Morphological and physiological responses of seagrasses (Alismatales) to grazers (Testudines: Cheloniidae) and the role of these responses as grazing patch abandonment cues
Elizabeth A. Lacey1*, Ligia Collado-Vides2* & James W. Fourqurean2 ]]>
Abstract
Green sea turtles, Chelonia mydas, are grazers influencing the distribution of seagrass within shallow coastal ecosystems, yet the drivers behind
C. mydas patch use within seagrass beds are largely unknown. Current theories center on food quality (nutrient content) as the plant responds to grazing disturbances; however, no study has monitored these parameters in a natural setting without grazer manipulation. To determine the morphological and physiological responses potentially influencing seagrass recovery from grazing disturbances, seagrasses were monitored for one year under three different grazing scenarios (turtle grazed, fish grazed and ungrazed) in a tropical ecosystem in Akumal Bay, Quintana Roo, Mexico. Significantly less soluble carbohydrates and ]]>
Thalassia testudinum were indicative of the stresses placed on seagrasses during herbivory. To determine if these physiological responses were the drivers of the heterogeneous grazing behavior by C. mydas recorded in Akumal Bay, patches were mapped and monitored over a six-month interval. The abandoned patches had the lowest standing crop rather than leaf ]]>
C. mydas grazing. This study is the first to apply GUD theory, often applied in terrestrial literature, to explain marine ]]>
Key words: seagrass, Thalassia testudinum, patch abandonment, Chelonia mydas, herbivory, grazing behavior, giving up density (GUD).
Resumen
Respuestas morfológicas y ]]>
Chelonia mydas, son herbívoros que influencian la distribución de pastos marinos en sistemas costeros someros, sin embargo los factores que hay detrás del uso de un parche de pastos marinos por C. mydas son desconocidos. Teorías actuales se centran en calidad de alimento (contenido de nutrientes en tejido) conforme la planta responde a la alteración causada por el pastoreo; sin embargo no hay estudios que hayan monitoreado estos parámetros en un diseño natural sin manipulación del herbívoro. Para deter minar las respuestas morfológicas y fisiológicas (productividad, morfología y almacenamiento de nutrientes) que ]]>
C. midas, reportado en Akumal, se mapearon y monitorearon parches de pastos en intervalos de seis meses. Los parches abandonados tuvieron menor biomasa, en vez de bajo contenido de nutrientes en hoja, o de carbohidratos en raíces. Estos resultados sugieren un comportamiento modificado de renunciamiento por densidad: el umbral crítico donde el costo de pastoreo continuo no provee los nutrimentos mínimos, por lo tanto nuevos parches deberán ser utilizados, explicando así el abandono del recurso y mecanismo detrás del comportamiento de pastoreo por C. mydas. Este es el primer estudio en aplicar la teoría de renuncia por densidad, frecuentemente utilizada en la literatura terrestre, para explicar el comportamiento de pastoreo de herbívoros.
Palabras clave: pastos marinos, Thalassia testudinum, abandono de parche, Chelonia mydas, herbívoro, pastoreo, renunciamiento por densidad (GUD).
Herbivores are important drivers of ecosystem dynamics (Heck & Valentine, 2007; Valentine & Duffy, 2006) and seagrass productivity (Valentine, Heck, Busby, & Webb, 1997; Zieman, Iverson, & Ogden, 1984), reducing canopy cover and freeing space for competitive interactions between macrophytes (Heck & Valentine, 2006). Grazing by herbivores is not uniform as they selectively feed in certain areas depending on abiotic and biotic factors. This patchy grazing behavior opens space for successional dynamics heterogeneously throughout the seagrass ecosystem, changing ]]>
Grazing disturbances by marine herbivores varies in intensity and ]]>
pers observation). Green sea turtles (Chelonia mydas Linnaeus 1758) grazing behavior differs from fishes in that they repeatedly graze distinct patches of seagrass, which optimizes food quality, as new leaves have higher nutrient content than old leaves (Aragones, Lawler, Foley, & Marsh, 2006; Bjorndal, 1980). The drivers behind the creation of these grazing patches, as ]]>
When the potential harvest rate of thin, sparse, slow-growing seagrasses by sea turtles can no longer balance the associated metabolic costs of foraging, sea turtles should abandon grazing in those patches for areas with higher potential harvest rates, the concept of Giving Up Density (GUD) (Brown, 1988). Giving Up Density theory extends the optimal foraging theory and ]]>
Our study was designed to evaluate the effect of different types of herbivory (fish and turtle) on morphological and physiological characteristics of seagrasses. We also wanted to determine which of the monitored seagrass characteristics may ]]>
C. mydas, we predict that patches abandoned would have ]]>
Materials and Methods
Study site: Akumal (“Place of the Turtles” in Mayan) is located in Quintana Roo, Mexico, in the Yucatan Peninsula along the Caribbean ]]>
Chelonia mydas and near the interspersed patch reefs, members of the family Scaridae (parrotfish) and Acanthuridae (surgeonfish). Seagrass beds consist of Thalassia ]]>
(Banks ex König), Halodule wrightii (Ascherson) and Syringodium filiforme (Kuetz), with few intermixed macroalgae typified by calcareous Chlorophytes (e.g., Halimeda, Udotea) and calcareous Rhodophytes (e.g,. Neogoniolithon).
Seagrass response: To determine the species specific response of seagrasses to herbivory, indicators of physiological responses were monitored in turtle and fish grazed and ungrazed patches in the seagrass beds. Distinct patches exist where seagrass beds have been grazed (by either turtle or fish) or not grazed, and within these patches small cages (20cm diameter) were haphazardly placed for six days at a time during two growing seasons ]]>
T. testudinum only), leaf morphology, tissue nutrients and soluble carbohydrates of all species present (n=12, 6, 12 in winter for ungrazed, fish grazed and turtle grazed, respectively; n=11, 10, 8 in summer for ungrazed, fish grazed and turtle grazed, respectively). Cages were created with vinyl-coated hardware mesh (ca. 1cm mesh) and four 3mm diameter vinyl-coated metal stakes secured the cage into the substrate. These cages were not intended to be experimental exclosures; rather, they allowed us to measure the production of material for ]]>
2 patch reef adjacent to the same seagrass beds where turtle grazing occurred. Fish herbivory was confirmed by the patterns of bite marks on leaves, which were indicative of parrotfish grazing. Turtle herbivory was confirmed during visual census of the study area.
]]>
To measure productivity, morphology and physiological status of seagrass within these cages, all T. testudinum blades were punched using a modified leaf marking technique (Fourqurean, Willsie, Rose, & Rutten, 2001; Zieman, 1974). At the end of six days, aboveground and belowground biomass was collected for all seagrass species present ]]>
T. testudinum were measured for length, width and elongation rate (mm/shoot/ day), from which elongation area (width x length of new growth at base of leaf) was calculated. All seagrass samples were cleaned of epiphytes, dried to a constant weight at 60ºC and ground to a fine powder using a mortar and pestle. Samples were ]]>
H. wrightii and S. filiforme ]]>
In order to test for the hypothesized differences between herbivory patches and seasons on ]]>
H. wrightii and S. filiforme in grazed cages, sample sizes varied between patches and seasons. Therefore, to test for differences between herbivory patches and between seasons, nutrient content and soluble ]]>
T. testudinum), two-way ANOVA (S. filiforme) or via a student’s t-test or Mann-Whitney nonparametric test (H. ]]>
), depending on how the data met normality assumptions. No S. filiforme was present during the winter in the fish grazed patch and no H. wrightii was present in the fish ]]>
Patch dynamics: To monitor changes in turtle grazed or ungrazed patch location and seagrass morphological and physiological changes in relation to changes in patch designation over time, patch locations in the seagrass ecosystem were identified and monitored using belt transects. Ten transects were established ]]>
2 plot placed haphazardly within each patch. For each macrophyte BB score, 1 indicated coverage less than 5% of the plot, 2 indicated 5-25% cover, 3 indicated 25-50% cover, 4 indicated 50-75% cover; and 5 indicated 75-100% cover. The presence of a different patch along a transect was recorded when grazing status (turtle grazed or ungrazed) or when BB score differed from adjacent ]]>
Braun-Blanquet scores were converted to abundances using the mean abundance within each score (e.g., 2.5% cover for score 1, 87.5% cover for score 5) and then averaged across similarly designated patches (e.g., all turtle grazed patches). Mean abundances are presented for turtle grazed and ungrazed patches during both surveys for comparisons between surveys. A ]]>
T. ]]>
and seagrass blade length for all species was recorded. In the second survey, patches labeled ungrazed had been ungrazed for six months or more while abandoned patches were recently ungrazed (less than six months since being grazed). An abandoned patch was a patch that was grazed by turtles during the first survey and whose blade lengths had increased and blade apexes were intact for the second survey. Following the second survey, a Student’s t-test was used to test for differences in proportions of each transect in each status ]]>
T. testudinum and S. filiforme (Fourqurean et al., 2001) and tested for signifi- cance using an ANOVA. ]]>
At 15 randomly chosen turtle grazed and ungrazed patches along the transects, six randomly selected shoots of T. testudinum were collected to determine the change in response variables between sampling intervals as a function of patch status following the methods outlined above. An ANOVA was used to test for ]]>
T. testudinum parameters (width, nitrogen, phosphorus, soluble carbohydrates) among patch status in the second survey (remain ungrazed, remain turtle grazed, abandoned) after examination of standardized residuals showed the data conformed to all normality assumptions. A Student’s t-test was used to test for differences in initial survey data between patches designated during the second survey as grazed and abandoned. ]]>
Results Seagrass response: While cages were deployed on two separate occasions during each season, ]]>
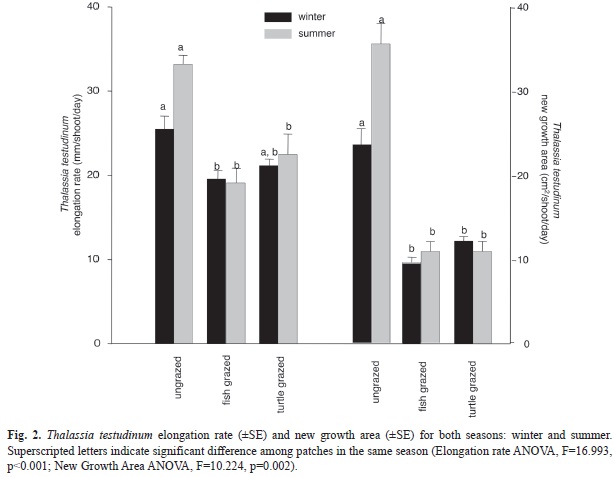
Thalassia testudinum blades (ANOVA F=12.06, p<0.001) with fastest elongation rate (ANOVA F=16.99, p<0.001) and largest new growth area (ANOVA F=10.22, p=0.002), when compared to fish and turtle grazed patches (
Fig. 1 and 2). The difference between ungrazed and grazed was greater in the summer months and significantly higher in ungrazed patches compared to grazed patches, regardless of grazer identity in both seasons (p<0.001 for all three parameters). Fish grazed and turtle grazed patches were significantly different from each other in width during the winter (p<0.001), but were not significantly different for other parameters or within the same patch between seasons. Grazed shoots had a maximum of three leaves, while ]]>
T. testudinum and occasionally S. filiforme
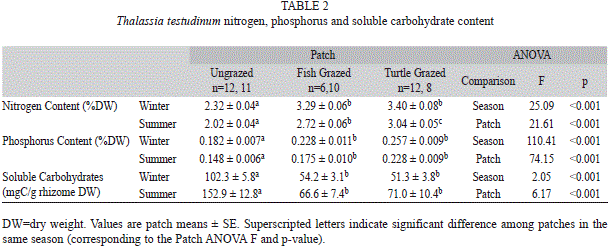
were present in grazed patches. Ungrazed patches of T. testudinum had lower nitrogen and phosphorus content than turtle and fish grazed patches in both seasons (Table 2). Soluble carbohydrates in T. testudinum rhizomes were significantly higher in the ungrazed patches for both seasons and were significantly higher in summer compared to winter in all three patches (Table 1, 2).
Similar significant ]]>
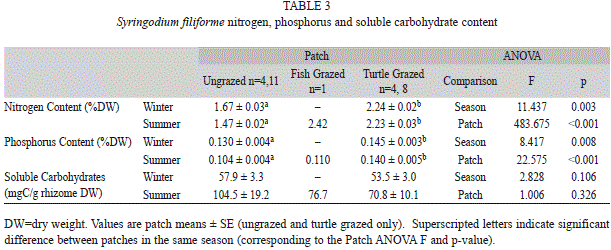
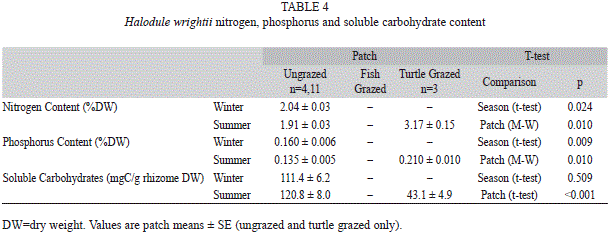
T. testudium were found in Syringodium filiforme, for which ungrazed patches had significantly lower nitrogen and phosphorus content than turtle grazed patches (Table 1, 3, sample size was too small to test for differences from fish grazed patches). For Halodule wrightii, significant differences were found in nitrogen and phosphorus ]]>
Table 1, 4). There was significantly higher nitrogen and phosphorus content in winter compared to summer in the ungrazed patches, similar to the trend found in T. testudinum and S. filiforme. Soluble carbohydrates of rhizomes were significantly higher in ungrazed patches than turtle grazed patches (Table 1), similar to T. testudinum. Grazed shoots had a maximum of ]]>
Patch dynamics: The majority of seagrass patches recorded during the initial survey were ungrazed by turtles (63%), while the remaining patches were turtle grazed. The percent of ungrazed patches did not significantly change during the second survey (Student’s t-test, ]]>
Table 5). ]]>
In the second survey, comparisons could only be made between ungrazed, turtle grazed and abandoned patches, as sample size of newly grazed patches was too small (n=1) for statistical comparison. Ungrazed patches had the highest abundance of T. testudinum and S. filiforme than grazed and abandoned patches (ANOVA, p=0.015 and p=0.037, respectively), while H. wrightii abundance was highest in abandoned patches (ANOVA, p=0.008) and increased significantly from initial to second survey (t-test, p<0.001). Syringodium filiforme and T. testudinum abundance did not significantly change for abandoned patches from initial to second survey (t-test, p=0.912). Two species of macroalgae not observed in the initial survey occurred within the one aban- doned plot ]]>
Halimeda sp., Penicillus sp.). In the one newly grazed patch, there was one less seagrass species present, but the small sample size did not allow statistical comparisons.
Within the fifteen transect patches of
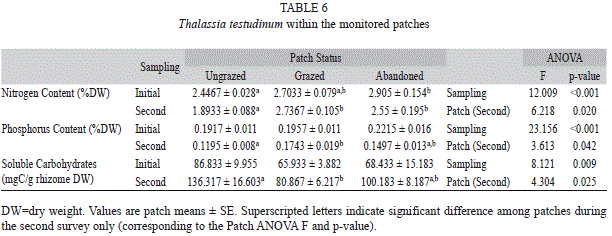
T. testudinum sampled for nutrients, soluble carbohydrates and morphology during the initial survey, ungrazed patches had significantly lower nitrogen and phosphorus content (Table 6; t-test, p<0.001 for both parameters) and significantly higher soluble sugar content (t-test, p=0.044) than grazed patches, similar to those results found in the cages. Phosphorus content and soluble ]]>
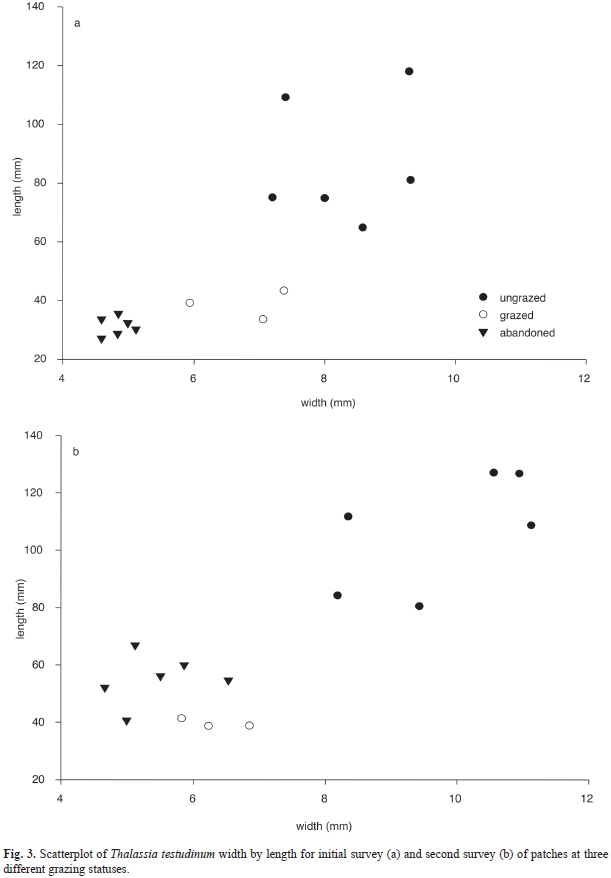
Thalassia testudinum. Prior to abandonment, leaves were short and narrow, and a distinct shift in these parameters followed abandonment (Fig. 3). Standing crop ]]>
2, while grazed patches averaged significantly greater at 62.9g/m2, and ungrazed patches averaged significantly greater than abandoned and grazed patches at 99.7g/m2 (ANOVA, p<0.001).
Discussion Monitoring of patch status provided an opportunity to look at the changes in response variables (nutrient content, morphology, physiology) as a function of patch type. The driver(s) behind changes in patch use are elusive, as monitoring of Chelonia mydas patch use through time ]]>
C. mydas to consume enough plant biomass to fulfill their nutritional needs could outweigh the nutritional reward, and therefore induce abandonment of the patch by feeding turtles, similar to the mechanism regulating patch use by herbivores reported in terrestrial ecosystems (Brown, 1988). ]]>
In our system, patch abandonment was not driven by the quality of seagrass as would be reflected in nutrient content, nor by stored soluble carbohydrates, but cued directly by the low quantity provided by the sparse, thin seagrass blades of the heavily grazed patches. Based on the biomass found in grazed patches during the initial survey that were later abandoned, we propose a threshold of approximately 17g/m2, below which patches can be predicted ]]>
]]>
The relative physiological (productivity, tissue nutrients and soluble carbohydrates) and morphological effects of grazing on seagrasses were similar to those previously measured from simulated grazing or exclosure experiments (Fourqurean et al., 2010; Moran & Bjorndal, 2005, 2007). Leaf widths of fish grazed (4.9-5.4mm) and turtle grazed (5.0-5.9mm) patches in this study were within range of other studies on grazing (5.3-6.3mm, Zieman et al., 1984; 5.6-6.3mm, ]]>
Smaller sample sizes ]]>
Syringodium filiforme and Halodule wrightii limited our determination of significant differences in nutrient and soluble carbohydrate content between patches but their absence in grazed patches may indicate herbivore preference (Armitage & Fourqurean, 2006). The absence of certain species may also indicate a lower tolerance to repeated or ]]>
S. filiforme during the winter may decrease the ability of this species to recover from grazing. Larger species with thicker rhizomes, such as T. testudinum
, may also be better at translocating their stored carbohydrates longer distances and are therefore better able to compensate for loss than those species with thinner rhizomes, e.g., H. wrightii and S. filiforme (Marba, ]]>
]]>
Over a six month interval, the location of turtle grazed patches was largely stable, with only a small fraction of area changing status. Our data indicate grazing patches were long- lived enough to affect soluble carbohydrate storage, morphology and nutrient content of seagrass leaves. As expected during the initial survey, turtle herbivory reduced seagrass abundance and canopy complexity (macrophyte species composition and blade length and width). During the second survey, after patches were abandoned, ]]>
Halodule wrightii) increased canopy complexity. The abundance of this species in recently abandoned patches is consistent with successional dynamics reported for other types of seagrass ecosystems, especially in nutrient-replete systems (Duarte, 1995; Fourqurean, Powell, Ken-worthy, & Zieman, 1995). Although we cannot statistically test changes in macroalgae as a result of the small sample size, the one ]]>
Halimeda during the initial study did have a new genus, Penicillus, during the second survey. It was surprising that macroalgae were scarce throughout grazed patches, where canopy cover was decreased and potential competitive interactions increased, or in ungrazed patches, where macroalgae may potentially find refuge within the complex canopy. The absence of macroalgae could be due to limited algal reproductive ]]>
Impacts of grazers on different seagrass species may also be an important determinant of primary producer distribution, as found in other studies (Armitage & ]]>
Halodule wrightii over slower-growing, climax species such as T. testudinum (Armitage & Fourqurean, 2006; ]]>
T. testudinum (Mortimer, 1981), which may be an artifact of the lower availability and abundance of H. ]]>
relative to other species as well as spatial variability between grazing populations and habitat types (Bjorndal, 1997). This is the first study to simultaneously consider the impact of turtle and fish grazing behaviors on morphological and physiological parameters important to seagrass growth and recovery (e.g., productivity, blade width).
Considering the ]]>
Halodule wrightii and Syringo- dium filiforme (Armitage & Fourqurean, 2006; Burkholder, Heithaus, & Fourqurean, 2012), the presence of these species in the ungrazed beds may provide a source for recolonization by these species once patches are abandoned. Similar responses have been reported after other disturbances ]]>
H. wrightii can replace T. testudinum under conditions of high nutrient availability (Fourqurean et al., 1995), if no source for H. wrightii propagules exists, the ecosystem may transition to bare sand rather than supporting a macrophyte assemblage. Alternatively, this study suggests that heterogeneous grazing may promote a more species-rich seagrass community in a eutrophic ecosystem.
Our study determined ]]>
The application of this theory provides novel insight on the behavior of mesograzers and the crossover between terrestrial and aquatic theories on grazing yield/grazing effort or risk. The application of GUD theory should be examined in other sites experiencing patch abandonment by green sea turtles in order to test this proposed threshold. ]]>
Acknowledgments We are grateful to Centro Ecológico Akumal staff and volunteers for field support and logistics, in particular Y. Kleinschmidt and B. van Tussenbroek from Universidad Nacional Autónoma de México provided research support, and suggestions from J. Rehage ]]>
References ]]>
Aragones, L. V., Lawler, I. R., Foley, W. J., & Marsh, H. (2006). Dugong grazing and turtle cropping: grazing optimization in tropical seagrass systems? Oecologia, 149, 635-647. [ Links ]
Armitage, A. R. & Fourqurean, J. W. (2006). The shortterm influence of herbivory near patch reefs varies between seagrass species. Journal of Experimental Marine Biology and Ecology, 339(1), 65-74. [ Links ]
Armitage, A. R., Frankovich, T. A., & Fourqurean, J. W. (2011). Long-term effects of adding nutrients to an oligotrophic coastal environment. Ecosystems, 14(3), 430-444. [ Links ]
Bjorndal, K. A. (1980). Nutrition and grazing behavior of the green turtle Chelonia mydas. Marine Biology, 56, 147-154. [ Links ]
Bjorndal, K. A. (1997). Foraging ecology and nutrition of sea turtles. In P. L. Lutz & J. A. Musick (Eds.), The biology of sea turtles (pp. 432). Boca Raton, FL: CRC Press. [ Links ]
Brown, J. S. (1988). Patch use as an indicator of habitat preference, predation risk, and competition. Behavioral Ecology and Sociobiology, 22, 37-47. [ Links ]
Burkholder, D. A., Heithaus, M. R., & Fourqurean, J. W. (2012). Feeding preferences of herbivores in a relatively pristine subtropical seagrass ecosystem. Marine and Freshwater Research, 63, 1051-1058. [ Links ]
Dawes, C. J., Bird, K., Durako, M., Goddard, R., Hoffman, W., & McIntosh, R. (1979). Chemical fluctuations due to seasonal cropping effects on an alga-seagrass community. Aquatic Botany, 6, 79-86. [ Links ]
Duarte, C. M. (1995). Submerged aquatic vegetation in relation to different nutrient regimes. Ophelia, 41, 87-112. [ Links ]
Fourqurean, J. W., Manuel, S., Coates, K. A., Kenworthy, W. J., & Smith, S. R. (2010). Effects of excluding sea turtle herbivores from a seagrass bed: overgrazing may have led to loss of seagrass meadows in Bermuda. Marine Ecology Progress Series, 419, 223-232. [ Links ]
Fourqurean, J. W., Powell, G. V. N., Kenworthy, W. J., & Zieman, J. C. (1995). The effects of long-term mani- pulation of nutrient supply on competition between the seagrasses Thalassia testudinum and Halodule wrightii in Florida Bay. Oikos, 72, 349-358. [ Links ]
Fourqurean, J. W., Willsie, A. W., Rose, C. D., & Rutten, L. M. (2001). Spatial and temporal pattern in seagrass community composition and productivity in south Florida. Marine Biology, 138, 341-354. [ Links ]
Heck, J. K. L. & Valentine, J. F. (2006). Plant-herbivore interactions in seagrass meadows. Journal of Experimental Marine Biology and Ecology, 330, 420-436. [ Links ]
Heck, J. & Valentine, J. F. (2007). The H.T. Odum synthe sis essay: The primacy of top-down effects in shallow benthic ecosystems. Esutaries and Coasts, 30(3), 371-381. [ Links ]
Kuiper-Linley, M., Johnson, C. R., & Lanyon, J. M. (2007). Effects of simulated green turtle regrazing on seagrass abundance, growth and nutritional status of Moreton Bay, south-east Queensland, Australia. Marine and Freshwater Research, 58, 492-503. [ Links ]
Lal, A., Arthur, R., Marba, N., Lill, A. W. T., & Alcoverro, T. (2010). Implications of conserving an ecosystem modifier: increasing green turtle (Chelonia mydas) densities substantially alters seagrass meados. Biological Conservation, 143, 2730-2738. [ Links ]
Lee, K. S. & Dunton, K. H. (1997). Effects of in situ light reduction on the maintenance, growth and partitioning of carbon resources in Thalassia testudinum Banks ex Konig. Journal of Experimental Marine Biology and Ecology, 210, 53-73. [ Links ]
Marba, N., Santiago, R., Diaz-Almela, E., Alvarez, E., & Duarte, C. M. (2006). Seagrass (Posidonia oceanica) vertical growth as an early indicator of fish farm- derived stress. Estuarine, Coastal and Shelf Science, 67, 475-483. [ Links ]
Moran, K. L. & Bjorndal, K. A. (2005). Simulated green turtle grazing affects structure and productivity of seagrass pastures. Marine Ecology Progress Series, 305, 235-247. [ Links ]
Moran, K. L. & Bjorndal, K. A. (2007). Simulated green turtle grazing affects nutrient composition of the seagrass Thalassia testudinum. Marine Biology, 150, 1083-1092. [ Links ]
Mortimer, J. A. (1981). The feeding ecology of the West Caribbean green turtle (Chelonia mydas) in Nicara- gua. Biotropica, 13, 49-58. [ Links ]
Pakulski, J. D. & Benner, R. (1992). An improved method for the hydrolysis and MBTH analysis of dissolved and particulate carbohydrates in seawater. Marine Chemistry, 40, 143-160. [ Links ]
Perez, D., Guevara, M., & Bone, D. (2006) Temporal variation of biomass and productivity of Thalassia testudinum (Hydrocharitacea) in Venezuela, Southern Caribbean. Revista de Biología Tropical, 54(2), 329-339. [ Links ]
Randall, J. E. (1965). Grazing effect on seagrasses by herbivorous reef fishes in the West Indies. Ecology, 46, 255-260. [ Links ]
Thayer, G. W., Bjorndal, K. A., Ogden, J., Williams, S., & Zieman, J. (1984) Role of larger herbivores in sea- grass communities. Estuaries, 7, 351-376. [ Links ]
Valentine, J. F. & Duffy, J. E. (2006). The central role of grazing in seagrass ecology. In T. Larkum, R. J. Orth, & C. M. Duarte (Eds.), Seagrass: Biology, ecology and their conservation (pp. 463-501). Dordrecht: Springer. [ Links ]
Valentine, J. F., Heck, K. L., Busby, J., & Webb, D. (1997). Experimental evidence that herbivory can increase shoot density in a subtropical turtlegrass (Thalassia testudinum) meadow. Oecologia, 112, 193-200. [ Links ]
Williams, S. L. (1988). Thalassia testudinum productivity and grazing by green turtles in a highly disturbed seagrass bed. Marine Biology, 98, 447-455. [ Links ]
Zieman, J. C. (1974). Methods for the study of the growth and production of turtle grass, Thalassia testudinum Konig. Aquaculture, 4, 139-143. [ Links ]
Zieman, J. C., Iverson, R. L., & Ogden, J. C. (1984). Herbivory effects on Thalassia testudinum leaf growth and nitrogen content. Marine Ecology Progress Series, 15, 151-158. [ Links ]
1. School of Natural Sciences and Mathematics, The Richard Stockton College of New Jersey, Galloway, New Jersey 08205 USA; elizabeth.lacey@stockton.edu 2. Department of Biological Sciences, Florida International University, Miami, Florida 33199 USA; colladol@fiu.edu, jim.fourqurean@fiu.edu
Received 17-XII-2013. Corrected 26-VI-2014. Accepted 21-VII-2014.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}