La semilla de Campsiandra angustifolia (Fabaceae:Caesalpiniodeae)como un reflejo de las presiones selectivas sobre su dispersión y establecimiento
Alejandro G. Farji-Brener 1 ,Sandra Durán 2 ,Alejandro Valerio 3 ,Estela Herbas 4 , Mario Castañeda 5, José Ochoa 6 & Mónica Romo 7
1 Lab. Ecotono, Depto de Ecología, CRUB-UNC, Unidad Postal Universidad del Comahue, 8400 Bariloche,Argentina; alefarji@crub.uncoma.edu.ar;alefarji@yahoo.com.*Corresponding author.
2 Facultad de Ciencias, Universidad del Valle, Cali, Colombia;aphe@hotmail.com
3 Dept.of Entomology,University of Arkansas,321 Agri Building Arkansas 72701, USA; avaleri@uark.edu
4 Cerro Alto de la Alianza 192,Cochabamba 3592,Bolivia;ehbaeny@yahoo.com
5 Universidad Autónoma de México,México DF,México;marioc@ibiologia.unam.mx
6 Urb.Kennedy A,Los Brillantes A-19,Cusco,Perú;jaochoac@hotmail.com
7 Cl.Chinchon 858-A,Lima 27,Lima,Perú;romomonica@hotmail.com ]]>
Recibido 05-V-2003. Corregido 22-IV-2004. Aceptado 28-V-2004.
Abstract
The seeds of Campsiandra angustifolia (Fabaceae:Caesalpiniodeae) as a reflex of selective pressures on dispersal and establishment. We indirectly evaluated the selective pressures on dispersal and establishment of Campsiandra angustifolia, a common water-dispersed tree from the Peruvian Amazon, analyzing the variation in the relationship between the volume occupied by dispersal and establishment structures in a total of 535 seeds from 13 trees located at three different habitats. The seeds differed one order of magnitude in their total volume. However, independently of their size and the location of the maternal tree, the relationship between the volume occupied by dispersal and establishment structures was relatively constant (~1) and showed a normal distribution with low skewness, indicating stabilizing selection. These results suggest that, in the habitats studied, dispersal and establishment processes may have similar importance to C.angustifolia. In species with seeds confined in pods, and therefore strongly space-limited, the relative volume of their seeds occupied by dispersal and establishment structures could be a better measure of the trade-off between these two processes than the variation in seed size. Rev. Biol. Trop. 53(1-2):63-71. Epub 2005 Jun 24.
Key words: Amazonian, dispersal, establishment, hidrochory, seasonally flooded forest.
En general, los organismos acceden a una cantidad limitada de recursos que deben asignar a numerosos procesos; en consecuencia, la selección natural beneficiará al organismo que realice una mayor inversión en el proceso más limitante para su supervivencia y reproducción (Futuyma 1986). Sin embargo, la importancia relativa de los procesos -y por ende los beneficios netos de realizar inversiones en ellos- puede variar espacialmente en función de las características del hábitat (Thompson 1999). Por ejemplo, para las plantas el beneficio neto de asignar recursos a su crecimiento, defensa o reproducción dependerá de la importancia relativa de las perturbaciones, los herbívoros y la disponibilidad de recursos, factores que varían de un hábitat a otro (Grime 1979, Coley et al .1985, Herms y Mattson 1992).
Las variaciones en la forma, tamaño o volumen de ciertas estructuras en las plantas pueden interpretarse como el reflejo de las presiones de selección sobre las funciones que cumplen (Futuyma 1986). Por lo tanto, una forma indirecta de estimar la importancia relativa de ciertos procesos ecológicos es analizar, entre los organismos de una población, las variaciones de las estructuras relacionadas con dichos procesos.
La dispersión y el establecimiento son procesos críticos para la supervivencia de las plantas (Harper 1977), pero su importancia relativa puede variar de acuerdo a las condiciones del ambiente. Por ejemplo, si la planta madre atrae depredadores de semillas o si hay baja abundancia de dispersores, los individuos que asignen una mayor inversión relativa a estructuras que maximicen la dispersión de sus semillas serán positivamente seleccionados (Augspurger 1983). Por otra parte, si la dispersión no es limitante pero las áreas potenciales de colonización poseen suelos pobres o bajos niveles de luz, es posible que los individuos con una mayor inversión relativa en estructuras de establecimiento sean positivamente seleccionados (Howe y Smallwood 1982, Dalling et al.1998).
En plantas con dispersión abiótica, la importancia relativa de la dispersión y el establecimiento pueden expresarse en una misma estructura (i.e., cuando la diáspora es la semilla), constituyendo fuerzas selectivas de sentido opuesto y permitiendo así el estudio indirecto de dichos procesos. Por ejemplo, si el establecimiento es un factor limitante, las semillas grandes se verán beneficiadas ya que tienen mas reservas para que la plántula sobreviva en ambientes de baja luminosidad y poca disponibilidad de nutrientes (Walters y Reich 2000). Por otra parte, si existe una mayor presión selectiva para optimizar la dispersión, en general las semillas pequeñas se verán beneficiadas debido a que son más efectivas en recorrer mayores distancias (Augspuger y Hogan 1983, Greene y Johnson 1993). Consecuentemente, el tamaño de la semilla es considerado como una solución de compromiso entre los procesos de establecimiento y dispersión (Ganeshaiah y Uma Shaanker 1991, Greener y Johnson 1993). Sin embargo, el tamaño de la semilla no es el único reflejo del compromiso entre estos dos procesos. En muchas especies dispersadas por el viento o el agua, la semilla, aparte de albergar estructuras para su establecimiento (e.g., cotiledones), posee modificaciones del tegumento adaptadas para su dispersión como alas o tejido flotante (Augspurger 1986, López 2001). La variación relativa del volumen que en la semilla ocupan las estructuras para su dispersión y establecimiento, independientemente de su tamaño, también puede interpretarse como un producto de la selección natural y por lo tanto ayudarnos a evaluar la importancia relativa de estos procesos. Esta variación se torna especialmente importante en semillas cuyo tamaño final se encuentra limitado (e.g., dentro de vainas), ya que el volumen ocupado por estructuras relacionadas con un proceso implica un detrimento en el volumen ocupado por estructuras relacionadas con el otro. Consecuentemente, la distribución en la semilla del espacio ocupado por estructuras relacionadas con la dispersión y el establecimiento debería estar sujeta a una gran presión de selección, y por ende, variaciones en esta distribución relativa deberían reflejar variaciones en la importancia relativa de estos procesos.
]]> El objetivo de este trabajo fue estimar indirectamente la importancia relativa de la dispersión y el establecimiento en Campsiandra angustifolia, un árbol común de la Amazonía peruana que presenta dispersión hidrocórica (Vásquez 1997), analizando la variación del volumen ocupado por estructuras de dispersión y establecimiento en sus semillas. En particular, se analizó la variación de esta relación entre semillas de diferente tamaño y entre árboles localizados en diferentes hábitats.Material y métodos
Este estudio se llevó a cabo durante el mes de mayo del 2001 en bosques inundables cercanos al Río Sucusari, provincia de Maynas, departamento de Loreto, Amazonía Peruana (3º 17 59 S, 72º 54 25 O).La temperatura promedio anual es de 29.5º C y la precipitación promedio de 2 950 mm (Vásquez 1997). Para obtener semillas de árboles potencialmente sujetos a diferentes presiones ambientales, se trabajó en tres tipos de ambiente que representan los diferentes hábitats existentes en el área de estudio (Vásquez 1997). Se seleccionó un total de 13 árboles adultos de C.angustifolia, cuatro ubicados sobre el margen del Río Sucusari, cuatro sobre las márgenes de un pequeño riachuelo a 3 km río arriba de su desembocadura (caño Shimigay), y cinco sobre la costa de una laguna fluvial de aguas negras (laguna Lorenzo). Se consideró que estos hábitats presentaban diferencias en factores que potencialmente limitan la dispersión o el establecimiento de C.angustifolia (i.e., la calidad nutritiva del suelo, la velocidad de la corriente del agua, el tiempo de inundación, la composición y abundancia de los depredadores de semillas, etc.). Es importante destacar que no se tenía ninguna hipótesis a priori sobre cual de estos hábitats favorecía o desfavorecía la importancia relativa de la dispersión o el establecimiento en C.angustifolia.
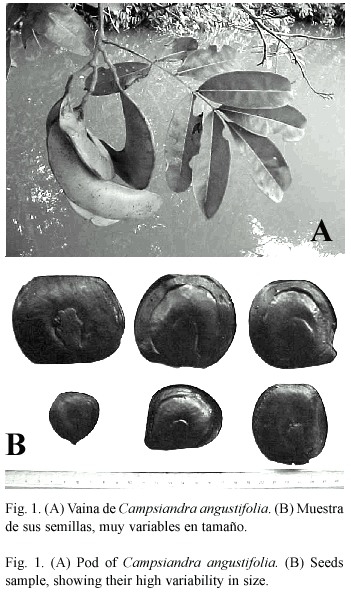
De cada árbol se seleccionaron ramas al azar y se recolectaron alrededor de 50 semillas por árbol provenientes de varias vainas maduras sin abrir (Fig.1). Para determinar indirectamente la importancia relativa de los procesos de dispersión y establecimiento, se midió el volumen ocupado por las estructuras relacionadas con ambos procesos. Los cotiledones y el embrión se consideraron como estructuras relacionadas con el establecimiento de la planta, mientras que el resto de la semilla -debido a sus adaptaciones para la flotación-, fue considerado como una estructura relacionada con la dispersión (Vásquez 1997). Se estimó el volumen total de la semilla (VT) midiendo el líquido desalojado (en ml) al sumergirla cuidadosamente en un envase graduado. Luego se abrió cada semilla y se determinó el volumen ocupado por los cotiledones y el embrión (volumen asignado a estructuras de establecimiento =VEE), utilizando la misma metodología (ver también Williamson et al. 1999). El volumen asignado a estructuras de dispersión (VED) fue calculado por diferencia, como VED =VT VEE. Con estos valores se calculó un índice de dispersión, ID =VED / VEE. Considerando al volumen ocupado de la semilla como un indicador de la inversión de la planta hacia la dispersión y el establecimiento, variaciones en el ID sugieren variaciones en la importancia relativa de ambos procesos. Si se analiza un ejemplo hipotético, se tiene que, si los árboles del hábitat B poseen un ID mayor que los ubicados en el hábitat A, esto sugiere que existe una mayor limitación para la dispersión en el hábitat B (o sea, la importancia relativa de la dispersión es allí mayor), ya que la selección natural estaría eliminando a los individuos cuyas semillas poseen poco espacio relativo asignado a estructuras de dispersión. Siguiendo el mismo razonamiento, este ejemplo hipotético sugiere una mayor importancia relativa del establecimiento en el hábitat A (en donde los árboles poseen ID menores), ya que se seleccionarían negativamente aquellos individuos con un mayor volumen ocupado por estructuras de dispersión en sus semillas, y por lo tanto, con una menor inversión relativa en estructuras de establecimiento (Fig.2). Por otra parte, si las presiones de selección hacia los procesos de dispersión y establecimiento son igualmente importantes en los diferentes hábitats, los árboles presentarán valores de ID semejantes, independientemente de su ubicación. En consecuencia, la distribución de los ID nos puede sugerir la existencia de selección disruptiva (i.e.,Fig.2) o estabilizante (Futuyma 1986).
La comparación de los ID entre árboles se hizo de dos formas. Primero, se calculó un ID promedio por árbol y se comparó su variación entre los tres hábitats estudiados mediante una prueba no paramétrica de Kruskal-Wallis (Zar 1999). Segundo, se compararon los ID entre árboles, independientemente de su ubicación, utilizando los ID de todas las semillas medidas por árbol (aproximadamente n =50) mediante un análisis de varianza de una vía, considerando a los árboles como factor aleatorio. En éste último caso los datos mostraron una distribución normal y homogeneidad de varianzas, por lo cual no fue necesario realizar ninguna transformación. A partir de la suma de cuadrados del análisis de varianza se calculó el porcentaje de variación del ID debido únicamente al efecto árbol. También calculamos el coeficiente de variación del ID entre árboles. Para determinar si variaciones en el ID dependían del tamaño de la semilla se realizó una regresión simple. Para analizar si la distribución del ID se ajustaba a una distribución normal, tanto a nivel de semillas como a nivel de individuos, se aplicaron pruebas de Wilk-Shapiro y se calculó el grado de asimetría de la distribución (Zar 1999).
]]> Adicionalmente, se llevó a cabo un pequeño experimento para verificar si variaciones en el ID o en el tamaño de las semillas determinaban cambios en su capacidad de dispersión. Se seleccionaron 40 semillas al azar, la mitad con un volumen total <20 ml (pequeñas) y la otra mitad >30 ml (grandes). Se formaron 20 pares al azar de semillas grandes y pequeñas, y en el río Sucusari se soltaron, desde un bote, cada par desde un punto fijo, para que flotaran a favor de la corriente. En total se realizaron 20 "pruebas de velocidad" y se registró cual semilla del par llegaba primero a una distancia de 220 cm, y la distancia recorrida por la otra semilla. Posteriormente se determinó para cada semilla su ID de la manera previamente detallada. Se aplicó una prueba pareada de Wilcoxon para comparar la distancia recorrida entre las semillas grandes y pequeñas, y una correlación de Pearson para determinar la relación entre el ID y la distancia recorrida.Resultados
C. angustifolia presenta frutos en forma de vainas cerradas (Fig.1A), en cuyo interior pueden encontrarse entre 4-7 semillas de diferente tamaño dependiendo de su posición dentro de la vaina (Fig.1B;Vásquez 1997). Estas semillas presentan claras adaptaciones para la dispersión por agua, ya que los cotiledones y el embrión se encuentran rodeados por un tejido esponjoso y la semilla posee un tegumento externo impermeable. La estrategia de flotación mediante un tejido esponjoso, de textura similar al corcho, ha sido descrita para varias especies de bosques inundables tropicales. Esta estructura permite la flotación a largo plazo de la semilla facilitando su dispersión (Kubitzki y Ziburski 1994, Williamson et al .1999, López 2001).
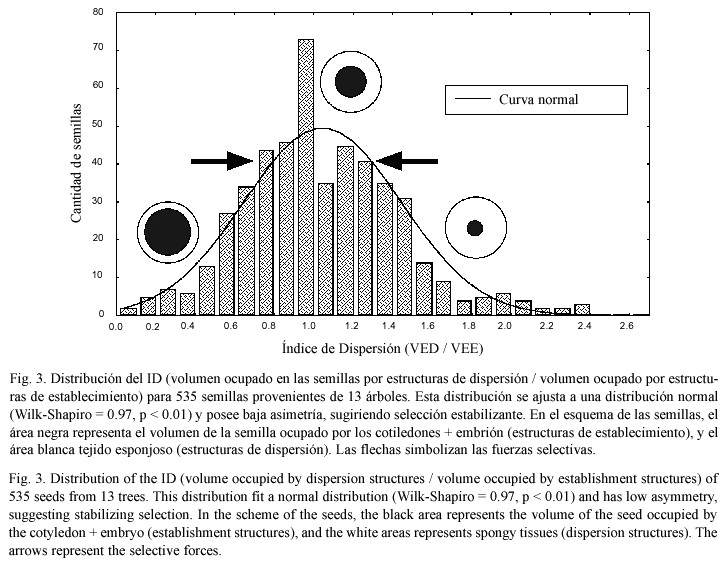
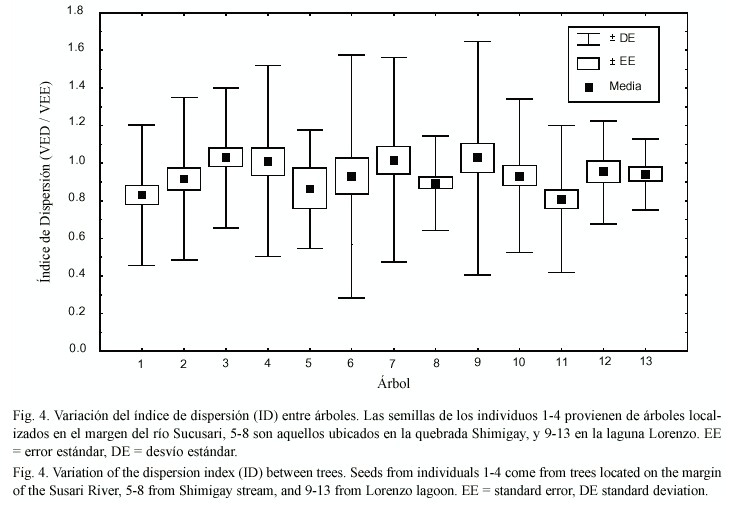
Se midió un total de 535 semillas provenientes de 13 árboles, las cuales presentaron una gran variación en su volumen total, desde 5 hasta 81 ml. Sin embargo, tanto semillas grandes como pequeñas tuvieron una relación entre el volumen asignado a estructuras de establecimiento y a estructuras de dispersión similar (ID ~1), por lo cual, variaciones en el tamaño total de las semillas no determinaron variaciones en el ID (F1,533 =2.7,p >0.05, R2 =0.5%). Por otra parte, el ID presentó una distribución normal con baja asimetría tanto agrupando todas las semillas (Wilk-Shapiro test, W =0.93, p <0.01, n =535, promedio (± 1 EE)= 0.91 ±0.01, asimetría =0.5;Fig.3), como al considerar el ID promedio por árbol (W =0.97,n =13,p <0.01, promedio (±1 DE) =0.94 ±0.01,asimetría =0.2). Por otra parte, el ID promedio fue similar entre hábitats (KW =0.38,p =0.82). Los árboles ubicados en la margen del río Sucusari presentaron un ID promedio (±1 DE) de 0.95 ±0.09, los ubicados en la quebrada Shimigay 0.93 ± 0.07, y los localizados en la Laguna Lorenzo 0.93 ±0.08. Consistentemente con este resultado, el ID fue similar entre árboles, independientemente del hábitat de donde provenían los individuos (F1,12 =1.26,p =0.23;Fig.4). Solo el 2.7% de la variación total del ID fue explicada por la variación encontrada entre árboles, y el ID presentó un coeficiente de variación promedio muy bajo (7.8%). Por otra parte, tanto semillas grandes (>30 ml) como pequeñas (<20 ml) tuvieron velocidades similares de dispersión en el agua (W =37,n =20 pares,p =0.90). Sin embargo, encontramos una relación positiva entre el ID y la distancia recorrida en un tiempo fijo (r =0.51, gl =18,p =0.02).
Discusión
La relación entre el volumen asignado a estructuras de dispersión y establecimiento fue similar entre semillas de diferente tamaño, y entre árboles pertenecientes a diferentes hábitats. Esta relación tuvo una distribución normal con baja asimetría tanto agrupando todas las semillas o usando a los árboles como réplicas, indicando que esta relación está sometida a una selección estabilizante (Fig.3;ver Futuyma 1986). Adicionalmente, los árboles presentaron una variación muy baja del ID entre individuos (Fig.4). Esto sugiere que, al menos en los hábitats muestreados, tanto la dispersión como el establecimiento tendrían una importancia relativa similar para C.angustifolia ,independientemente del tamaño de sus semillas o de la ubicación de sus individuos.
Esta interpretación se basa en suponer que la relación entre los volúmenes ocupados por determinadas estructuras es un buen estimador de la importancia relativa de sus funciones; y en consecuencia, que la capacidad de dispersarse y establecerse mejora cuanto más volumen relativo de sus semillas es ocupado por las estructuras relacionadas con estos procesos. En otras palabras, que para determinado tamaño de semilla, más volumen de tejido esponjoso favorece la flotación, y que cotiledones más grandes favorecen la supervivencia de la plántula.
Por un lado, este trabajo ha demostrado que, independientemente de su tamaño, semillas con más espacio ocupado por tejido esponjoso tuvieron una mayor capacidad de dispersión. Pese a que la velocidad de dispersión no necesariamente ha sido considerada un carácter adaptativo en las semillas hidrocóricas debido a que su destino depende de las corrientes (Dirzo y Domínguez 1986, Schneider y Sharitz 1988), el tiempo de flotación se considera una buena medida de la eficiencia de la dispersión (Williamson et al. 1999, Williamson y Costa 2000,López 2001). Dado que las semillas con dispersión hidrocórica no pueden germinar en el agua, un mayor tiempo de flotación incrementa la probabilidad de que sean depositadas en sitios más favorables para su germinación y establecimiento (Schneider y Sharitz 1988,López 2001). Es muy posible que las semillas con un mayor volumen relativo ocupado por estructuras esponjosas, las cuales en este estudio demostraron dispersarse más rápido, también floten mas tiempo.
Por otra parte, pese a no estar medido directamente en este trabajo, muchos otros estudios han demostrado una relación positiva entre el tamaño de las estructuras de establecimiento y la supervivencia de la plántula, tanto en especies de bosques de tierra firme (Augspurger 1984, Howe y Smallwood 1982, Ganeshaiah y Uma Shaanker 1991,Westoby et al.1992,Dalling et al.1998),como de bosques inundables (Jhones et al.1994,López 2001).
Si, tal como sugieren los resultados de este estudio, en C.angustufolia la relación entre el volumen de semilla asignado a estructuras de dispersión y establecimiento se encuentra fijado por selección estabilizante (Fig.3), desvíos de esta relación deberían ser negativamente seleccionados. Por ejemplo, un mayor volumen neto en estructuras de dispersión para un tamaño fijo de semilla implicaría disminuir el tamaño neto de los cotiledones. Es conocido que esta disminución afecta negativamente la supervivencia de la plántula por disminuir (a) el aporte de carbono en ambientes con baja luminosidad (Walters y Reich 2000), (b)el desarrollo de raíces (Westoby et al.1982), (c) su resistencia ante daños físicos ocasionados por fragmentos flotantes (Foester 1986),y (d) la capacidad de tolerar depredación en sus semillas (Mack 1998). Esta última característica podría ser importante en C. angustifolia, dado que el 80% de sus semillas son atacadas por larvas de insectos cuando se encuentran en tierra firme (A.Valerio, sin publicar). Por otro lado, producir cotiledones más grandes para un tamaño fijo de semilla implica una disminución del volumen asignado a estructuras de dispersión, reduciendo la flotabilidad de la semilla (Williamson y Costa 2000,Lopez 2001). En el caso de C.angustifolia, la flotabilidad de sus semillas no solo es importante para permitirle colonizar nuevos ambientes sino para escapar de sus depredadores, ya que en agua solo el 2% sufre depredación, y en tierra sufren entre 3-10 veces más ataques por larvas de insectos cuando están más cerca que lejos del árbol madre (A.Valerio, sin publicar). En consecuencia, una mayor inversión en estructuras relacionadas con la dispersión en detrimento de las estructuras relacionadas con el establecimiento (o viceversa) sería "penado "por la selección natural, manteniéndose un equilibrio entre ambos procesos en el uso del espacio de la semilla (llamativamente, en este caso un 50% cada uno; Fig.3). La baja variación del ID (~3%) entre árboles (en definitiva la unidad de selección), refuerza la hipótesis de que esta relación está sometida a selección estabilizante. Las características de las semillas que favorecen la flotación y su supervivencia luego de la germinación han sido propuestas ambas como los factores más importantes en promover el establecimiento exitoso en las especies de bosques inundables (López 2001). Sería interesante, entonces, determinar si el balance encontrado en este estudio es característico de las especies con dispersión por hidrocoria.
Por otra parte, se ha propuesto que el mantenimiento de la variación existente en el tamaño de las semillas es el producto de una solución de compromiso entre la selección para la dispersión, la cual favorece semillas pequeñas, y la selección para el establecimiento, la cual favorece semillas grandes (Ganeshaiah y Uma Shaanker 1991,Westoby et al.1992, Dalling y Hubbell 2002). Sin embargo, contrariamente a lo que sucede en otros tipos de dispersión abiótica (e.g.,viento, ver Greene y Johnson 1993), algunos estudios con especies hidrocóricas han encontrado una mayor dispersión en semillas grandes, sugiriendo que la variación en el tamaño de las semillas es el resultado de una solución de compromiso entre dos estrategias de dispersión: dehiscencia y flotación. Por un lado, las semillas grandes, más efectivas para dispersarse a largas distancias, funcionarían mejor para colonizar hábitats alejados. Por otra parte, las semillas pequeñas, que se hunden en pocos días, serían más efectivas para colonizar áreas cercanas al árbol madre, en donde habría baja competencia debido al efecto de las inundaciones (Williamson et al.1999, Williamson y Costa 2000).
]]> Sin embargo este no parece ser el caso de C.angustifolia. La flotabilidad de sus semillas no estuvo determinada por su tamaño, sino por el volumen relativo ocupado por tejido esponjoso. Adicionalmente, semillas cerca de la planta madre sufrieron mayor depredación por larvas de insectos (A. Valerio, sin publicar), sugiriendo que dispersarse a distancias cortas es desventajoso. Posiblemente, en esta especie la variación en el tamaño de la semilla se deba a la limitación física de su desarrollo en los extremos de la vaina, la cual está cerrada y en consecuencia limita el tamaño de las semillas terminales (Williamson y Costa 2000). Este trabajo sugiere que, para especies con semillas empaquetadas, el volumen relativo ocupado por estructuras relacionadas con la dispersión o el establecimiento podría ser una medida más adecuada de la solución de compromiso entre estos procesos que la variación del tamaño de la semilla per se .Agradecimientos
A dos revisores anónimos y Marcelo Aizen por sus comentarios, y a la Organización para Estudios Tropicales (OET /OTS)por su apoyo logísico y técnico.
Resumen
Evaluamos indirectamente las presiones selectivas sobre la dispersión y el establecimiento en Campsiandra angustifolia, un árbol de la Amazonía Peruana dispersado por agua, analizando variaciones de la relación entre el volumen ocupado por las estructuras de dispersión y de establecimiento en sus semillas. Medimos un total de 535 semillas de 13 árboles ubicados en tres hábitats diferentes, las cuales presentaron una gran variación en su volumen total. Independientemente del tamaño de la semilla y de la ubicación del árbol de origen, la relación entre el volumen asignado a estructuras de establecimiento y a estructuras de dispersión fue relativamente constante (~1) y presentó una distribución normal con baja asimetría, indicando selección estabilizante. Este resultado sugiere que los procesos de dispersión y establecimiento poseen una importancia relativa similar para C.angustifolia en los hábitats estudiados. En especies con semillas empaquetadas, el volumen relativo ocupado por estructuras relacionadas con la dispersión o el establecimiento podría ser una medida más adecuada de la solución de compromiso entre estos dos procesos que la variación del tamaño de la semilla.
Palabras clave: Amazonía, bosques inundables, dispersión, establecimiento, hidrocoria.
Referencias
Augspurger, C. 1983. Seed dispersal of the tropical tree, Platypodium elegans, and the escape of its seedlings from fungal pathogens. J. Ecol. 71:759-771. [ Links ]
Augspurger, C. 1984. Seedling survival of tropical tree species; interactions of dispersal distance, light gaps and pathogens. Ecology 65:1705-1712. [ Links ]
Augspurger, C.& P. Hogan. 1983. Wind dispersal of fruits with variable seed number in a tropical tree (Lonchocarpus pentaphyllus :Leguminosae). Amer. J. Bot. 70:1031-1037. [ Links ]
Coley, P., J. Bryant & F. Chapin III. 1985. Resource availability and plant anti-herbivory defense. Science 230: 895-899. [ Links ]
Dalling, J. & S. P. Hubbell. 2002. Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species. J. Ecol. 90: 557-568. [ Links ]
Dalling, J., S. P. Hubbell & K. Silvera. 1998. Seed dispersal, seedling establishment and gap partitioning among tropical pioneer trees. J. Ecol. 86:674-689. [ Links ]
Dirzo, R. & C. Domínguez. 1986. Seed shadows, seed predation and the advantages of dispersal, p. 237-249. In A. Estrada & T. Fleming (eds.). Frugivores and Seed Dispersal, Junk. [ Links ]
Foster, S. 1986. On the adaptive value of large seeds for tropical moist forest trees: a review and synthesis. Bot. Rev. 52:260-299. [ Links ]
Futuyuma, D. J. 1986. Evolutionary Ecology. Sinauer, Massachusetts. [ Links ]
Ganeshaiah, K. & R. Uma Shaanker. 1991. Seed size optimization in a wind dispersed tree Butea manosperma : a trade-off between seedling establishment and pod dispersal efficiency. Oikos 60:3-6. [ Links ]
Greene, D. & E. Johnson. 1993. Seed mass and dispersal capacity in wind-dispersed diaspores. Oikos 67: 69-74. [ Links ]
Grime, J. 1979. Plant Strategies and Vegetation Processes. Wiley, Chichester. [ Links ]
Harper, J. L. 1977. The Population Biology of Plants. Academic, London. [ Links ]
Herms, D. & W. Mattson. 1992. The dilemma of plants: to grow or defend. Quart. Rev. Biol. 67:283-335. [ Links ]
Howe, H. & J. Smallwood. 1982. Ecology of seed dispersal. Ann. Rev. Ecol. Syst. 13:201-228. [ Links ]
Jhones, R., R. Sharitz, P. Dixon, D. Segal & R. Schneider. 1994. Woody plant regeneration in four flood plain forest. Ecol. Monogr. 64:345-367. [ Links ]
Kubitzi, K. & A. Ziburski. 1994. Seed dispersal in flood plain forest of Amazonia. Biotropica 26:30-43. [ Links ]
López, O. 2001.Seed flotation and post flooding germination in tropical terra firme and seasonally flooded forest species. Funct. Ecol. 15:763-771. [ Links ]
Mack, A. 1998. An advantage of large seed size:tolerating rather succumbing to seed predators. Biotropica 30: 604-608. [ Links ]
Schneider, R., & R. Sharitz. 1988. Hydrocory and regeneration in a bald cypress-water tupelo swamp forest. Ecology 69:1055-1063. [ Links ]
Thompson, J. 1999. Specific hypotheses on the geographic mosaic of coevolution. Amer. Nat. 153:S1-S14. [ Links ]
Vásquez, R. 1997. Flórula de las Reservas Biológicas de Iquitos, Perú. Missouri Botanical Garden. 1046 pp. [ Links ]
Walters, M. & P. Reich. 2000. Seed size, nitrogen supply, and growth rate affect tree seedling survival in deep shade. Ecology 81:1887-1901. [ Links ]
Westoby, M., E. Jurado & M. Leishman. 1992. Comparative evolutionary ecology of seed size. Tree 7:368-372. [ Links ]
Williamson, B. & F. Costa. 2000. Dispersal of Amazonian trees: hydrocory in Pentaclethra macroloba. Biotropica 32:548-552. [ Links ]
Williamson B., F. Costa & C. Minte Vera. 1999. Dispersal of Amazonian tress: hydrocory in Swartzia polyphylla. Biotropica 31:460-465. [ Links ]
Zar, J. 1999. Biostatistical analysis. 4 th ed. Prentice-Hall, New Jersey, 663 p. [ Links ]
]]>