The relationship between physical and biological habitat conditions and hermatypic coral recruits abundance within insular reefs (Colombian Caribbean)
Katherine Bernal-Sotelo1* & Alberto Acosta1
*Dirección para correspondencia
Abstract Little evidence exists on the dependence between the presence and abundance of juvenile hermatypic corals and the conditions of their habitats, despite that juveniles contribute with the understanding of the community structure and its reproductive success. To assess this, the abundance of nine species of juvenile corals was correlated with eight macro-habitat (location of ]]>
Agaricia tenuifolia ]]>
Porites astreoides presented four moderate dependencies; ocean reefs between 2-16m depths, a high frequency of juveniles on horizontal substrates, exposed to light, non-sedimented and occupied by competitors.
Siderastrea siderea displayed three moderate dependences for juveniles in cryptic zones, inclined substrate and devoid of competitors. A. lamarcki, Leptoseris cucullata and A. agaricites presented two moderate dependences; these species share high abundance of juveniles in habitats with no sediment, exposed to light and occupied by competitors (except A. agaricites). ]]>
P. porites, Favia fragum and Montastraea cavernosa species had a moderate dependence with high incidence of juveniles in ocean reefs and microhabitats exposed to light. For the nine species, results indicate that the presence (colonization), abundance and survival of juveniles, depend on certain species-specific particularities of the habitat. However, the juveniles show high tolerance and plasticity to a range of habitat variables, given their independence and low dependence observed in over 50% of the variables assessed.
Key words: recruitment, coral recruits, habitat, insular reefs, Caribbean, Colombia, Agaricia, Porites, Siderastrea, Leptoseris, Favia,
Montastraea. Resumen Existe poca evidencia sobre la dependencia entre la abundancia de juveniles de corales ]]>
Agaricia ]]>
mostró las dependencias más robustas, siendo significativas para juveniles presentes frecuentemente en arrecifes continentales, sustrato horizontal expuesto a luz, con competidores. Los juveniles se asociaron con moderado a alto sedimento muy fino acumulado en el fondo. Porites astreoides presentó cuatro dependencias; alta frecuencia en sustrato expuesto a luz, horizontal, sin sedimento, con competidores y en arrecifes oceánicos entre 2-16m. Siderastrea siderea ]]>
A. lamarcki, Leptoseris cucullata, A. agaricites, P. porites, Favia fragum y Montastraea cavernosa mostraron el menor número de dependencias, compartiendo alta frecuencia en ]]>
Palabras clave: reclutamiento, corales juveniles, hábitat, arrecifes insulares, Caribe, ]]>
Agaricia, Porites, Siderastrea, Leptoseris, Favia, Montastraea.
Hermatypic coral ]]>
et al. 2010). The ecological and evolutionary significance is that recruitment is a key process within the reef’s successional cycle, since it determines the structure of the coral community, the renewal and continuity of local populations, favoring an increase of genetic variability, which in turn implies an adaptive advantage to climate change (Porter & ]]>
Recruitment measurable in situ by the presence of juveniles between 4mm and 2-4cm (depending on species) is considered the result of the synergy of several processes: partial and total mortality, that results in the quantity or coverage of reproductive adults, the total number of gametes and larvae released through sexual reproduction (reproductive output), the dispersal and survival of larvae in the water column, larval macro and micro habitat ]]>
et al. 2003, Sale et al. 2010). The process of metamorphosis to form polyps, asexual reproduction for juvenile growth, resistance to the physical environment conditions (Baird et al. 2003), and competition for resources such as substrate and light (McCook et al. ]]>
There are several possible ]]>
et al. 2006): 1- settlement and thus recruitment is an entirely random spatial process (not larval selection), in which the physical and biological habitat do not determine the pattern of recruitment (independent); 2- larvae of each species settle on preferential reef habitats according to their resource requirements, and habitat factors do not affect the distribution of recruits; 3- larvae select suitable settlement habitat, and habitat factors cause post-settlement mortality to produce the ]]>
Vermeij et al. (2006) have reported that in the case of environments providing conditions of high stress (drastic changes in salinity, sedimentation, light and competition) larvae do not present specific habitat selection; therefore, settlement becomes opportunistic and random. Contrastingly, coral larvae when ]]>
et al. 2004). There is no consensus on which of these hypotheses correctly explain the distribution and abundance of juvenile corals in a reef; it is a complex process, where multiple habitat variables interact, at different temporal and spatial scales, with larval settlement behavior. What is evident is that the fitness of any adult coral depends on the conditions of the habitat where the juvenile ]]>
Globally, we know that there are key factors of macro and micro-habitat that affect the survival of juvenile coral. At a macro-scale, for example, are the type and geographic location of the reef, depth and the degree of environmental degradation (Richmond 1997). At micro-scale are the availability and complexity of the substrate (Ruiz-Zárate et al. 2000), the type ]]>
et al. 2001). At micro-scale, the characteristics of the settlement substrate, is one of the factors that determine habitat preference and the pattern of juvenile survival (Ruiz-Zárate et al. 2000, Sale et al. 2010). The ]]>
et al. 2000, Baird et al. 2003). There is evidence that suggests that most larvae prefer roughened substrates, such as rocks and dead coral, as they offer countless micro-spaces with binding potential (Smith 1997); however, larvae species such as the Manicina ]]>
prefer attachment onto encrusting red algae because they facilitate settlement, metamorphosis and survival in conditions of high sedimentation (Ruiz-Zárate et al. 2000). Factors such as vulnerability to foraging (sea urchins, fish) and the ability to compete with macroalgae and other sessile organisms for space are involved in juvenile ]]>
et al. 2000, McCook et al. 2001). It has been shown that some algae not only compete for space and light with adult corals, but also with juveniles (overgrowth and suffocating), especially under conditions of nutrient enrichment, high sedimentation and low herbivores density (Birrell et al. 2008, Elmhirst et al. 2009). ]]>
The type and amount of sediment accumulated in the microhabitat is considered as the most significant factor determining larval and juvenile survival worldwide (Edmunds et al. 2004, Fabricius 2005). The proximity to river mouths causes an increase in the amount of particulate organic and inorganic matter entering the reef, causing nutrification, turbidity and sedimentation. ]]>
et al. 2004, Hughes et al. 2007); a decrease of the larvae’s sensory ability to select microhabitat, and a decline in the rate of juvenile survival and diversity (Gardner et al. 2003). The negative effects of sediment runoff entering the reef have been reported by ]]>
Fabricius (2005) in even the early reproductive stages (gametogenesis, fertilization and embryogenesis) as well as the late stages (larval survival, settlement, metamorphosis, juvenile growth and survival). An increase in reef regime alterations since the early seventies has caused a modification in the ]]>
et al. 2003). Juvenile coral communities are enduring a “community shift” (Aronso et al. 2004, Hughes et al. 2007), caused by the dominance of brooding species such as the Agaricia spp. and Porites spp., which replaced spawning species, such as
Montastraea annularis and Siderastrea siderea (Porter & Tougas 2001, Green et al. 2008). Although it is clear that the habitat factors mentioned above are important for both adult and juvenile corals, the differential effects of each factor in the post-settlement survival by species that determine the frequency, ]]>
et al. 2008). Consequently, it is unknown whether the recruitment and survival of coral species in the Caribbean, in fact, depends on certain physical and biological habitat factors or if it is entirely random (Mundy & Babcoock 2000). Investigating the consequences (juveniles of the same species present in the same macro or micro-habitat=frequency) could help to infer the causal agents that are ]]>
The contribution of this study is to demonstrate whether the frequency of juvenile hermatypic corals depends on certain physical factors of the habitat on a macro-scale (location of the reef on the shelf and depth) and on a micro-scale (type and angle of the substrate, exposure to light, texture and amount ]]>
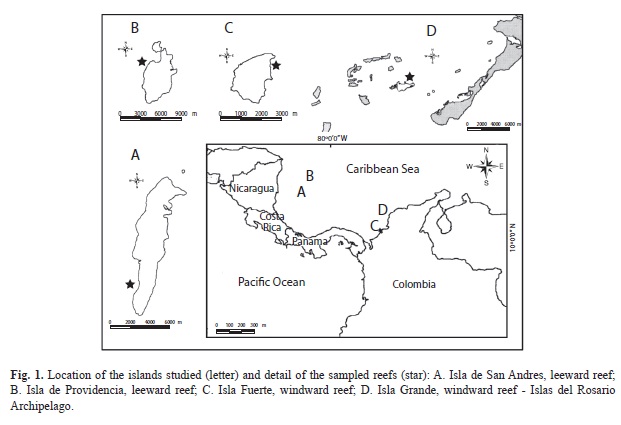
Materials and methods Study area: The study was conducted in four insular reefs in the Colombian Caribbean (Fig. 1), two on the continental shelf (Isla Fuerte and Isla Grande) subject to different levels of ]]>
Continental shelf reefs: Isla Grande is part of the Islas del Rosario archipelago, located Southwest of Cartagena in the Parque Nacional Natural Corales del Rosario and San Bernardo. It is influenced by the Magdalena River (average flow=7 149.53m3/s, Garay 2001), which flows into the Canal del ]]>
3/s (Garay 2001) and is the main affluent of the archipelago. The profile of the windward terraces (10º10’21’’ N - 75º42’36’’ W) is characterized by a reef crest less than 10m wide and a slope with coral cover up to 25m. The reefs feature a predominance of M. annularis, Agaricia tenuifolia and Porites astreoides, along with deteriorated coral patches of
Acropora palmata and extensive coverage of macroalgae such as Dictyota spp. and Halimeda spp. (Alvarado et al. 1989). Isla Fuerte is part of the Bajo Bushnell and Bajo Burbujas reef complex (Díaz
et al. 1996); it is influenced by the following rivers: the Sinú (average flow=290.8m3/s, Garay 2001), the Atrato (average flow=2 366.13m3/s, Garay 2001) and the Magdalena (7 149.53m3/s, Garay 2001). The windward zone (9º23’15.6’’ N - 76º10’10.1’’ W) has a terrace formed by a fringing reef up to a depth of 20m, followed by a sandy bottom, with a coral zonation typical of the Caribbean (Díaz et al. 2000), ]]>
Dictyota spp.) and the accumulation of extremely fine sediment (siltclay). The annual loss rate by species or genus in continental reefs, caused by sedimentation and pollution has been estimated at 0.4-2.2% (Acosta & Martínez 2006). Ocean platform reefs: Sampling was conducted west of San Andres and Providencia, at coordinates ]]>
Dendrogyra cylindrus, Diploria labyrinthiformis, D. strigosa, Colpophyllia natans and
Montastraea (Díaz et al. 1995). The latter terrace ends abruptly at 20-22m with an inclined coral covered slope and sediment to 30-35m (Díaz et al. 1995, Acosta & Martínez 2006). Leeward in Providence has a broad lagoon terrace covered with sediment and sea grass areas. The shallow ]]>
Acropora palmata, Millepora complanata, zoanthids, and fleshy and coralline algae. The fore-reef terrace (12-15m) has an assortment of corals, predominantly of the Diploria spp., M. annularis, Dichocoenia stokesi, P. astreoides species, as well as octocorals, while in deeper waters, the coverage of Montastraea cavernosa,
M. annularis, D. labyrinthiformis and C. natans increases, as well as the presence of sponges and octocorals (Díaz et al. 2000). Coral coverage on the slope decreases rapidly (Acosta & Martínez 2006).
Sampling: A line was traced from the deepest point (maximum 30m), where the reef ends or the sandy bottom begins, to the coast, in areas of the most thriving coral reefs (richness, coral coverage). Perpendicular to this line and at increasing depths of 2m, 12 quadrants of 1m2 were measured, six on each side of the line and 1m from each other (following Vidal et al. 2005); this procedure was repeated along the entire length of the reef depth gradient. The transects were grouped by depth in four categories ]]>
et al. 2011): 1- Shallow (2, 4, 6 and 8m), 2- Medium (10, 12, 14 and 16m), 3- Deep (18, 20, 22 and 24m) and 4- Very deep (>26m), the latter only found in oceanic reefs, totaling 600m2 sampled. Juveniles were defined as colonies of less than 4cm in diameter for large species (diameter>15cm) such as Montastraea spp., Diploria ]]>
S. sidereal (Bak & Engel 1979, Richmond & Hunter 1990, Dueñas et al. 2010); and 2cm in diameter for small species (diameter<15cm), such as P. astreoides and Favia fragum. Taxonomic identification was done according to the Dueñas et al. (2010) guide. ]]>
As the number of species and juveniles per quadrant was recorded, the physical and biological characteristics of the habitat in which each one of the juveniles was found were described as follows: 1- Location of the reef on the platform; 2- Range of depth; 3- Type of substrate colonized by the juvenile; 4- Substrate inclination on which the juvenile was found; 5- Juvenile’s exposure to light; 6- Texture of the sediment accumulated on the substrate around the juvenile; 7- Amount of accumulated sediment on ]]>
table 1 for further details on classification and measurement criteria for each variable.
The 3 000 juveniles and 45 species ]]>
α2)(pq)/n, where Zα=1.96 at 95% confidence, p=percentage of the community expected to be sampled (75%), q=1-p, n=sample size (total number of individuals per species). These species were Agaricia agaricites,
Leptoseris cucullata, P. astreoides, S. siderea, F. fragum, Scolymia spp., A. lamarcki, M. cavernosa, A. fragilis, P. porites, S. radians and
A. tenuifolia. These 12 species were used to analyze the pattern of dependencies between juveniles and the characteristics of their habitat, by employing the chi-square (X2) test for contingency tables (CT). The Yates continuity correction was used on 2x2 CT. When at maximum 20% of the table cells had an expected frequency under five, we applied Fisher’s exact test. However, the proper application of the X2 test to Scolymia spp., A. fragilis and S. ]]>
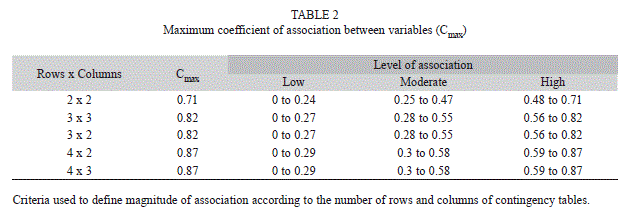
was impossible, as more than 20% of the cells in the contingency tables continuously resulted in frequencies equal to zero; consequently, they were excluded from the analysis. For the nine remaining species, in cases where H0 was rejected (independence of variables), we calculated the contingency coefficient (C) to determine the extent of dependence. This coefficient ranged between zero and a maximum value of association (Cmaximum) which corresponds to the number of categories of the variables that were evaluated. Values ]]>
maximum as a high association (Table 2). Not all combinations of variables were tested; when crossing certain categories of independent variables in the CT some had frequencies of zero and expected frequencies under five, this precluded the application of the test. Subsequently, a multiple correspondence analysis (MCA) was used to reduce variable information to two dimensions on a Cartesian plane (Díaz 2002), graphically ]]>
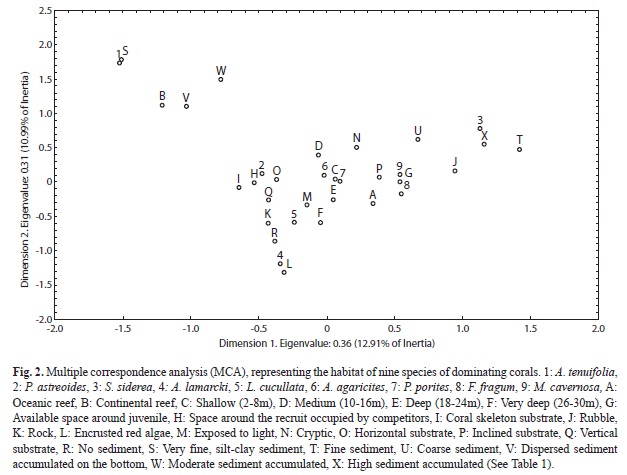
Results The X2 test (p<0.05) and the MCA (Fig. 2) evidenced the ]]>
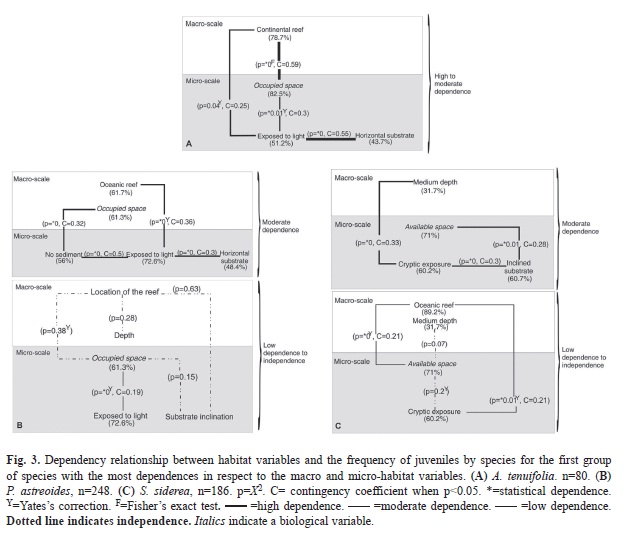
A. tenuifolia. Only this species showed a significantly high dependence (CT 2x2, C=0.59; Fig. 3A), between the frequency of juveniles and habitats located in continental reefs and juveniles surrounded by competitors (78.7% continental reefs and 82.5% with macroalgae and corals). High dependence was also found among juveniles on substrates exposed to light and those on horizontal substrates (CT 3x2, C=0.55; Fig. 3A). ]]>
Fig. 3A). Graphically, the MCA illustrates how A. tenuifolia settled apart from other species, beside the habitat with moderate to high silt-clay sediment in continental ]]>
Table 3). The frequency of juvenile P. astreoides, moderately depended (CT 2x2, 3x2 y 4x2, C=0.3-0.5; Fig. 3B), on oceanic reefs (61.7%) with substrates exposed to light (72.6%), ]]>
It was also established that the frequency of juvenile S. siderea ]]>
Fig. 3C). Graphically (by the MCA) it was established that the higher frequency of juveniles of the species was on rubble-type substrates (78.5%), cryptic (60.2%), of coarse and fine sediment (47.3% and 39.8% respectively), accumulated in amounts greater than 1cm (69.9%). See Appendix 1 ]]>
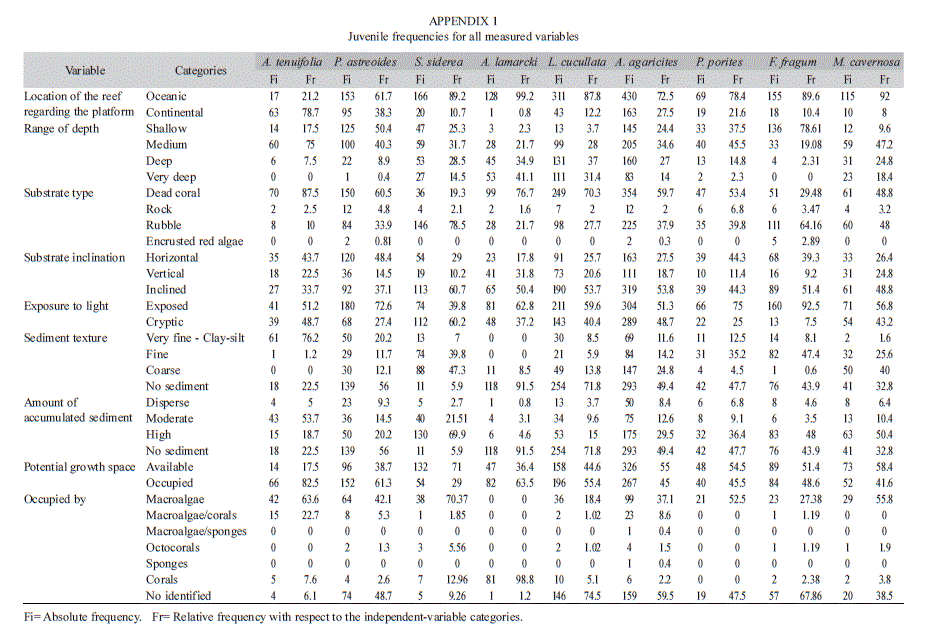
The highest frequency of A. lamarcki juveniles depended on substrates occupied by competitors (63.5%), with no sediment accumulated (91.5%) on substrate exposed to light (62.8%). The magnitude of the dependence between variables ranged from low to moderate (CT 2x2 and 4x2; C=0.24-0.34;
Fig. 4A). Graphically it was confirmed that the factor closest to A. lamarcki and which determined the frequency of its juveniles, was the absence of sediment on the substrate since, through the MCA, the species was located apart from the other species. However, statistical independence (X2, p>0.05) was proved between light exposure and inclination of the substrate, and between the inclination of the substrate and space for growth around the juveniles (
Fig.4A). Abundance was lowest in continental reefs (0.8%) and at depths under 10m (2.3%). Like P. astreoides, juveniles of L. cucullata and A. agaricites showed that despite being in a variety of ]]>
Fig. 4B and 4C) and the MCA’s two-dimensional plane, the strongest dependence (moderate) was presented by juveniles in substrate devoid of accumulated sediment and substrate exposed to light; with 71.8% and 59.6% of L. cucullata juveniles under these conditions (CT 4x2, C=0.43; Fig. 4B). Additionally, moderate dependence ]]>
Fig. 4B) for L. cucullata between the frequency of juveniles on non-sedimented substrates (71.8%) and substrates occupied by competitors (55.4%). A. agaricites also showed moderate dependence (CT 4x2, C=0.42; Fig. 4C) between non-sedimented substrates (49.4%) and light availability (51.3%), and those without ]]>
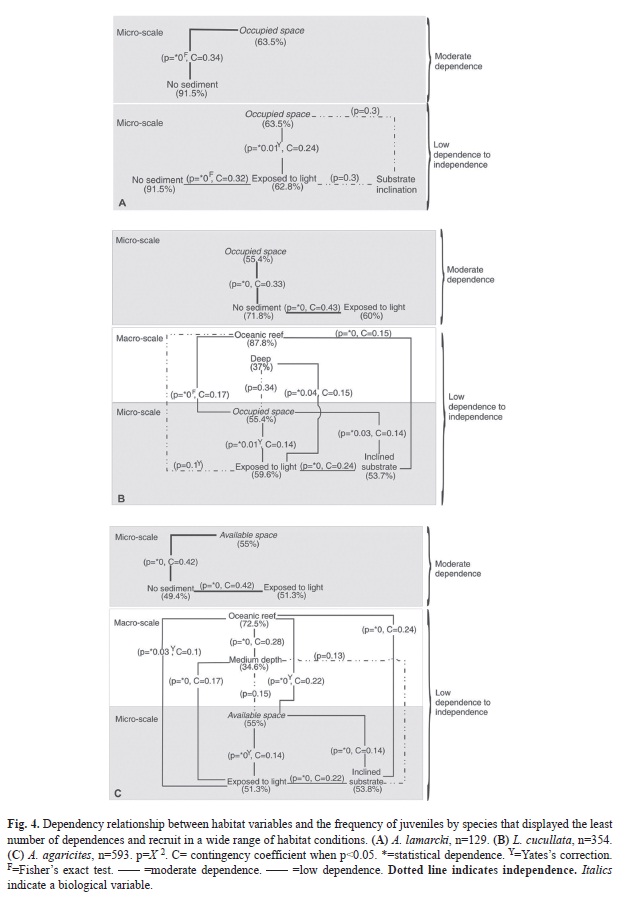
The juveniles of P. porites showed dependence on oceanic environments (78.4%) and microhabitats without interaction or the presence of other sessile organisms (54.5%), of moderate dependence (CT 2x2; C=0.42; Fig. 4D). Independence between its ]]>
2, p>0.05) was proven for this species. As for F. fragum, we found that the abundance of juveniles depended on ocean reefs (89.6%) and substrates exposed to light (92.5%) of moderate intensity (CT 2x2; C=0.32; Fig. 4E). The species was abundant at depths under 16m and on rubble, ]]>
Appendix 1). However, statistical independence was confirmed between the location of the reef and the potential space available for juvenile growth, and the latter with light exposure (X2, p>0.05). The juveniles of M. cavernosa ]]>
showed moderate dependence (CT 4x2; C=0.35; Fig. 4F) on substrates exposed to light (56.8%), at depths of 10-16m (47.2%). In addition, the independence between light exposure and the potential space to grow was established, and the latter with substrate inclination (X2, p>0.05). While juveniles of species such as ]]>
S. siderea, were most abundant in environments with accumulated sediment, the strongest relationships (higher frequency of juveniles) for A. lamarcki, P. astreoides, L. cucullata and A. agaricites juveniles, were present in micro-habitats exposed to light and without sediment accumulation around juveniles; this evidenced that ]]>
L. cucullata, A. agaricites, P. porites, F. fragum and M. cavernosa were grouped within the MCA plane, amid many categories of the different physical and biological variables ]]>
Discussion ]]>
These results suggest that the frequency, abundance and survival of juvenile hermatypic corals in insular reefs of the Colombian Caribbean are affected by physical and biological features of their environment and that they depend on certain peculiarities of the habitat, which are species-specific. According to the characteristics of relationships and the number of dependences of moderate to high magnitude, the following two groups of species were recognized: 1- species with the highest number of dependences (3-4) with regard to the macro and micro-habitat variables ]]>
A. tenuifolia,
P. astreoides and
S. siderea) and species that displayed the least number of dependences (1-2) and recruit in a wide range of habitat conditions (
A. lamarcki,
L. cucullata,
A. agaricites,
P. porites,
F.
fragum and
M. cavernosa).
The mechanism that describes the dependence of juveniles on certain habitat categories and variables could be simply explained as the balance between larval input where the larvae perform or not a selection within the spectrum of habitat conditions and provide a new individual to the population, and the output to the system generated by the differential mortality of ]]>
et al. 2000, Harrington et al. 2004), 2. No larval ]]>
et al. 2006) and strong selection pressure in all but one of the habitat categories, which would explain the dependence identified. In turn, larval substrate selection depends on habitat characteristics (quality, quantity and resource availability) and conditions of the environment, which have not been quantified for any species, in the reefs assessed. Meanwhile, the number of larvae reaching a system depends on population size, number of reproductive individuals, reproductive effort (Alvarado & ]]>
et al. 2010). Also, the natural selection that occurs in larval stages, affects the settlement and metamorphosis (Mundy & Babcock 2000, Baird et al. 2003) and later post-settlement (Vermeij et al. 2006, Victor 2008). The complexity of the variables involved and the lack of understanding regarding the early life stages ]]>
The independence between habitat variables and the abundance of juveniles could be explained by the combination of the following: 1. Absence of larval selection or opportunistic behavior at the time of settlement (Vermeij et al. 2006), concurrently with low natural selection of juveniles within the ]]>
et al. 2003) in a system with high spatial heterogeneity and where resources and conditions are distributed in patches or follow a gradient.
Some of the traits most relevant to the higher survival rate of coral species are: high phenotypic plasticity, which confers them tolerance to various conditions of the environment (Miller & Barimo 2001, Green et al. 2008) and resistance to changes in habitat conditions (Lirman & Manzello 2009), efficiency in the removal of accumulated sediment (mostly by ]]>
et al. 2008); rapid colonization of empty substrates as a result of a high investment in sexual reproduction and periodic release of planulae resistant to limiting resources (Smith 1997, Kramer 2003). These strategies are likely to provide them predominance as an adult and even as a youth in many regions of the world (Smith 1997). The graphic association (on the MCA) of species of the second group with various habitat factors confirms ]]>
The robust pattern of high and moderate dependences displayed by A. tenuifolia could be accounted by their life history traits and their ability to withstand extreme conditions. Its vertical growth could provide an advantage by ]]>
et al. 2004). The combination of turbidity, high sedimentation rate and increased nutrient concentrations, typical of continental reefs in this region (Aronso et al. 2004, Gardner
et al. 2003, Acosta & Martínez 2006) especially of Isla Fuerte, favors the proliferation of macroalgae and decreases substrate availability for coral recruitment. Because it is a brooder and has a high level of reproductive effort and tolerance to adverse mediums, A. tenuifolia has been able to rapidly colonize empty substrates and increase their recruitment (Hughes & Jackson 1985). As a result, the presence of A. tenuifolia ]]>
Porites sp. in recent decades (Aronso et al. 2004). The low number of A. tenuifolia juveniles in ocean reefs, where there is adequate numbers of adult reproductive individuals and where sedimentation is lower, may suggest that the species is a better competitor in stressful conditions, such as in areas with high silt-clay sediment (Miller & Barimo 2001), or that this ocean reef ]]>
P. astreoides’ light requirements are consistent with the high frequency of juveniles in horizontal locations, exposed to light, in clear water ocean reefs and recruitment at shallow depths (Vidal et al. 2005). Cofforth (1985) and Miller et ]]>
. (2000) assert that P. astreoides invest high levels of energy in protection and competition, producing mucus and presenting a gregarious behavior, which reduces possible cases of inter-specific competition and aggression by algae, giving them a higher rate of survival (Green et al. 2008). This strategy and the colonization of substrates devoid of sediment accumulation or with strong hydrodynamics, produces a lower energy expenditure for cleaning, energy that it can invest in competition with macroalgae such as ]]>
sp. and sp., which occupy 12-15% of the substrate (Alvarado & Acosta 2009). Although Miller & Barimo (2001) and Green Halimedaet al. (2008) state that P. astreoides is tolerant to different physical and biological habitats, juveniles appear to show specificity to habitat factors. The favorable ]]>
S. sidereal juveniles and cryptic locations, inclined substrates and the lack of competitors could reveal the strategy employed by the larvae to colonize crevices, clefts and holes. This occurs when the larvae are faced with limitations of settlement substrate (e.g. due to high algal cover). This type of environment is abundant in ocean reefs at depths between 10-16m, where the reef has higher rugosity and presents a 3D structure. The amount of coarse and fine sediment accumulated in holes (greater than 1cm) presents a disadvantage for juveniles in this type ]]>
S. siderea boasts a resistance to sedimentation, which probably gives it an advantage to settle in holes or rubble, as shown in the MCA in this study. Another disadvantage caused by the crevices is the reduction of direct light necessary for juvenile growth, which in turn is advantageous since it repels macroalgae and prevents the expenditure of energy on competition. Barrios (2000) stresses the high sensitivity of
S. siderea juvenile to the aggression of other sessile organisms. However, although it has been proposed that S. siderea is an indicator species for pollution and sedimentation (Guzmán & Jiménez 1992), juveniles were scarce in continental environments where these factors are present, this may be due to low roughness and 3D structure of the system to recruit, a small number of adults and hence low reproductive effort of this spawner species, as Alvarado & Acosta (2009) found with lower number of gamets for M. annularis ]]>
For its part, the dependence of juvenile A. lamarcki, L. cucullata and A. agaricites on a non-sedimented bottom, exposed to light and ocean reef habitats, ]]>
A. lamarcki larvae, this species has high survival rate (Hughes 1988), higher than even A. agaricites and L. cucullata, the dominant species of juveniles ]]>
A. lamarcki, the monopolization of substrate indeep oceanic reefs the interaction with corals of the same species. Among all the species, A. agaricites ]]>
et al. 2005), in agreement with Chiappone & Sullivan (1996) and the study by Edmunds (2000) in Florida and St. John – Virgin Islands, respectively. Juvenile
P. porites, F. fragum and M. cavernosa had a dependence on ocean reefs, a microhabitat exposed to light and unoccupied by competitors. The susceptibility of P. porites to competition with macroalgae may explain its requirement to colonize spaces without competitors and with direct light. River & Edmunds (2001) assert that the growth rate of P. porites decreases up to 80% when surrounded by macroalgae. Contrary to Lewis (1974), who found that most larvae colonize cryptic sites, displaying a gregarious behavior by ]]>
Although the variables assessed here are those considered as critical to the recruitment of hermatypic corals in Caribbean reefs, exploring and evaluating other biotic (foraging) and abiotic factors of the habitat is essential, given the dependence of the species on specific factors. Moreover, the diverse causes of the observed patterns of dependency and the synergy between ]]>
]]>
The density of juveniles found is not considered sufficient to replace the dying adults in the reefs studied; this makes juvenile richness and density very low when compared to that of adult corals (Acosta et al. 2011). As the fitness of coral species decreases because of recruitment failure, so does the resilience of the reef, its recovery and populations’ viability (Sale et al. 2010). Seven of the most active species in recruitment were ]]>
S. siderea, which as Carlon (2002) proposes, may be an indication of a potential change in dominance of the coral assemblage from building species for shortlived brooding species (juvenile
A. agaricites,
L. cucullata and
P. astreoides). The low recruitment and the poor recovery of populations in the system also facilitate a community shift in continental reefs ]]>
et al. 2011).
The reality of Caribbean reefs is the great decline caused by anthropic disturbance and inland waters that increase the direct input of sediment and nutrients. This according to Alvarado & Acosta (2009) creates stress, population mortality, reduction in colony size, low number of breeding ]]>
et al. 2006), recruitment in sub-optimal locations and postsettlement mortality (Fabricius 2005, Vermeij et al. 2006, Victor 2008) and consequently, low recruitment (Acosta et al. 2011). To achieve ]]>
In conclusion, the ]]>
S. sidereal and F. fragum on rubble), on inclined areas, exposed to direct light (except S. siderea and species of the Agaricia genus, cryptic), where F. ]]>
Porites genus are highly dependent on light (abundant in shallow and medium zones). This indicates the importance of clear water with low turbidity (low concentration of dissolved and particulate organic matter). Few species used rocky or encrusted red algae substrates; this reflects their low number and availability. Similarly, juveniles seem to prefer substrates where sediment does not accumulate, particularly A. lamarcki and L. cucullata, in contrast to what ]]>
S. siderea, A. tenuifolia and M. cavernosa that tolerate high sediment loads. Juveniles recruit and survive on substrates occupied, primarily by macroalgae; this is evident for A. tenuifolia and in a lesser degree for A. lamarcki and P. astreoides, while S. siderea displays the lowest number of interactions. Most species use a ]]>
Acknowledgments
To the Academic Vice-rectory of the Pontificia Universidad Javeriana that financed this project (0417; 4803), to Flavia Cárdenas, Mónica Sepúlveda, Claudia Villamil and Andrés Vidal for the field work that yielded the data used in this study, as well as to Margarita Ordoñez and Miguel Pinzón for their statistical advice. To Gypsy Espanol for the translation into the English version. ]]>
References Acosta, A. & S. Martínez. 2006. Continental and oceanic coral reefs in the Colombian Caribbean after a decade of degradation. Proc. 10th Int. ]]>
Coral Reef Symp. 2: 1926-1930. [ Links ] Acosta, A., L.F. Dueñas & V Pizarro. 2011. Review on hard coral recruitment (Cnidaria: Scleractinia) in Colombia. Univ. Sci. 16: 200-218. (también disponible en línea: www.javeriana.edu.co/universitas_scientiarum/articulos/online.html). [ Links ] Alvarado, E., G. Pinilla, T. León & A. Cortes. 1989. Plan de Manejo - Parque Nacional Corales del Rosario. Inderena, Universidad Jorge Tadeo Lozano, Bogotá, Colombia. [ Links ] Alvarado, E. & A. Acosta. 2009. Fertilidad y fecundidad de Montastraea annularis en un arrecife degradado. Bol. Invest. Mar. Cost. 38: 91-108. [ Links ] Aronso, R.B., I.G. Macintyre, C.M. Wapnick & M.W. O’Neill. 2004. Phase shifts alternative states and the unprecedented convergence of two reef systems. Ecology 85: 1876-1891. [ Links ] Baird, A.H., R.C. Babcock & C.P. Mundy. 2003. Habitat selection by larvae influences the depth distribution of six common coral species. Mar. Ecol. Progr. 252: 289-293. [ Links ] Bak, R.P. & M.S. Engel. 1979. Distribution, abundance and survival or juvenile hermatypic corals (Scleractinia) and the importance of life history strategies in the parent coral community. Mar. Biol. 54: 341-352. [ Links ] Barrios, L.M. 2000. Evaluación de las principales condiciones de deterioro de los corales pétreos en el Caribe colombiano. Tesis de Posgrado en Biología – Línea Biología Marina, INVEMAR, Universidad Nacional de Colombia, Santa Marta, Colombia. [ Links ] Birrell, C.L., L.J. McCook, B.L. Willis & G.A. Díaz-Pulido. 2008. Effects of benthic algae on the replenishment of corals and the implications for the resilience of coral reefs. Oceanogr. Mar. Biol. 46: 25-63. [ Links ] Carlon, D.B. 2002. Production and supply of larvae as determinants of zonation in a brooding tropical coral. J. Exp. Mar. Biol. Ecol. 268: 33-46. [ Links ] Chiappone, M. & K.M. Sullivan. 1996. Distribution, abundance and species composition of juvenile scleractinian corals in the Florida reef tract. Bull. Mar. Sci. 58: 555-569. [ Links ] Cofforth, M.A. 1985. Mucous sheet formation on poritiid corals: effects of altered salinity and sedimentation. Proc. 5th Int. Coral Reef Symp. 4: 165-170. [ Links ] Díaz, J.M., J. Garzón-Ferreira & S. Zea. 1995. Los arrecifes coralinos de la Isla de San Andrés, Colombia: estado actual y perspectivas para su conservación. Acad. Col. Cienc. Exact. Fís. Nat., Colección Jorge Álvarez Lleras 7, Bogotá, Colombia. [ Links ] Díaz, J.M., J.A. Sánchez & G. Díaz-Pulido. 1996. Geomorfología y formaciones arrecifales recientes de Isla Fuerte y Bajo Bushnell, Plataforma continental del Caribe colombiano. Bol. Invest. Mar. Cost. 25: 87-105. [ Links ] Díaz, J.M., L.M. Barrios, M.H. Cendales, J. Garzón-Ferreira, J. Geister, M. López-Victoria, G.H. Ospina, F. Parra-Velandia, J. Pinzón, B. Vargas-Ángel, F.A. Zapata & S. Zea. 2000. Áreas Coralinas de Colombia. INVEMAR, Ser. Pub. Esp. 5, Santa Marta, Colombia. [ Links ] Díaz, L.G. 2002. Estadística multivariada: inferencias y métodos. Universidad Nacional de Colombia, Panamericana, Bogotá, Colombia. [ Links ] Dornelas, M., S.R. Connolly & T.P. Hughes. 2006. Coral reef diversity refutes the neutral theory of biodiversity. Nature 440: 80-82. [ Links ] Dueñas, L.F., J. Montenegro, A. Acosta, F. Cárdenas, M. Sepúlveda, A. Vidal & C. Villamil. 2010. Guide to Scleractinian coral recruits from the Caribbean. Pontificia Universidad Javeriana, INVEMAR Serie de Documentos Generales No. 42, XPRESS, Bogotá, Colombia. [ Links ] Edmunds, P.J. 2000. Patterns in the distribution of juvenile corals and coral reef community structure in St. John, US Virgin Islands. Mar. Ecol. Prog. Ser. 202: 113-124. [ Links ] Edmunds, P.J. 2004. Juvenile coral population dynamics track rising seawater temperature on a Caribbean reef. Mar. Ecol. Prog. Ser. 269: 111-119. [ Links ] Edmunds, P.J., J.F. Bruno & D.B. Carlon. 2004. Effects of depth and microhabitat on growth and survivorship of juvenile corals in the Florida Keys. Mar. Ecol. Progr. 278: 115-124. [ Links ] Elmhirst, T., S.R. Connolly & T.P. Hughes. 2009. Connectivity, regime shifts and the resilience of coral reefs. Coral Reefs 28: 949-957. [ Links ] Fabricius, K.E. 2005. Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Mar. Pollut. Bull. 50: 125-146. [ Links ]
Garay, J. 2001. Calidad ambiental marina en Colombia año 2000, p. 97-114. In INVEMAR (eds.). Informe del estado de los ambientes marinos y costeros en Colombia: 2000. Ser. Doc. Gral. 7, Santa Marta, Colombia. [ Links ] Gardner, T.A., I.M. Cote, F.A. Gill, A. Grant & A.R. Watkinson. 2003. Long-term region-wide declines in Caribbean corals. Science 301: 958-960. [ Links ] Garzón-Ferreira, J., M.C. Reyes-Nivia & A. Rodríguez-Ramírez. 2002. Manual de métodos del SIMAC - Sistema Nacional de monitoreo de Arrecifes Coralinos en Colombia. INVEMAR, Ministerio del Medio Ambiente, Santa Marta, Colombia. [ Links ] Green, D.H., P.J. Edmunds & R.C. Carpenter. 2008. Increasing relative abundance of Porites astreoides on Caribbean reefs mediated by an overall decline in coral cover. Mar. Ecol. Progr. 359: 1-10. [ Links ] Guzmán, H.M. & C.E. Jiménez. 1992. Contamination or coral reefs by heavy metals along the Caribbean coast of Central America (Costa Rica and Panamá). Mar. Pollut. Bull. 24: 554-561. [ Links ] Harrington, L., K. Fabricius, G. De’ath & A. Negri. 2004. Recognition and selection of settlement substrata determine post-settlement survival in corals. Ecology 85: 3428-3437. [ Links ] Hughes, T.P. 1985. Life histories and population dynamics of early successional corals. Proc. 5th Int. Coral Reef Symp. 4: 101-106. [ Links ] Hughes, T.P. 1988. Long-term dynamics of coral populations: contrasting reproductive modes. Proc. 6th Int. Coral Reef Symp. 2: 721-725. [ Links ] Hughes, T.P. & J.B.C. Jackson. 1985. Population dynamics and life histories of foliaceous corals. Ecol. Monogr. 55: 141-166. [ Links ] Hughes, T.P. & J.E. Tanner. 2000. Recruitment failure, life histories and long term decline of Caribbean corals. Ecology 81: 2250-2263. [ Links ] Hughes, T.P., M.J. Rodriguez, D.R. Bellwood, D. Ceccarelli, O. Hoegh-Guldberg, L. McCook, N. Moltschaniwskyj, M.S. Pratchett, R.S. Steneck & B. Willis. 2007. Phase shifts, herbivory and the resilience of coral reefs to climate change. Curr. Biol. 17: 360-365. [ Links ] Huitric, M. & M. McField. 2001. Effects of multiple disturbances on hard coral recruits in Glovers Reef Atolls lagoon, Belize. Beijer Discussion Paper 138: 1-20. [ Links ] Kramer, P.A. 2003. Synthesis of coral reef health indicators for the Western Atlantic: results of the AGRRA program (1997-2000). Atoll. Res. Bull. 496: 1-57. [ Links ] Lewis, J.B. 1974. The settlement behavior of planulae larvae of the hermatypic coral Favia fragum (Esper). J. Exp. Mar. Biol. Ecol. 15: 165-172. [ Links ] Lirman, D. & D. Manzello. 2009. Patterns of resistance and resilience of the stress-tolerant coral Siderastrea radians (Pallas) to sub-optimal salinity and sediment burial. J. Exp. Mar. Biol. Ecol. 369: 72-77. [ Links ] Martínez, S. & A. Acosta. 2005. Cambio temporal en la estructura de la comunidad coralina del área de Santa Marta - Parque nacional Natural Tayrona (Caribe Colombiano). Bol. Invest. Mar. Cost. 34: 161-191. [ Links ] McCook, L.J., J. Jompa & G. Díaz-Pulido. 2001. Competition between corals and algae on coral reefs: a review of evidence and mechanisms. Coral Reefs 19: 400-417. [ Links ] Miller, M.W. & J. Barimo. 2001. Assessment of juvenile coral populations at two reef restoration sites in the Florida Keys National Marine Sanctuary: indicators of success? Bull. Mar. Sci. 69: 395-405. [ Links ] Miller, M.W., E. Weil & A.M. Szmant. 2000. Coral recruitment and juvenile mortality as structuring factors for reef benthic communities in Biscayne National Park, USA. Coral Reefs 19: 115-123. [ Links ] Montenegro, J. & A. Acosta. 2008. Havistat 2008 V.1.0. ©Programa para inferir uso y preferencia de hábitat. Unesis, Departamento de Biología, Facultad de Ciencias, Pontificia Universidad Javeriana, Bogotá, Colombia. [ Links ] Montenegro, J. & A. Acosta. 2010. Habitat preference of Zoantharia genera depends on host sponge morphology. Universitas Scientiarum 15: 110-121. [ Links ] Mundy, C.N & R.C. Babcock. 1998. Role of light intensity and spectral quality in coral settlement: implications for depth-dependent settlement? J. Exp. Mar. Biol. Ecol. 223: 235-255. [ Links ] Mundy, C.N & R.C. Babcock. 2000. Are vertical distribution patterns of scleractinian corals maintained by pre- or post-settlement processes? a case study of three contrasting species. Mar. Ecol. Progr. 198: 109-119. [ Links ] Pita, S. 1996. Determinación del tamaño muestral. Cad. Aten. Primaria 3: 138-141. [ Links ] Porter, J.W. & J.I. Tougas. 2001. Reef ecosystem: threats to their biodiversity, p. 73-95. In S.A. Levin (ed.). Encyclopedia of Biodiversity. Academic, San Diego, USA. [ Links ] Richmond, R.H. 1997. Reproduction and recruitment in corals: critical links in the persistence of reefs, p. 175- 197. In C. Birkeland (ed.). Life and death of Coral Reefs. Chapman & Hall, New York, USA. [ Links ] Richmond, R.H. & C.L. Hunter. 1990. Reproduction and recruitment of corals: comparison among the Caribbean, the Tropical Pacific, and the Red sea. Mar. Ecol. Progr. 60: 185-203. [ Links ] River, G.F. & P.J. Edmunds. 2001. Mechanisms of interactions between macroalgae and scleractinian on a coral reef in Jamaica. J. Exp. Mar. Biol. Ecol. 261: 159-172. [ Links ] Ruiz-Zárate, M.A., J. Espinoza-Avalos, J.P. Carricart-Ganivet & D. Fragososo. 2000. Relationships between Manicina areolata (Cnidaria: Scleractinia), Thalassia testudinum (Anthophyta) and Neogoniolithon sp. (Rhodophyta). Mar. Ecol. Progr. 206: 135-146. [ Links ] Sale, P.F., H. Van Lavieren, M.C.A. Lagman, J. Atema, M. Butler, C. Fauvelot, J.D. Hogan, G.P. Jones, K.C. Lindeman, C.B. Paris, R. Steneck & H.L. Stewart. 2010. Preserving Reef Connectivity: A Handbook for Marine Protected Area Managers. Connectivity Working Group, Coral Reef Targeted Research & Capacity Building for Management Program, UNUINWEH, Melbourne, Australia. [ Links ] Smith, S.R. 1997. Patterns of coral settlement, recruitment and juvenile mortality with depth at Conch Ref., Florida. Proc. 8th Int. Coral Reef Symp. 2: 1197-1202. [ Links ] Vermeij, M.J.A., N.D. Fogarty & M.W. Miller. 2006. Pelagic conditions affect larval behavior, survival, and settlement patterns in the Caribbean coral Montastraea faveolata. Mar. Ecol. Progr. 310: 119-128. [ Links ] Victor, S. 2008. Stability of reef framework and post settlement mortality as the structuring factor for recovery of Malakal Bay Reef, Palau, Micronesia: 25 years after a severe COTS outbreak. Estuar. Coast. Shelf Sci. 77: 175-180. [ Links ] Vidal, A., A. Acosta & C. Villamil. 2005. Composición y densidad de corales juveniles en dos arrecifes profundosde San Andrés Isla, Caribe colombiano. Bol. Invest. Mar. Cost. 34: 211-225. [ Links ]
*Correspondencia:
Katherine Bernal-Sotelo: Unidad de Ecología y Sistemática, Departamento de Biología, Pontificia Universidad Javeriana, Bogotá, Colombia. johanna.bernal@javeriana.edu.co ]]>
Alberto Acosta: Unidad de Ecología y Sistemática, Departamento de Biología, Pontificia Universidad Javeriana, Bogotá, Colombia. laacosta@javeriana.edu.co 1. Unidad de Ecología y Sistemática, Departamento de Biología, Pontificia Universidad Javeriana, Bogotá, Colombia; johanna.bernal@javeriana.edu.co, laacosta@javeriana.edu.com
Received 11-VII-2011. Corrected 20-II-2012. Accepted 19-III-2012.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}