
Análisis dialélico del rendimiento de Chile por el Método IV de griffing
Mirna Hernández-Pérez2, Alfonso López-Benítez2, Fernando-Borrego Escalante2, Sandra Roxana López-Betancourt2, Moisés Ramírez-Meraz3
1 Parte de una tesis de doctorado en el departamento de fitomejoramiento de la Universidad Autónoma Agraria Antonio Narro.
2 Departamento de Fitomejoramiento, Universidad Autónoma Agraria Antonio Narro. Dir. Hacienda de Enmedio, manzana 14, lote 32, casa B. Toluca, Edo. de México. C.P. 50210. hepemi@hotmail.com; alobe42@hotmail.com. fborregoe@hotmail.com
3 Campo Experimental Las Huastecas (CEHUAS). Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Villa Cuauhtémoc, Tamaulipas. Correspondencia alobe42@hotmail.com
Dirección para correspondencia
Resumen
]]> Análisis dialélico del rendimiento de chile por el método IV de Griffing. El objetivo de este trabajo fue estimar la aptitud combinatoria general de genotipos de chile y la aptitud combinatoria específica de sus cruzas directas. Se utilizó el método IV modelo fijo de Griffing, con la finalidad de definir el esquema de mejoramiento genético. Los cruzamientos se realizaron en invernadero en la Universidad Autónoma Agraria Antonio Narro (UAAN), en Saltillo, Coahuila. La evaluación se realizó en el Campo Experimental Huastecas, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Villa Cuauhtémoc, Tamaulipas, en el ciclo otoño-invierno de 2008. Se utilizaron seis tipos de chile tres tipo jalapeño (4, 6 y 118), tipo húngaro amarillo (8), uno tipo ancho (16) y uno tipo mulato. Se evaluó la progenie de seis cruzas directas, para días a floración y días a corte, y rendimiento de frutos por plantas, diámetro, longitud y peso promedio del fruto y rendimiento por hectárea. Los mejores genotipos fueron 6 y 16, por su mayor aptitud combinatoria general, y las mejores cruzas por su aptitud combinatoria específica fueron 6x16 y 4x6, la cruza 16x118 mostró alto rendimiento.Palabras clave: Efectos genéticos, Capsicum annuum, aptitud combinatoria general, aptitud combinatoria específica, análisis genético, cruzamiento dialélico.
Abstract
Diallel analysis of yield in hot pepper by the method IV of Griffing. The objective of this study was to estimate the General and Specific Combining Ability (GCA and SCA) values of yield in pepper. The IV fixed model of Griffing was used to define the best breeding method. Crosses were conducted in a greenhouse at the Universidad Autónoma Agraria Antonio Narro (UAAAN ), Saltillo, Coahuila. The evaluation was realized in the Experimental Station of Huastecas, National Institute of Research Forestry Agricultural and Livestock (INIFAP), Villa Cuauhtémoc, Tamaulipas, in the autumn-winter cycle of 2008. Six types of peper were utilized, three belonging to jalapeño type (4, 6 y 118), one Yellow wax type (8), one broad ("ancho") type (16) and a brown ("mulato") type. Six direct crosses were evaluated for days to flowering and days to first harvest, fruit yield per plant, fruit diameter, and fruit length, and average weight per fruit and yield per hectare. The best genotypes were 6 and 16, and the best crosses for their specific combining ability were 6x16, 4x6. Cross 16x118 showed higher yield.
Key words: Genetic effects, Capsicum annuum, general combining ability, specific combining ability, genetic analysis, diallel cross.
Introducción
Un diseño dialélico, según Layrisse (1981), está constituido por todos los cruzamientos posibles entre un conjunto de padres, pero frecuentemente no incluye a los padres o a las cruzas recíprocas. Las conclusiones de un análisis dialélico, de acuerdo al autor, dependen del modelo genético-estadístico, de las suposiciones planteadas y del método de selección de los padres. El análisis dialélico ha sido usado para estimar heterosis en especies cultivadas, para evaluar el comportamiento de líneas en combinaciones híbridas, para seleccionar ciertos padres o cruzas como fuente de germoplasma mejorado (Murray et al. 2003).
Por otro lado, entre los métodos existentes para evaluar las cualidades de un conjunto de progenitores se encuentran los diseños dialélicos propuestos por Griffing (1956), que permiten identificar las mejores combinaciones. Sprague y Tatum (1942) propusieron el método que incluye a las cruzas dialélicas y que originó los conceptos de aptitud combinatoria general y específica.
El término de Aptitud Combinatoria (AC) se refiere a la capacidad que tiene un individuo o una población de combinarse con otros, dicha capacidad medida por la progenie (Márquez 1988). Estudios de aptitud combinatoria, tienen suma importancia en cuanto a la identificación de los progenitores potenciales que pueden ser útiles para producción de híbridos o para el desarrollo de poblaciones compuestas o sintéticas (Martínez 1983). Sin embargo, la aptitud combinatoria debe determinarse no sólo en un individuo de la población sino en varios; a fin de poder seleccionar los que exhiban la más alta aptitud combinatoria. Al respecto Hoegenmeyer y Hallauer (1976) señalaron que la aptitud combinatoria específica (ACE) es más importante que la aptitud combinatoria general (ACG) en un programa de mejoramiento cuya finalidad sea la obtención de híbridos, ya que con la ACE se puede hacer mejor uso de los efectos no aditivos como la dominancia y la epistásis. Además, la ACG explica la proporción de la varianza genotípica debida a los efectos aditivos de los genes, mientras que la ACE revela la proporción de éste que puede deberse a las desviaciones de dominancia. Los efectos de ACE fueron más importantes que los de ACG cuando los materia les fueron sometidos a selección (Si ngh y Chaudary 1985).
Si las variedades o líneas representan un grupo fijo, las estimaciones de las constantes genéticas suministran información acerca de un grupo particular de padres y sus cruzas, mientras que las estimaciones de los componentes de varianza podrían tener poco valor porque no hay una población base a la cual podría aplicarse (Gardner-Eberhart 1966). Dichos autores mencionan que el método dos en el modelo uno del análisis de Griffing (1956) está diseñado para el caso de un grupo fijo de padres y su cruce dialélico, de tal manera que el análisis de varianza es el mismo que el análisis dos de su modelo.
]]> Los componentes aditivos y no aditivos de la varianza genotípica paterna son estimados al usar los componentes de varianza de las capacidades combinatorias general y específica (Griffing 1956). Este autor desarrolló dos modelos diferentes de análisis dependiendo de los diferentes supuestos de muestreo: Cuando se asume sólo las líneas paternas o el material experimental como un todo, son una muestra aleatoria de alguna población sobre la cual se van a hacer inferencias y cuando se seleccionan las líneas deliberadamente y no pueden ser consideradas como una muestra al azar de alguna población, el material experimental constituye la población entera sobre la cual se van a hacer inferencias válidas. El objetivo de este trabajo fue estimar la aptitud combinatoria general de genotipos de chile y la aptitud combinatoria específica de sus cruzas directas.Materiales y métodos
El presente estudio se realizó con seis materiales genéticos (Cuadro 1) de diferentes tipos de chile con diferentes orígenes, tres líneas tipo jalapeño derivados de colectas realizadas por la Universidad Autónoma Agraria Antonio Narro (4, 6 y 130), uno tipo Húngaro amarillo con el mismo origen (8), uno tipo ancho procedente de la compañía de semillas Berensen (16), uno tipo, jalapeño procedente de Río Bravo Tamaulipas y uno tipo mulato (130) procedente del Centro de Investigaciones Agrícolas del Bajío (CIAB) del Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP). Estos materiales fueron elegidos para hacer un cruzamiento dialélico por haber presentado diferentes reacciones a la infección por Phytophthora capsici en evaluaciones previas pero que no son consideradas en este estudio.
Las cruzas se realizaron en las instalaciones de la Universidad Autónoma Agraria "Antonio Narro", ubicada al sur de la Ciudad de Saltillo, Coahuila, México, localizada a 25º 22’ latitud N; 101º 00’ longitud W y con una altitud de 1742 msnm, con una temperatura media anual de 16,8ºC. El modelo genético estadístico estuvo basado en una muestra fija, sin considerar las cruzas recíprocas. La evaluación de campo se hizo en el Instituto Nacional de Investigaciones Agrícolas y Pecuarias (INIFAP) de Tampico, Tamaulipas, en el Campo Experimental Las Huastecas (CEHUAS), localizado el km 55 de la carretera Tampico-Mante, con las coordenadas: 23 latitud 22°34´ Norte y longitud 98°05´ Oeste, y altitud de 60 msnm, en la Huasteca al Nororiente de la República Mexicana. Tanto los seis progenitores como sus quince cruzas simples F1 resultantes del cruzamiento dialélico se establecieron en el ciclo primavera-verano del 2008 bajo un diseño experimental de bloques completos al azar con cuatro repeticiones. Cada uno se estableció con dos surcos de 2,5 m de longitud, una separación entre estos de 0,9 m y 0,3 m entre plantas, para una densidad de 32 000 plantas/hectárea.
]]> El manejo del experimento en general, fue de acuerdo a las recomendaciones del mismo campo experimental. Las variables evaluadas fueron días a floración (DAF), días al primer corte de frutos (DAC), número de frutos promedio por planta (FTO/PTA), longitud de fruto (LFTO), diámetro de fruto (DFTO)), peso promedio de frutos (PPF) y rendimiento (t/ha). Para días a floración se consideró el número de días transcurridos desde el día del transplante hasta que el 50 % de las plantas estuvieran en floración. Para días al primer corte se contó el número de días transcurridos desde la fecha del transplante hasta el día en que se realizó la primera cosecha. Para el número promedio por planta, se cortaron todos los frutos de todas las plantas de cada genotipo. La longitud promedio, se determinó en cinco frutos tomados al azar de cada genotipo y se midió su longitud. El diámetro promedio se determinó en los mismos frutos considerados para la longitud promedio. Para el peso promedio de frutos por planta se pesaron los frutos cosechados. El rendimiento en toneladas por hectárea se calculó determinando el rendimiento obtenido en ocho plantas de cada genotipo en una superficie de 2,25 m2 y se estimó el rendimiento por hectárea.Análisis genético
Se empleó el método IV de Griffing (1956) para el análisis del cruzamiento dialélico, en el cual se estudian solo las cruzas directas, teniendo quince por evaluar. Se estimó la habilidad combinatoria de las líneas y se comparó el comportamiento de las mismas en combinaciones híbridas, con el Programa Diallel (Burow y Coors 1994), se estimó ACG y ACE para cada una de las características evaluadas. El análisis se realizó bajo el modelo genético estadístico siguiente:
Yijk = μ+ Gi + Sij + ijk
Donde:
Yijk = Observación ijk-ésima.
μ = media de la poblaciσn.
Gi = efecto de la CCG del i-ésimo y j-ésimo padre o línea.
Sij = efecto de la CCE del cruce entre el i-ésimo y el j-ésimo padre o línea.
ijk = error experimental.
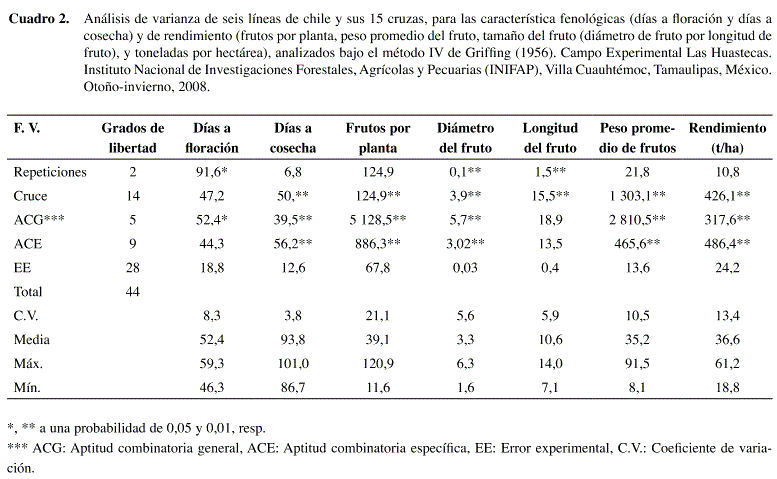
El análisis de varianza (ANOVA) para los siete parámetros evaluados, cuyos cuadrados medios, coeficientes de variación, así como su respectiva media y rango se muestra en el Cuadro 2.
Para la variable de respuesta días a floración, no se encontró significancia en las fuentes de variación ACE ni en la de cruzas; sin embargo, se detectaron diferencias (P≤0,05) en ACG y en repeticiones. En esta variable las cruzas tuvieron un comportamiento muy similar.
Para la variable días a cosecha, la cruza resultó significativa (P≤0,01), así como también para la ACG y la ACE.
Para la variable número de frutos por planta (FTO/PTA), los valores para ACG en progenitores y para ACE en las cruzas también resultaron con diferencias significativas (P≤0,01), lo anterior indica el gran potencial de producción que presentan estos materiales.
En diámetro de fruto (DFTO), las repeticiones, cruzas, ACG y ACE, mostraron diferencias significativas (P≤0,01), y en longitud de fruto (LFTO), hubo diferencias significativas (P≤0,01), tanto, repeticiones como cruzas.
El peso promedio por fruto (PPF), las cruzas, ACG y ACE se comportaron de manera similar al anterior, siendo estas significativas (P≤0,01). Para toneladas por hectárea, la cruza resultó ser significativa (P≤0,01), así como para ACG y ACG, que también fueron significativas (P≤0,01).
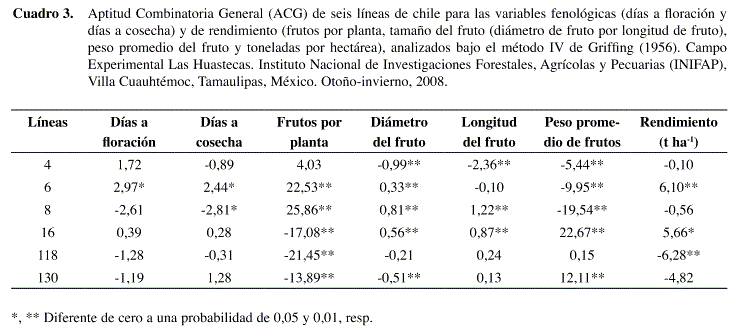
Para la variable FTO/PTA el genotipo 8 obtuvo el valor más alto de ACG seguida del genotipo 6; respecto al valor más bajo observado en el genotipo 118 (Cuadro 3).
De acuerdo con los datos observados, la variable PPF el genotipo 8 sería la que presentara el mayor valor mayor de ACG (Cuadro 3), sin embargo se observa que es la que presentó el valor más bajo (-19,54) lo que nos indica que en realidad se produjeron muchos frutos por planta pero de muy bajo peso, siendo el genotipo 16 el que presentó el valor más alto de ACG en esta variable, por lo anterior podemos decir que este comportamiento es inverso, donde se produjeron pocos frutos por planta pero pesados.
Las variables diámetro y longitud de fruto, nos dan su tamaño real (Cuadro 3), el genotipo 8 mostró el valor más alto de ACG y el de menor valor fue el genotipo 4. Aquí podríamos pensar que el genotipo 16 es el que debería ser más grande, debido a que se trata de un ancho, pero no fue así, aunque este genotipo también fue altamente significativo.
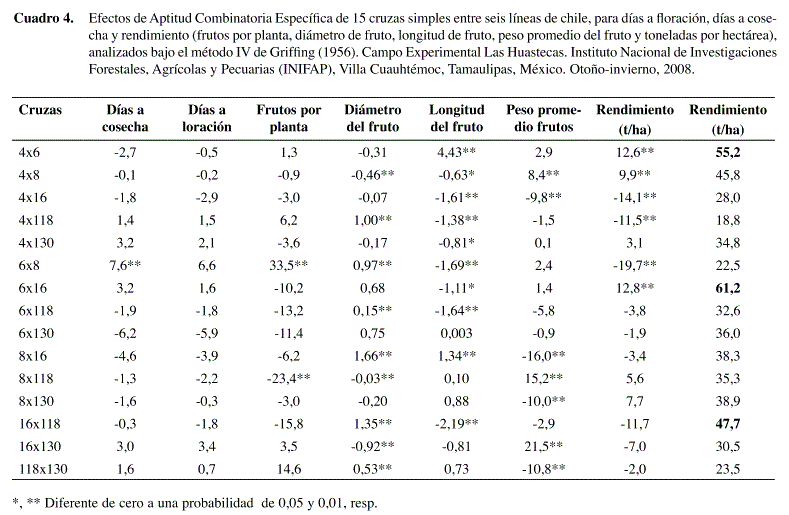
]]> Para la variable rendimiento, el genotipo 6 fue el mejor, seguido del 16. Debido al mayor número de días a floración (DAF) y mayores días a cosecha (DAC) y con mayor número de cosechas (Cuadro 3), este genotipo obtuvo una diferencia altamente significativa para la variable FTO/PTA, lo cual nos indica que la planta obtuvo un buen número de frutos y un peso promedio por fruto (PPF) promedio (no muy pesados), esto nos refleja el mayor número de toneladas por hectárea. Para el genotipo 16 se puede decir que tuvo pocos frutos y de alto peso (alta ACG) y por lo tanto obtuvo mayores rendimientos. De acuerdo con Dorantes et al. (2004), al trabajar con siete líneas progenitoras, de chile jalapeño, se encontró que el rendimiento de fruto es un carácter determinado tanto por efectos aditivos como de dominancia (50%), longitud y diámetro de fruto están determinados solamente por efectos de dominancia.En los efectos de ACE de las quince cruzas simples (Cuadro 4), la cruza 6x8 resultó ser significativa para DAC , FTO/PTA, DFTO, LFTO, pero tuvo un bajo rendimiento (22,5 t/ha), a pesar de que el progenitor 6 tuvo ACG significativa en t/ha, pero el progenitor 8 sólo tuvo una alta ACG en frutos por planta y en t ha-1 la ACG resultó ser negativa (-19,7 t/ha).
La mejor cruza fue la 6X16, ya que obtuvo una ACE significativa y es la que tuvo un mayor número de toneladas por hectárea (61,2 t/ha). El genotipo 6 obtuvo la mayor ACG en t/ha y el genotipo 16 tuvo una ACG significativa en PPF, que al ser cruzados dio un efecto aditivo para t/ha, no así para PPF (Cuadro 4).
También se observa que la cruza 4X6 fue altamente significativa para ACE y la segunda en rendimiento con 55,2 t/ha, donde el progenitor 4 tuvo una ACG significativa pero negativa y el progenitor 6, tuvo la mayor ACG , y al ser cruzados se obtuvo una alta ACE (Cuadro 4).
La cruza 16x118 mostró un alto rendimiento (47 t/ha), donde los progenitores, el genotipo 16 como ya se mencionó fue el más alto ACG para PPF y el progenitor 118 tuvo significancia negativa tanto en frutos por planta, como para toneladas por hectárea.
Pérez et al. (2009) al utilizar seis variedades criollas (cinco colectadas en México y una en Perú) de chile manzano (Capsicum pubescences R y P) y todas sus posibles cruzas directas, encontró que tanto para ACG, como para ACE mostraron efectos significativos en el rendimie nto de fruto, volumen, grosor de pericarpio, peso y número de semillas y número de lóculos por fruto. El valor más alto se registró en la variedad "Puebla", ya que generó el mayor número de híbridos de alto rendimiento, volumen de fruto y grosor de pericarpio, en comparación con los otros cinco progenitores.
Una cruza simple será de alto rendimiento si sus dos líneas o genotipos progenitores son de alta ACG o si su efecto de ACE es alto y al menos uno de sus progenitores es de alta ACG. El máximo rendimiento de una cruza ocurre cuando sus dos progenitores son de alta ACG y su efecto de ACE es alto.
Los mejores genotipos obtenidos fueron la 6 y la 16 y las mejores cruzas obtenidas fueron 6x16, 4x6 y 16x118.
El genotipo seis fue considerado el mejor porque obtuvo la mayor ACG en rendimiento, debido a que este genotipo haya sido el mejor en rendimiento se debe a que a mayores DAF habrá mayores DAC y por lo tanto un mayor número de cortes, así obtuvo una diferencia altamente significativa para la variable frutos por planta, lo cual nos indica además que obtuvo un PPF promedio (no muy pesados), y el mayor número de toneladas por hectárea.
La línea 16 tuvo una diferencia altamente significativa de ACG en PPF y valores altos de ACG en rendimiento, tuvo pocos frutos pero pesados y obtuvo los mayores rendimientos.
]]> La mejor cruza fue la 6x16, que obtuvo una ACE altamente significativa y un mayor rendimiento (61,1667 t/ha), lo anterior se debe a sus progenitores; el genotipo 6 obtuvo la mayor ACG en t/ha y el genotipo 16 tuvo una ACG altamente significativa en PPF, que al ser cruzados nos dio un efecto aditivo para t/ha, no así para PPF.La cruza 4x6, quien también es considerada como una de las mejores, tuvo ACE altamente significativa para rendimie nto y es la segunda en producción (55,2333 t/ha). El progenitor 4 tuvo una ACG altamente significativa pero negativa y el progenitor 6, tuvo la mayor ACG, que al ser cruzados mostraron una alta ACE debido a efectos de aditividad.
La cruza 16x118 tuvo un alto rendimiento (47 t/ha), aunque en ninguna variable fue significativa para ACE. Entre sus progenitores, el genotipo 16 tuvo la más alta ACG para PPF y la línea 118 tuvo una significancia alta pero negativa tanto en ftos/pta, como para rendimiento, lo anterior se puede deducir por efectos de dominancia.
Literatura citada
Burow, MD; Coors, JG. 1994. DIALLEL: A microcomputer program for the simulation and analysis of diallel crosses. Crop Sci. 86:154-158. [ Links ]
Dorantes, GJRA; Martínez, ZG; Ramírez, MM; Rosa de la, LA; Campodónico, PO. 2004. Efectos genéticos y heterosis de caracteres de calidad comercial en chile serrano. Primera Convención Mundial del Chile. p. 21-28. [ Links ]
Gardner CO; Eberhart, SA. 1966. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 22:439-452. [ Links ]
]]>Griffing, BJ. 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. Biol. Sci. 9:463-493. [ Links ]
Hoegenmeyer, TC; Hallauer, AR. 1976. Selection among and within full-sib families to develop single crosses of maize. Crop Sci.16:76-80. [ Links ]
Layrisse. 1981. Análisis de un cruce dia lélico de 10 líneas de maní Arachis hypogaea provenie ntes de cinco centros suramericanos de diversidad, en la generación F2. Universidad Central de Venezuela. Facultad de Agronomía. Trabajo de ascenso. Mimeografiado (Ven). 917 p. [ Links ]
Márquez, SF. 1988. Genotecnia vegetal. Tomo II. Editor AGT. México. 563 p. [ Links ]
Martínez, GA. 1983. Diseños y análisis de experimentos de cruzas dialélicas. Colegio de Posgraduados, México. 251 p. [ Links ]
]]>Murray, LW; Ray, IM; Segovia, LA. 2003. Clarification and reevaluation of population-based diallel analysis: Gardner and Ebrhart analysis II and II revisited. Crop Sci. 43:1930-1937. [ Links ]
Pérez, GM; González, HVA; Peña, LA; Sahagún, CJ. 2009. Combining ability and heterosis for fruit yield and quality in manzano hot pepper (Capsicum pubescens r & p) landraces. Universidad Autónoma Chapingo. Revista chapingo serie horticultura 15(1): 47-55. [ Links ]
Singh RK; Chaudary, BD. 1985. Biometrical methods in quantitative genetic analysis. Kalyani Publishers. New Delhi, India. 319 p. [ Links ]
Sprague GF; Tatum LA. 1942. General vs specific combining ability in single crosses of corn. J. Am. Soc. Agron. 34:923-932. [ Links ]
Recibido: 27 de octubre, 2009. Aceptado: 16 de mayo, 2011.
]]>{kind=link}
{kind=link}
{kind=link}