ Links ]
Sader, S.A. & A.T. Joyce. 1988. Deforestation rates and trends in Costa Rica, 1940 to 1983. Biotropica 20: 11-19. [ Links ]
Sun, D., G. Dickenson & A. Bragg. 1995. Direct seeding of Alphitonia peteri for gully revegetation in tropical Northern Australia. Forest Ecol. Manage. 73: 249-257. [ Links ]
Virgílio, M.V., A.A. J. Tabanez & J.L.F. Batista. 1997. Dynamics and restoration of forest fragments in the Brazilian Atlantic moist forest, p. 351-365. In W.F. Laurence & R.O. Bierregaard, Jr. (eds.). Tropical forest remnants. Ecology, management, and conservation of fragmented communities. University of Chicago, Chicago. [ Links ]
Uhl, C. 1988. Restoration of degraded lands in the Amazon Basin, p. 326-332. In W.O. Wilson (ed.). Biodiversity. National Academy, Washington, D. C. [ Links ]
Uhl, C. & C. Jordan. 1984. Succession and nutrient dynamics following forest cutting and burning in Amazonia. Ecology 75: 377-407. [ Links ]
Wheelwright, N.T. 1985. Fruit size, gape width, and the diets of fruit-eating birds. Ecology 66: 808-818. [ Links ]

