











 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkRevista de Biología Tropical
versión On-line ISSN 0034-7744versión impresa ISSN 0034-7744
Rev. biol. trop vol.49 no.2 San José jun. 2001
fragments in Brazil: A test of models
Samuel Jorge Leite1 Frederico Santos Lopes2
Received 3-V-2000. Corrected 1-XI-2000. Accepted 13-XI-2000.
Abstract
Patterns of local abundance and regional distribution of tree species were analyzed on three spatial scales and compared with those predicted by Hanski's 1982 original core-satellite model, Brown's 1984 nichebased model, and Tokeshi's 1992 model. Data were collected in seven forest fragments dispersed over approximately 7200 km2, remnants of a former continuous forest in the south of Mato Grosso do Sul state, southwestern Brazil. Fifty-six east-west-oriented 10 m ¥ 150 m plots were randomly demarcated in those fragments from 1994 to 1996. In each plot all trees having DBH ?0.15 m were recorded and measured. The smallest scale was that of the fragments themselves and the greatest was that of the entire region. No evidence was found of the bimodality predicted by Hanski's 1982 model on any scale. On all scales, an unimodal pattern was found whose mode was on the left side (satellite mode) of the distribution. This pattern was in best agreement with the predictions of Tokeshi's 1992 model.
Key words
Abundance, biogeography, core-satellite hypothesis, spatial distribution.
The relationship between species abundance and geographic distribution is a central issue in modern ecological studies (see Gaston 1990, 1996, Gaston and Lawton 1990, Brown et al. 1996, Gaston et al. 1997, for reviews). This relationship has important implications for the understanding of community structure and for the description of biodiversity patterns, as well as for proposing and testing hypotheses regarding the processes underlying these patterns (Brown and Maurer 1987, Hanski et al. 1993, Brown et al. 1995, Gaston and Blackburn 1996, Johnson 1998).
Gotelli and Simberloff (1987) recognized two kinds of models that relate geographic distribution and abundance: static and dynamic. In static models, distribution and abundance of species do not vary over time, except as a consequence of major climatic or habitat changes. In contrast, dynamic models posit that both the geographic range and the local abundance of a species vary over time.
Some static models, as that of Brown (1984), consider that the relationships between a species local abundance and its geographic distribution are based on its niche amplitude. Brown's (1984) model further predicts that both the abundance and the frequency of a species decrease from the center of its geographic distribution toward all its boundaries. In addition. Brown (1984) emphasized that the distribution of species by sites is usually unimodal, and suggested that a bimodal pattern, such as that proposed by Hanski (1982), is a sample artifact derived from a small number of samples on a very local scale. Scheiner and Rey-Benayas (1997), analyzing data for terrestrial vascular plants from 74 landscapes throughout the world, concluded that niche-based models are robust on scales greater than 1 km2, while current metapopulation models are not.
Levins's (1969) metapopulation model, which is dynamic, is not based on any consideration about niche, but on colonization and extinction rates instead. The metapopulation approach is directly related to the life history of organisms, their life span, fecundity, dispersar, and recruitment (Eriksson and Jakobsson 1998). Hanski's (1982) core-satellite metapopulation model, derived from Levins's model, applies to a suite of species where dispersar ability between sub-populations is high enough to allow all individuals to reach all sites in a region (Collins and Glenn 1991,1997; Scheiner and Rey-Benayas 1997). Another assumption of Hanski's (1982) model is that the environment should be nearly homogeneous for the set of species analyzed, which is a corollary of the assumption that only colonization and extinction rates determine the spatial distribution and abundance patterns of the species.
Results obtained by Scheiner and ReyBenayas (1997) and by Collins and Glenn (1997) suggested that scale might be a key factor in contrasting Brown's (1984) and Hanski's (1982) models. Most of the studies on this issue were done with small and short life cycles organisms, and very few tests were performed using long-lived and large-sized organisms. The reasons for this paucity are obviously the great effort and time needed to obtain data for organisms such as trees, for example. One of the few studies of this kind was that of Scheiner and Rey-Benayas (1997), on forests in Italy, South Aflica, and Ghana.
The Tokeshi's model is a variant of the island -biogeography model, in which the extinction rate is a decreasing function of the species frequency. The more widespread species tend to have a higher local abundance, with a positive correlation between frequency and abundance. The abundant species should a have a lower extinction rate than rare species. The result is a negative correlation of species frequency and extinction rate. This model assumes a positive correlation between frequency and abundance and predicts that species frequency distributions will either be flat or have a mode at or near 1, depending on immigration and extinction probabilities. The Tokeshi's model predicts as much the unimodality as the bimodality (Tokeshi 1992, Colins and Glenn 1997, Scheiner and ReyBenayas 1997).
In the present study, we examined data from 56 plots in seven forest fragments. Predictions of niche-based and metapopulation models were compared on three different spatial scales. Our aim was to investigate whether the patterns of distribution and abundance observed in differed among scales. On the smaller ones, we expected the environment heterogeneity to be also smaller and dispersar ability to be more homogeneous among species. lf this held true, the chances of finding patterns in agreement with Hanski's (1982) model would be improved. In contrast, on greater scales we expected a more pronounced influence of phenomena linked to the general geographic distribution of species, and therefore, a smaller chance of fínding the patterns predicted by Hanski (1982).
Materials and Methods
Study site: The semi-deciduous seasonal forests of Mato Grosso do Sul state are usually associated with the presence of fertile soils (IBGE 1992, Campos and Souza 1997). Its floristic structure is very similar to that of the Brazilian Atlantic Forest formation and presents three very well differentiated strata: two of them arboreal and one herbaceousshrubby (IBGE 1989, Souza et al. 1997). This physiognomy is related to the seasonal climate of the region, to the rainy, hot season occurring from October lo March, and to the dry and cold season extending from May lo September. This seasonality is responsible for the leaf fall in the arboreal stratum, which is particularly evident for individuals in the upper storey, whose heights range from 20 to 30 m (SEPLAN-MS 1989, IBGE 1992, Souza et al. 1997).
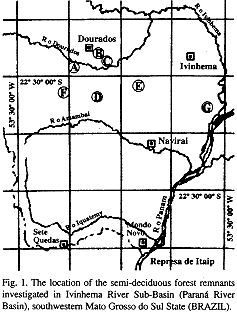
T'he study area lies in the south of Mato Grosso do Sul state, southwestern Brazil, in the sub-basins of the Dourados and Guiraí rivers (22°15'- 23°00'S, 53°30'- 55°30'W), within the basin of the Ivinhema, a tributary of the Paraná river (Fig. 1).
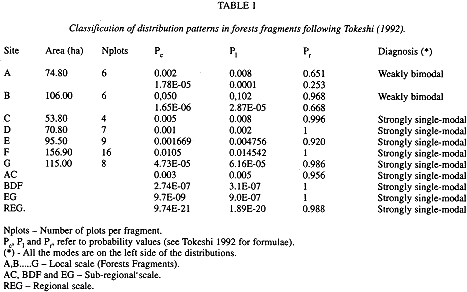
Methods and statistical analysis: Data were obtained from 1994 to 1996 in seven forest fragments located as far apart as 120 km. A total of 56 east-west-oriented 10 m x 150 m plots were randomly demarcated in those fragments. Each fragment contained from four to 16 plots, depending on its area. (see Table 1).
In each plot, all trees having DBH ?0.15 m (1.3 m from the ground or above the buttresses) were recorded. Trees were identified in accordance with Lorenzi (1992). From the field data, abundance and frequency, both of them regional, sub-regional and local, were calculated.
To test the patterns predicted by Brown's (1984) niche-based model, Hanski's (1982) core-satellite hypothesis and Tokeshi's model, the distribution and abundance of species were analyzed at three levels. The highest was that of the whole region composed of all seven fragments. At an intermediate, sub-regional level, the fragments were grouped according to their similarities in species composition and species abundance, using multivariate analysis (TWINSPAN and DCA) (Leite and Lopes, in prep.). The lowest level was that of each fragment.
For each species, the following parameters were employed: regional frequency (expressed as the ratio between the number of fragments where the species was found and the total number of fragments); sub-regional frequency (the ratio between the number of plots possessing that species and the total number of plots of a sub-region); and local frequency (the ratio between the number of plots possessing that species and the number of plots in a fragment). The parameter abundance was considered as the mean abundance of the plots where the species occurred.
Values thus obtained were tested for modality. Although no general tests for bimodality exist (Ellison 1993, in Scheiner and Ray-Benayas 1997), Tokeshi (1992) proposed a particular test that allows to assess the exact probability of some classes having more occurrences that those expected by chance.
Results
Regional frequency andlocalabundance: The seven forest fragments may be arranged in three groups (Leite and Lopes, in prep.). Fragments A and C constitute the first group, that of fragments of riparian forest; the other fivefragments make up a gradient ranging from the more humid conditions (fragments B, D, and F: the second group) to the drier ones (fragments E and G: the third group).
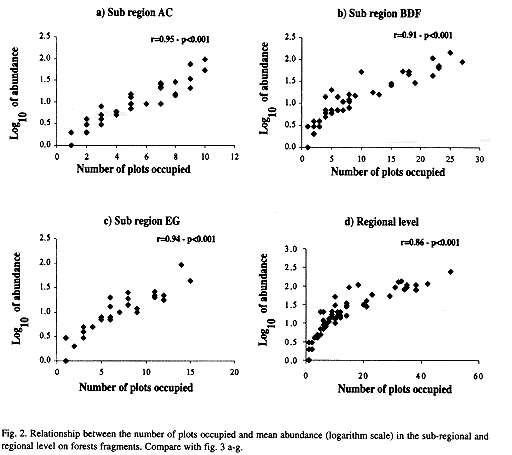
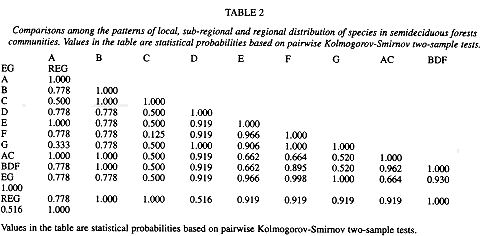
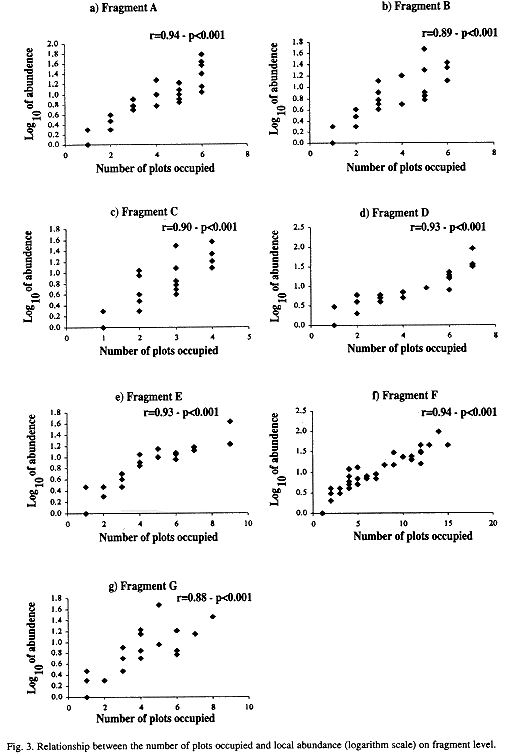
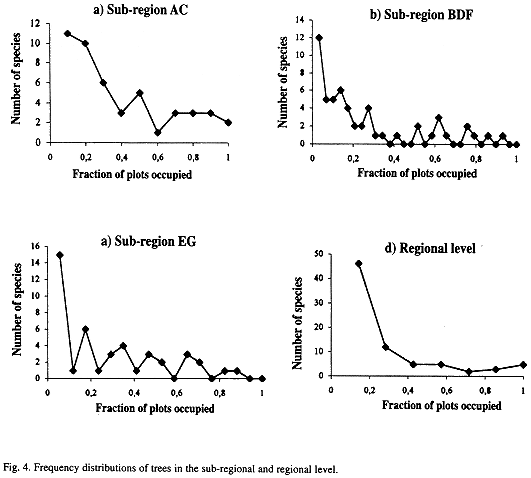
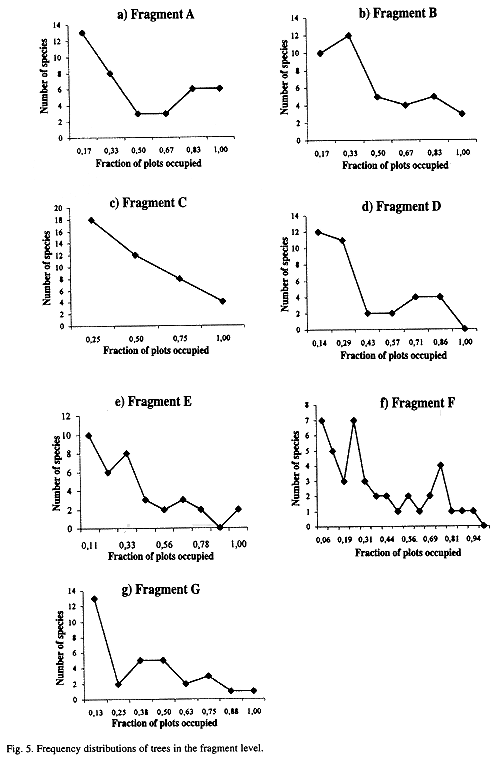
The relationship between number of plots and the logarithm of the species mean abundance presented a positive correlation at all three -regional, sub-regional, and fragment- levels (Figs. 2 a-d and 3 a-g). Relationships,between the fraction of occupied plots and the proportion of species for the whole region, for the three sub-regions, and for the fragments are shown in Figs. 4 a-b and 5 a-g. The data for bimodality were tested employing the procedure proposed by Tokeshi (1992), and the results obtained can be seen in Table 1. Only in fragments A and B we found some evidence for bimodality. However, when we compared all distributions to each other, using a pairwise Kolmogorov-Smirnov test (Table 2), no statistically significant differences were found among them.
Discussion
In the present study, no clear evidence was found for bimodality either on the regional or the sub-regional scales. On the local scale, the test proposed by Tokeshi (1992) revealed that five of the seven fragments presentes a strong single left mode. Only in fragments A and B was there any evidence of weak bimodality. However, when the distributions of those two fragments were compared with the other ones, no significant differences emerged. Furthermore, in all cases, regardless of scale, the mode was always on the left side of the distribution. Our data therefore suggest that for tree communities of this region the bimodality expected from Hanski's (1982) core-satellite model, in which the right mode is greater than the left one, does not occur on any scale. Also, no evidence was found for the central unimodality suggested by Gotelli and Kelley (1993).
Brown (1984), criticizing the Hanski's (1982) model, supposed that bimodality, where found, would be a consequence of sampling artifact. According to him, if a small number of sites in a local region are sampled, the most widespread and abundant species will constitute a relatively high proportion of the sampled species living there. On greater scales, the number of the most widespread and abundant species remains approximately constant, but throughout the entire region many new rare species will be sampled. As a consequence, on smaller scales may there might be a mode on the right side of the distribution, while on larger scales bimodality would not occur.
Furthermore, as Scheiner and Rey-Benayas (1997) had already suggested, at least on scales greater than 1 km2 (100 ha), Brown's (1984) model is robust, while models based on metapopulation dynamics are not. The former investigators, therefore, did not discard the possibility that metapopulation models may be robust on smaller scales. All our seven forest fragments were smaller than 160 ha and three were smaller than 100 ha, thus having nearly the minimum size of those sites studied by Scheiner and Rey-Benayas. In spite of this, our data did not show any evidence of bimodality.
Our results, however, are diose expected by employing the metapopulation model proposed by Tokeshi (1992), which predicts the occurrence of both unimodality and bimodality. Additionally, Tokeshi (1992) criticized Hanski's (1982) model because its predictions assume a very high variance in the value of the difference between parameters i (colonization parameter) and e (extinction parameter). In other words, only if stochasticity is very high does Hanski's model predict a bimodal pattern. Tokeshi's model, in turn, does not require such an assumption.
Collins and Glenn (1997), comparing different spatial scales for four groups of organisms (grasshoppers, small mammals, birds, and herbaceous plants), found that on local scale grasshoppers and small mammals presented a signifícant bimodality, with a core mode greater than the satellite mode. Such a pattern, however, was not observed for birds and plants. For all these four groups the core mode decreased on the greatest scale and the pattern tended to unimodality on the satellite mode, except for grasshoppers, which presented a significant intermediate second mode.
Collins and Glenn's (1997) data suggest that, for different kinds of organisms, different patterns of distribution and abundance should be expected on different scales, and that such differences are caused by differences in the dispersal ability of the organisms. The larger the region being considered, the greater the dispersal ability required of organisms to conform to the core-satellite predictions.
Plant dispersar ability, on average, is not as great as that of free-living animals. If Collins and Glenn (1997) are right, one should expect the predictions of die core-satellite model to be verified for plants on very local scales. However, their data presented unimodality on local scale for the prairie plants studied by them, as did our data for trees. Furthermore, these authors suggested that longer-living organisms are more likely to conform lo the core-satellite predictions than short-living ones with the same dispersal abilities, and that this occurs because chance dispersal events in a long-living organism can be compounded over time. Trees, nonetheless, are very long-living organisms, but our data did not show any evidence that could reinforce the predicted bimodality.
It is not possible lo separate the predictions of Brown's (1984) model from those of Tokeshi's (1992) only on the basis of the frequency distribution of species by sites and its abundances. Although Scheiner and Rey-Benayas (1997) have concluded that Brown's (1984) model is robust on greater scales, they tested it only against the species frequency distribution by sites. Another prediction of Brown's (1984) model is that the species abundance and frequency should increase toward the center of their geographical distribution, decreasing toward the boundaries.
Tokeshi's (1992) model, in tum, is more parsimonious that Brown's (1984) because it requires oniy two parameters (extinction and immigration). So, despite our lack of data on the geographical distribution of the species here considered, we can conclude that Tokeshi's (1992) metapopulation model is more robust than both Hanski's (1982) and Brown's (1984) models in describing patterns of frequency and abundance for tree species.
Acknowledgments
This paper resulted from the program "Padrões de Diversidade de Fauna e Flora do Pantanal Sul-Mato-Grossense" (CNPq process 521746/97-3 (NV)), CNPq grant (process 351220/97.6). Tle authors thank Júlio Louzada, Og F.F. de Souza and Rosane Collevatti for their helpful comments on the manuscript.
Resumen
Los patrones de abundancia local y la distribución regional de árboles fueron analizados en tres escalas espaciales y comparados con los modelos de Hanski 1982, núcleo-satélite, el modelo basado en el nicho de Brown 1984, y el modelo de Tokeshi 1992. Los datos fueron recolectados en siete fragmentos remanentes de bosque, distribuidos en aproximadamente 7 200 km2, en Mato Grosso do Sul, suroeste de Brasil. De 1994 a 1996, se demarcó aleatoriamente, en estos fragmentos, cincuenta y seis unidades de muestras, orientados en el sentido este-oeste, con 10 m x 150 m. En cada unidad se midió todos los árboles con DAP ?0.15 m. La menor escala fue el propio fragmento y la mayor escala toda la región. No encontramos evidencias de la bimodalidad prevista por el modelo de Hanski 1982. En todas las escalas se encontró un patrón unimodal, con moda a la izquierda de la distribución (moda satélite). El patrón observado en estos remanentes se ajustó más bien a las previsiones del modelo de Tokeshi 1992.
References
Brown, J.H. 1984. On the relationship between abundance and distribution of species. Am. Nat. 124: 255-279. [ Links ]
Brown, J.H. & B.A. Maurer. 1987. Evolution of species assemblages: effects of energetic constrains and
species dynamics on the diversification of the north American avifauna. Am. Nat. 130: 1-17
Brown, J.H., D.W. Mehlman & G.C. Stevens. 1995. Spatial variation in abundance. Ecology 76: 2028-2043. [ Links ]
Brown, J.H., G.C. Stevens & D.M. Kaufman. 1996. The geographic range: size, shape, boundaries and
internal structure. Annu. Rev. Ecol. Syst. 27: 597-623.
Campos, J.B. & M.C. Souza. 1997. Vegetação. p. 331-342. In Vazzoler, A.E.A.M., A.A. Agostinho &
N.S. Hahn (eds.). A planície de inundação do alto Rio Paraná - Aspectos físicos, biológicos e
socioeconômicos. EDUEM, Maringá, Paraná. 460 p.
Collins, S.L. & S.M. Gienn. 1991. Importance of spatial and temporal dynamics in species regional
abundance and distribution. Ecology 72: 654-664.
Collins, S.L. & S.M. Glenn. 1997. Effects of organismal and distance scaling on analysis of species
distribution and abundance. Ecol. App. 7: 543-551.
Eriksson, 0. & A. Jakobsson. 1998. Abundance, distribution and life histories of grassland plants: a comparative study of 81 species. J. Ecol. 86: 922-933. [ Links ]
Gaston, K.J. 1990. Patterns in geographic ranges of species. Biol. Rev. 65: 105-129. [ Links ]
Gaston, K.J. 1996. The multiple forms of the interspecific abundance-distribution relationship. Oikos 76:
211-220.
Gaston, K.J. & T.M. Blackbum. 1996. Global scale macroecology: interactions between population size, geographic range size and body size in the Anseriformes. J. An. Ecol. 65: 701-714. [ Links ]
Gaston, K.J., T.M. Blackburn & J.H. Lawton. 1997. Interspecific abundance-range size relationship: an appraisal of mechanisms. J. An. Ecol. 66: 579-601. [ Links ]
Gaston, K.J. & J.H. Lawton. 1990. Effeets oí scale and habitat on the relationship between regional distribution and local abundance. Oikos 58: 329-335. [ Links ]
Gotelli, N.J. & D. Simberloff. 1987. The distribution and abundance of tallgrass prairie plants: a test of
coresatellite hypothesis. Am. Nat. 130: 18-35.
Gotelli, N.J. & W.G. Kelley.1993. A general model of metapopulation dynamics. Oikos 68: 36-44. [ Links ]
Hanski, 1. 1982. Dynamics of regional distribution: the core and satellite species hypothesis. Oikos 38: 210-221.
Hanski, l. 1991. Single-species metapopulation dynamics: concepts, models and observations. Biol. J.
Linn. Soc. 42: 17-38.
Hanski, l., J. Konki & A. Halka. 1993. Three explanations of the positive relationship between distribution
and abundance of species, p. 108-116. In R.E. Ricklefs & D. Schluter (eds.). Species diversity in
ecological communities: Historical and geographical perspectives. The University of Chicago, Chicago,
Illinois.
IBGE - Fundação Instituto Brasileiro de Geografia e Estatística. 1989. Geografia do Brasil - Região
Centro-Oeste. Volume 1. Centro de Documentação e Disseminação de Informações. Rio de Janciro. 267p.
IBGE - Fundação Instituto Brasileiro de Geografia e Estatística. 1992. Manual técnico da vegetação
brasileira. Série Manuais técnicos em geociências n.° 1. Centro de Documentação e Disseminação de
Informações. Rio de Janeiro. 92 p.
Johnson, C.N. 1998. Species extinction and the relationship between distribution and abundance. Nature 394: 272-274. [ Links ]
Levins, R. 1969. Some demographic and genetic consequences of environmental heterogeneity for
biological control. Bull. Ent. Soc. Am. 15: 237-240.
Lorenzi, H. 1992. Árvores brasileiras: Manual de identificação e cultivo de plantas arbóreas nativas do
Brasil. Plantarum, Sáo Paulo. 368 p.
McArthur, R.H. & E.O. Wilson. 1967. The theory of island biogeography. Princeton University, New
Jersey. 203 p.
Mueller-Dombois, D. & H. Ellemberg. 1974. Aims and methods of vegetation ecology. Wiley, New York. 547 p. [ Links ]
Sebeiner, S.M. & J.M. Rey-Benayas. 1997. Placing empirical limits on metapopulation models for
terrestrial plants. Evol. Ecol. 11: 275-288.
SEPLAN-MS - Secretaria de Planejamento. 1989. Macrozoneamento geoambiental do Estado de Mato Grosso do Sul. FINPLAN-MS. Campo Grande, Mato Grosso do Sul. 242 p. [ Links ]
Souza, M.C., J. Cilinski & M.B. Romagnolo. 1997. Levantamento Florístico. p. 343-368. In Vazzoler,
A.E.A.M., A.A. Agostinho & N.S. Hahn (eds.). A planície de inundação do alto Rio Paraná - Aspectos
físicos, biológicos e socioeconómicos. EDUEM, Maringá, Paraná. 460 p.
Tokeshi, M. 1992. Dynamics of distribution in animal communities: Theory and analysis. Res. Popul. Ecol. 34: 249-27. [ Links ]
1 Correspondence to: Setor de Ecologia, Departamento de Biologia; Universidade Federal de Mato Grosso
do Sul; Caixa postal 549; Campo Grande, MS, 79070-900, Brazil. fax 00 5567 787 7555;
sjleite@nin.ufms.hr
2 Setor de Ecologia, Departamento de Biologia, Universidade Federal do Mato Grosso do Sul

