











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
Seaweed is considered an excellent source of bioactive natural products with anti-inflammatory, antioxidant, immunomodulatory, and antitumor effects, among others (Mayer et al., 2019). The advantage of using seaweed is its high availability and low processing costs compared to terrestrial plants (Lee et al., 2022). Recently, the use of seaweed as a source of bioactive principles for the treatment of various cancers, including breast cancer (BC), has been explored. In the United States, BC is the second leading cause of cancer death among women after lung cancer, and the leading cause of cancer death among black and Hispanic women (Giaquinto et al., 2022). Conventional cancer therapy causes negative side effects that debilitate patients. Therefore, there is a need to search for products that complement the classical management of the disease and help mitigate the side effects of treatment (Abudabbus et al., 2017). Proper management of the immune response is important in the fight against cancer. Monitoring the production of pro-inflammatory cytokines is key to the detection of an efficient anti-tumor immune response, whereas the predominance of anti-inflammatory cytokines is an indication that the tumor is gaining ground in the individual and has successfully reversed the immune defense (Lan et al., 2021).
The brown alga Lessonia trabeculata Villouta & Santelices, known as aracanto, palo, palo blanco (Peru) or huiro palo, huiro varilla (Chile), is an endemic species of the eastern Pacific coasts. It is distributed between the meridians 14º (central coast of Peru) and 55º (Chile) south latitude and is the dominant species of the intertidal and shallow subtidal rocky bottom (Santelices et al., 1980). Most studies on this species have focused on ecological and reproductive aspects (Campos et al., 2021; González et al., 2018; Tala et al., 2004).
The use of L. trabeculata is mainly artisanal, and it is mainly used for the extraction of alginates in the industrial sector (Gouraguine et al., 2021). Some reports suggest alternative uses, such as a bioabsorbent for Cd (II) and Hg (III) in environments contaminated by metals (Boschi et al., 2011). Fucoidan is the least studied of the bioactive compounds contained in this bioresource. The structure and composition of fucoidan vary depending on the algal species and abiotic factors of the marine environment (Wang et al., 2020), so it is essential to define the bioactive potential of fucoidan and other compounds in each species.
Regarding its anticancer activity, fucoidan has been shown to exert a cytotoxic effect on tumor cells by modulating apoptosis and the cell cycle (Jin et al., 2021). Fucoidan also stimulates immune functions such as maturation and proliferation of dendritic cells (Park et al., 2020), activation of NK cells (Zhang et al., 2021), macrophages (Ma et al., 2021), cytotoxic T lymphocytes (Kiselevskiy et al., 2022), and modulates cytokine production (Colona, 2022; Takahashi et al., 2018). These properties demonstrate that fucoidan is an enhancer of the immune system’s anti-tumor activity.
Tumor spheroids are three-dimensional in vitro culture models that allow the study of cell-extracellular matrix interactions, hypoxic conditions, drug penetration and tumor physiology. Because they more closely resemble the tumor microenvironment, spheroids are a great advantage for studying the use of fucoidan in cancer treatment (Nii et al., 2020). The objective of our study was to evaluate the antitumor and immunomodulatory activity of L. trabeculata fucoidan (LtF) on two types of multicellular tumor spheroids generated from triple-negative BC cells. We demonstrated that LtF has antitumor and immunomodulatory activity on spheroids. The simple and combined treatments modify the immunosuppressive microenvironment, so with these results we hope to promote the research and development of fucoidan-based products from L. trabeculata as a strategy for cancer prevention and treatment, as well as contribute to the sustainable use of Peru’s natural resources. This is the first study to report the antitumor and immunomodulatory activity of LtF on 4T1 and adds to the existing information on the utility of a 3D model consisting of tumor and immune cells (MTSs) for the evaluation of natural products.
Materials and methods
Chemical reagents and materials: Doxorubicin (Dox, cat. D1515), Roswell Park Memorial Institute medium (RPMI-1640, cat. R8005), 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT, cat. M5655, 0.5 mg/ml) and dimethyl sulfoxide (DMSO, cat. D2438) were purchased from Sigma-Aldrich (Merck KGaA, USA); fetal bovine serum (FBS, cat. S181H) was purchased from Biowest SAS (France); red blood cell lysis buffer (cat. A10492) from Gibco (USA); agarose (cat. 16500500) from Invitrogen (USA); trypan blue (cat. 1450013) from Bio-Rad (United Kingdom), and sandwich ELISA kits for TNF-α (cat. 88-7324-88), IL-6 (cat. 88-7064-88), IL-10 (cat. 88-7105-88) and TGF-β (cat. 88-8350-88) were purchased from Thermo Fisher Scientific Inc. (USA). All chemicals and solvents were of analytical grade.
0.22 μm Stericup-GP Sterile Vacuum Filtration System (cat. S2GPU05RE) was purchased from Millipore (USA). 25 cm2 flasks (cat. 430639), 70 μm cell strainer (cat. 431751), 96-well round-bottom microplates (cat. 3799), and flat-bottom microplate (cat. 3599) were purchased from Corning Inc. (USA). And Immulon 4HBX flat-bottomed microplates (cat. 3855) from Thermo Fisher Scientific Inc. (USA).
Fucoidan: Lyophilized LtF (sugars = 59 %, sulfates = 5.7 %, purity = 83.4 %) was provided by the company PSW SA (https://www.pswsa.com, Lima, Peru), which collected the samples from L. trabeculata in San Nicolas Bay (15°15’39’’ S & 75º13’47’’ W), Marcona District, Nasca Province, Ica Region, Peru.
Preparation of fucoidan and doxorubicin concentrations: A 10 mg/ml solution of LtF was prepared, from which dilutions were made to obtain concentrations of 1, 10, 100, 1 000, 2 000, 4 000, 6 000, 8 000, and 10 000 μg/ml. In the case of Dox, a 10 μg/ml solution was prepared from which dilutions were made to obtain concentrations of 0.01, 0.1, 0.5, 1, 5, and 10 μg/ml. Dox is an antineoplastic drug used to treat BC and was used as a positive control for antitumor and immunomodulating assays. All dilutions were prepared in RPMI-1640 supplemented with 10 % FBS (complete medium: CM), filtered through 0.22 μm membranes, and stored at 4 ºC.
Cell culture of 4T1: 4T1 is a mouse adenocarcinoma cell line (code BCRJ0022), metastatic and triple negative breast cancer (TNBC), was obtained from the Rio de Janeiro Cell Bank (Brazil). Cells were cultured in 25 cm2 flasks in complete medium under standard conditions (37 ºC, 5 % CO2, and 95 % relative humidity) to 80 % confluence. Cultures were maintained in an Esco CelCulture® CO2 incubator (CCL-170B-8, Singapore). Viable cell counts were performed using a Neubauer chamber and trypan blue.
Isolation of mouse splenocytes: Six nulliparous female BALB/c mice, 6 weeks of age, were obtained from the National Institute of Health (Lima, Peru). The use of these animals in the study was reviewed and approved by the Ethics Committee of the Faculty of Veterinary Medicine of the Universidad Nacional Mayor de San Marcos (CEBA code 2020-2). Splenocytes were obtained according to the methodology of Nilofar et al. (2017). Each spleen was fractionated into small pieces in CM, and the resulting cell suspension was passed through a 70 μm cell strainer. Cells were collected and washed with CM at 1 500 rpm for 5 min. The pellet was lysed with 2 ml red blood cell lysis buffer at 4 ºC for 2 min. It was then washed a second time with CM. Finally, the splenocyte pellet was resuspended in 1 ml CM for viable cell counting using a Neubauer chamber and trypan blue.
Preparation of multicellular tumoral spheroids formed with 4T1 (MTS): 96-well round-bottom microplates were coated with 50 μl of 1.5 % agarose per well. Then 200 μl of 4T1 cells (1×104 cells/well) in CM were added. To promote cell aggregation, the microplate was shaken at 60 rpm for 20 min using an incubator-shaker (ZHWY-2102C, Zhicheng, China), and then incubated for 96 h under standard culture conditions. The formation of MTS was verified using a Leica inverted microscope (DM IL LED, Germany).
Preparation of multicellular tumor spheroids composed of 4T1 cells and splenocytes (MTSs): 100 μl of 9×103 4T1 cells were combined with 100 μl of 1×103 splenocytes per well and processed as described for the preparation of MTS. Several preliminary assays were performed to obtain the appropriate cell ratios for the formation of MTS and MTSs.
Antitumor activity on MTS: The cytotoxicity of LtF was determined by a colorimetric assay using MTT. 200 μl of the prepared concentrations of LtF or Dox were added to each MTS and incubated for 72 h under standard conditions. Each concentration was tested in quadruplicate (N = 4). The MTS were then independently transferred to a flat-bottom microplate, 200 μl of MTT reagent dissolved in CM was added and incubated for 4 h under standard conditions. The medium from each well was discarded, 100 μl of dimethyl sulfoxide was added, and incubated for 30 min at 40 rpm on the shaker, at 25 °C. The microplate should be protected from light during the assay. The amount of formazan was quantified at 570 nm with a differential filter of 630 nm, on a spectrophotometer (EPOCH2, Biotek Instruments Inc., USA). Dox-treated MTS were used as positive and untreated MTS as negative controls.
The percentage of viable cells was calculated using the formula: Viability (%) = (average absorbance value of cells treated with the product or drug / average absorbance value of negative control) × 100. Where the product is LtF and the drug is Dox. At the end of the assay, the MTS were observed under an inverted microscope to analyze any changes in their appearance. The degree of cytotoxicity of each LtF or Dox concentration was quantified as the percentage of cell viability using the absorbance values obtained for each assay and the ISO 10993-2009 classification, 100-75 %: Non-cytotoxic, 74-50 %: Mildly cytotoxic, 49-25 %: Moderately cytotoxic, 24-0 %: Extremely cytotoxic.
Determination of the half-maximal inhibitory concentration (IC50): GraphPad Prism software version 8.0.1 (GraphPad Software Inc., San Diego, CA, USA) was used to determine the IC50 for LtF and Dox, using the following formula: Y = 100/1 + 10X-log (IC50) where Y is the cell viability defined between 0 % and 100 %, and X is the logarithm of the concentration of LtF or Dox.
Immunomodulatory activity on MTSs: After 72 h of treatment, under standard conditions, supernatants were collected from: (i) IC50 LtF alone (simple treatment); and (ii) IC50 (LtF+Dox) (combined treatment); positive control (IC50 Dox) and negative control (untreated), centrifuged at 1 500 rpm for 5 min to remove cell debris, and stored at -86 °C.
Quantification of cytokines: Concentrations in the supernatants of MTSs cultures from treatments (i) and (ii) were determined using sandwich ELISA kits for the pro-inflammatory cytokines: Tumor Necrosis Factor-Alpha (TNF-α), Interleukin-6 (IL-6) and the anti-inflammatory cytokines Interleukin-10 (IL-10), Transforming Growth Factor-Beta (TGF-β) (N = 6). 100 μl/well of capture antibody was added to 4HBX flat bottom microplates corresponding to the cytokine of interest diluted in coating buffer. The microplate was sealed and incubated overnight at 4 °C with shaking at 60 rpm according to the manufacturer›s instructions. The appropriate standard curve was generated to determine the cytokine concentrations. Absorbance was measured in a UV/visible spectrophotometer (EPOCH-2, Biotek Instruments Inc., USA) at 450 nm with a differential filter at 570 nm. The assay results were expressed as pg/ml.
Statistical analysis: For the analysis of antitumor activity, the effect of LtF on cell viability was considered, and values are expressed as the mean of replicates ± standard error (SEM). For the analysis of immunomodulatory activity, cytokine production in treatments (i) and (ii) was considered, and values are expressed as the mean of replicates ± standard deviation (SD). In both cases, differences between treatments and controls were analyzed using a one-way analysis of variance (ANOVA), followed by the Tukey’s honestly significant difference (HSD) test. P values: *P < 0.05, **P < 0.01 and ***P < 0.001, compared with the untreated were considered significant, P > 0.05 were not considered significant.
Results
Characteristics of MTS and MTSs spheroids: The spheroids used were of the aggregated type and could represent early-stage tumors given the time spent in their generation and testing (seven days total) with an average diameter of 0.421 mm (Gallardo et al., 2006). At the end of the trials, the untreated MTS and MTSs controls remained compact to the touch, with a dark central zone and a lighter peripheral zone (proliferative zone). However, changes such as cell detachment, increased cell debris, morphologic deformation, and central zone enlargement and darkening were induced by LtF, LtF-Dox or Dox treatments (Fig. 1).
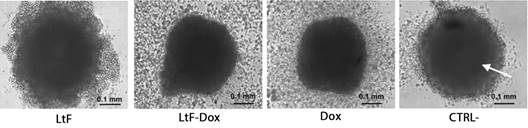
Fig. 1 Morphology of MTSs. Arrow points to central necrotic zone. Abundant cellular debris and peripheral deformation is observed in MTSs treated with LtF, LtF+Dox and Dox. Cell debris was reduced in the CTRL- ×100. IC50 was used for all treatments (N = 4).
Antitumor effect of LtF on MTS: All LtF concentrations showed a significant decrease in cell viability compared to the untreated control (CTRL-) (P < 0.001) (Fig. 2A). Cell viability of MTS treated with 1 and 10 μg/ml. LtF and 10 μg/ml Dox was less than 20 %. LtF may be considered extremely toxic on MTS at concentrations of 1, 10, 4 000, and 6 000 μg/ml. The lowest percentage of viability was observed at the 4 000 μg/ml (5.09 ± 1.52 %). A continuous dose-dependent response observed in the 100 to 6 000 μg/ml range (Fig. 1). The IC50 of LtF was 428 µg/ml.
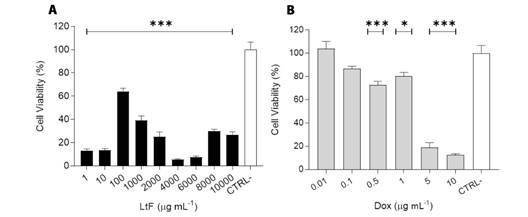
Fig. 2 Antitumor activity of LtF on MTS. A. LtF: 1, 10, 4 000 y 6 000 μg/ml demonstrated extreme cytotoxicity, B. Dox: 5 and 10 μg/ml were extremely cytotoxic. The IC50 value of LtF and Dox was 428 µg/ml and 2 µg/ml respectively. P values: *P < 0.05 and ***P < 0.001, compared with untreated (CTRL-). P < 0.05 was considered significant. Data are expressed as mean ± SEM (N = 4).
Dox-treated spheroids showed a decrease in cell viability of less than 12 % at 10 µg/ml (Fig. 2B). The antitumor activity of Dox was dose dependent and IC50 was 2 µg/ml. At a concentration of 1 µg/ml, LtF showed a greater cytotoxic effect than Dox (P < 0.001), and at 10 µg/ml the effect was similar (P > 0.05); however, the IC50 of Dox was much lower than that of LtF.
Immunomodulatory activity on MTSs: a) Pro-inflammatory cytokines. A significant increase in IL-6 production was observed in the combined and simple treatments compared to the untreated (P ≤ 0.0001) (Fig. 3A). No significant differences were observed between LtF+Dox and Dox (P = 0.55) or between LtF+Dox and LtF (P = 0.12). A significative difference was observed between LtF and Dox (P = 0.0074) (Fig. 3A).
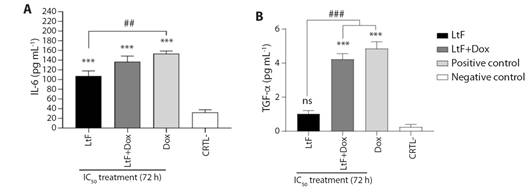
Fig. 3 Pro-inflammatory cytokines produced by MTSs. A. IL-6, B. TNF-α. MTSs were incubated alone or in combination with LtF or LtF+Dox. LtF IC50 = 428 µg/ml, Dox IC50 = 2 µg/ml. Significance obtain by one-way ANOVA is indicated as ***P < 0.001, and Tukey’s post-test significance is indicated as ###P < 0.001, ##P < 0.01; ns, statistically non-significant difference using one-way ANOVA. Data are expressed as mean ± SEM (N = 6).
As for TNF-α, a significant increase in production was observed only in LtF+Dox and Dox treatments compared to LtF and untreated (P = 0.001) (Fig. 3B). TNF-α production was higher with LtF+Dox and Dox than with LtF (P = 0.001, P < 0.001, respectively). There was a similarity between LtF+Dox and Dox (P = 0.44) (4.23 and 4.85 pg/ml, respectively). Although the results for LtF treatment are not significant with respect to LtF+Dox and Dox; however, it is interesting to note that the production of this cytokine was four times higher than in the untreated (1.03 and 0.25 pg/ml, respectively) (P = 0.259). In addition, similar trends were observed for both IL-6 and TNF-α, placing the combined treatment at an intermediate point (Fig. 3A, Fig. 3B).
Immunomodulatory activity on MTSs: b) Anti-inflammatory cytokines. Regarding IL-10, a decrease in its production was found in the simple and combined treatments, and Dox respect to the untreated (P ≤ 0.0001). Dox significantly reduced IL-10 levels compared to LtF+Dox (P = 0.033); IL-10 production was lower with LtF than with LtF+Dox (P = 0.01). This cytokine was similarly inhibited by both LtF and Dox (P = 0.945) (Fig. 4A).
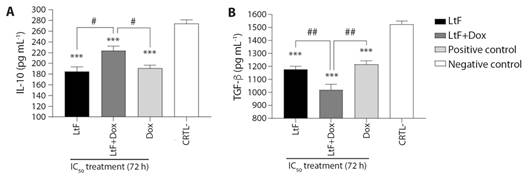
Fig. 4 Anti-inflammatory cytokines produced by MTSs. A. IL-10, B. TGF-β. MTSs were incubated alone or in combination with LtF or LtF+Dox. LtF IC50 = 428 µg/ml, Dox IC50 = 2 µg/ml. Significance obtain by one-way ANOVA is indicated as ***P < 0.001, and Tukey’s post-test significance is #P < 0.05, ##P < 0.01; ns, statistically non-significant difference using one-way ANOVA. Data are expressed as the mean ± SEM (N = 6).
Simple and combined treatments inhibited TGF-β secretion compared to the untreated (P < 0.001), LtF+Dox was highly significant (P ≤ 0.0001) compared to the untreated. No significant differences were found between LtF and Dox (P = 0.804). LtF+Dox affected the production of this cytokine with greater potency than LtF and Dox (P = 0.008 and P = 0.0018, respectively) (Fig. 4B).
It is interesting that in the simple treatment, IL-6 and IL-10 are found in similar concentrations (~100 pg/ml), showing a pattern that could be related to the efforts of immune cells to restore homeostasis and highlighting the immunomodulatory role of LtF.
It is also important to note that in untreated, pro-inflammatory cytokines were produced at minimal levels, approximately 0.25 pg/ml for TNF-α and 32.28 pg/ml for IL-6. It should be noted that 4T1 cells also produce IL-6 and TNF-α (Hsieh & Wang, 2018). In the positive control (Dox), a pro-inflammatory environment was present, whereas in the untreated, high levels of anti-inflammatory cytokines indicate a strong immunosuppressive microenvironment that would promote tumor development in vivo.
Discussion
Breast cancer is the most common malignancy in women, and its incidence is expected to increase by more than 60 % over the next 20 years. Despite advances in detection and treatment, the decline in mortality rates has slowed since 2010, making it a key area of research for the development of new therapeutic alternatives (Lainetti et al., 2020). Standard treatment with chemotherapy and radiotherapy can cause various adverse side effects, such as weakening of the immune system, making the search for new treatments imperative. It is important to note that the success of various treatments depends on a proper assessment that takes into account not only the neoplastic cells, but also the tumor microenvironment (Bożyk et al., 2022). In recent years, the use of three-dimensional (3D) cultures or tumor spheroids has gained popularity because they more accurately simulate in vivo tumor characteristics, including architecture that favors cellular and physiological interactions in the tumor microenvironment. This has reduced the need for laboratory animals and improved the effectiveness of in vitro assays in evaluating new compounds and therapies (Nii et al., 2020; Tevis et al., 2017).
Among the alternatives being explored are products derived from algae, one of whose polysaccharides is fucoidan, which has been shown to have a cytotoxic effect on in vitro cultures of mouse mammary adenocarcinoma 4T1 cells (Atashrazm et al., 2015; Hsu et al., 2013; Xue et al., 2012; Xue et al., 2013) and an antiproliferative effect on the HeLa and U937 cell lines due to selective apoptosis (Colona, 2022).
Comparative studies between 2D and 3D cultures give different results for the same product. This is the case for Fucus evanescens fucoidan (100-800 µg/ml), which has cytotoxic activity in SK-MEL-28 monolayers but loses it when using SK-MEL-28 spheroids (Malyarenko et al., 2021). In the present study, we used MTS spheroids, so it can be said that the results obtained are more like those that can be obtained in vivo. Regarding the effect of the treatments, the MTS or MTSs generally showed a greater number of debris and partial deformation of their appearance, which was more pronounced in those treated with Dox. Baek et al. (2016) observed similar effects on SH-SY5Y (human metastatic neuroblastoma) spheroids, in which the cytotoxic effects of Dox apparently caused degradation of the extracellular matrix, manifested by detachment of all cells from the spheroid in an experiment monitored every 30 min for 5 days. In the present study, the cytotoxic effect was measured after 3 days, possibly the longer the time, the complete deformation of the spheroid would be observed.
Regarding the cytotoxic effect of fucoidan, the reported concentrations for F. vesiculosus on 4T1 monolayers range from 90 to 120 µg/ml (Hsu et al., 2013) and from 50 to 200 µg/ml (Xue et al., 2013). The concentrations used in our study cover a wider range, from 1 to 10 000 µg/ml. It was shown that at a concentration of 1 µg/ml, LtF caused about 90 % cytotoxicity, while at the same concentration Dox caused about 20 %, suggesting that at low concentrations, the fucoidan from this alga would have a greater cytotoxic potency than Dox. At higher concentrations of LtF (8 000 and 10 000 µg/ml), cell viability is affected but not reduced to the low levels of the previous concentrations. This behavior may indicate a saturation of yet unknown receptors on tumor cells through which regulatory processes of the cell cycle and apoptosis are stimulated as cytotoxic (antitumor) mechanisms caused by fucoidan (Colona, 2022; Lin et al., 2020a). The presence of two peaks in MTS viability could be explained by the increased adaptation of tumor cells to these LtF concentrations (Di Nicolantonio et al., 2005). Results can be improved by using spheroids of uniform size, which can be obtained using agarose supports and collagen I matrix (Lin et al., 2020b) or at least two endpoint cell staining methods when performing cytotoxicity assessments (Holst & Oredsson, 2005).
The production of cytokines by immune and tumor cells is related to the stage of the cancer and therefore the treatment administered, the potential outcome depends on the timing of treatment administration. The shift from anti-inflammatory to pro-inflammatory cytokine production creates a favorable environment for tumor progression (Paulsen et al., 2017), which means that a modulating product can be used, considering the tumor stage. Cytokines may serve as markers of tumor stage. In the early stages of cancer, pro-inflammatory cytokines signal the proper functioning of the cellular immune system, the main mechanism for eliminating tumor cells (Berraondo et al., 2019). In a study conducted by Takahashi et al. (2018), fucoidan was administered orally to patients with advanced cancer, which reduced the levels of pro-inflammatory cytokines IL1-β, IL-6, and TNF-α.
In our study IL-6 production was increased after LtF treatment, like that reported for fucoidan from Ascophyllum nodosum, F. evanescens, F. vesiculosus, Sargassum fusiforme, Macrocystis pyrifera, and Undaria pinnatifida in cultures of splenic dendritic cells, bone marrow-derived macrophages, activated splenic macrophages, and human neutrophils (Hsu & Hwang, 2019). Considering that splenocytes (T lymphocytes, B lymphocytes, dendritic cells, macrophages and monocytes) are found in MTSs, it is expected that these cells will make direct contact with 4T1 tumor cells, become activated and produce IL-6; however, IL-6 levels were lower in the untreated than in the LtF, indicating that the antigenic stimulus of 4T1 was not sufficient and that there was a stimulatory effect caused by LtF. This is a multifunctional cytokine whose relationship to inflammation and BC is not easy to establish because it has pro-inflammatory and anti-inflammatory functions that depend on the signaling pathway. Elevated levels of IL-6 secretion may accelerate tumor cell growth by suppressing apoptosis and promoting angiogenesis; however, the results are controversial. In a study of patients with early-stage invasive BC, high IL-6 expression was associated with improved disease-free survival and specific survival (Chen J., et al., 2022).
TNF-α controls immune and inflammatory responses during the early stage of tumor development (Lee et al., 2022). In the combined treatment, there was a decrease with respect to the Dox treatment, which could be related to the modulation exerted by fucoidan, since in the simple treatment the production of this cytokine is not significant, although it is four times higher than in the untreated. To clarify the modulatory role of LtF in reducing TNF-α production, it would be necessary to perform assays taking into account the IC50 of Dox and different concentrations of LtF to determine the combination index (Yunita et al., 2020). Also, assays on the production kinetics of this cytokine should be included, considering times shorter than the 72 h used in the present study. In Dox-treated MTSs cultures, there was a significant increase in TNF-α levels as reported by Syukri et al. (2022), which was confirmed in the present study. This cytokine favors the activation, differentiation, survival or death of cancer cells under certain conditions, an excess could exacerbate the inflammatory process and accelerate tumor development, so reducing its production in advanced stages of cancer would be beneficial. Although the differences are not significant, for both IL-6 and TNF-α there is evidence of a trend towards a reduction of this cytokine with LtF+Dox. Under highly inflammatory conditions, S. hemiphyllum fucoidan reduced IL-6 and TNF-α levels (Chen B., et al., 2022).
Overexpression of IL-10 is a poor prognostic indicator associated with drug resistance, metastatic cancer, and a high probability of recurrence (Li et al., 2014). The decrease of this cytokine observed in supernatants of MTSs as a result of the treatments applied is significant compared to the untreated. A better decrease was observed with LtF and Dox than with LtF+Dox, highlighting the immunomodulatory capacity of LtF in the system used. Increased production of this cytokine in LtF+Dox may be beneficial in the treatment of advanced stage BC, as it would attenuate the Dox-induced inflammatory state.
Increased TGF-β production is associated with poor survival in BC because it suppresses the anti-tumor immune response and promotes metastasis through increased levels of angiogenic and connective tissue growth proteins that promote epithelial-to-mesenchymal transition, particularly in BC. It also stimulates several signaling networks involved in cell growth, and differentiation (Lee et al., 2022). In our study, the modulation exerted by LtF on Dox was demonstrated by the remarkable decrease of TGF-β in the combined treatment, as Dox has been shown to stimulate TGF-β signaling and consequently increase metastasis in BC cells (Yunita et al., 2020). Likewise, TGF-β levels decreased in all three treatments compared to untreated. Similar results were obtained by Xue et al. (2017) using fucoidan from F. evanescens in a drug-induced rat mammary carcinoma model. Chen L., et al. (2022) using fucoidan combined with olaparib from Laminaria japonica in human monocytes, and Guo et al. (2022) in mice with 4T1 adenocarcinoma, successfully using micelles formed by fucoidan and Dox to remodel the immunosuppressive microenvironment and prevent metastasis.
In our study, the increase in IL-6 production in the simple treatment and the decrease in TGF-β levels in the combined treatment indicate that LtF has a modulatory effect on the tumor microenvironment. Due to the elevated TGF-β concentration produced by untreated MTSs, it is confirmed that this is a poor prognostic marker for BC, even in early stage, these results could be compared with serum samples from individuals with early BC.
According to the literature review, the present study is the first to demonstrate the antitumor and immunomodulatory activity of LtF from the Peruvian Sea on BC tumor spheroids, but assays with more complex spheroids are needed to consolidate these results. In conclusion, the immunomodulatory activity of LtF, the single and combined treatments induced a significant production of IL-6. In the case of TNF-α, a study of the kinetics of its production in the model tested is required, since this cytokine is the first to be secreted, it is suggested to use shorter times than the one used. The presence of anti-inflammatory cytokines in the untreated controls confirms the strong immunosuppression generated in the tumor microenvironment. LtF is a good candidate for the treatment of BC with the ability to immunomodulate the tumor microenvironment alone or in combination with Dox, which should be further investigated.
Ethical statement: the authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgement section. A signed document has been filed in the journal archives.

