Javier Rodríguez-Barrios1, Rodulfo Ospina-Tórres2 & Rodrigo Turizo-Correa3
1. Grupo de Biodiversidad y Ecología Aplicada, Instituto de Investigaciones Tropicales, Universidad del Magdalena, Carrera 32 No 22-08, Santa Marta, Colombia. PBX: 4217940; jarodriguezba@gmail.comDirección para correspondencia
Abstract
Functional feeding groups of macroinvertebrates in Gaira river, Colombia. Tropical rivers are frequently described on their biodiversity but few studies have considered the ecological value of this richness in their food webs. We determined the trophic structure of aquatic macroinvertebrate communities (expressed in the richness and abundance of taxa and biomass proportions of different functional feeding groups) at the level of the river, stretch and microhabitats (functional units - UFs). We evaluated the spatial and temporal variation of these descriptors during wet and dry events, and selected three sites associated with different altitudinal belts. We reported 109 taxa, with 11 167 individuals who contributed 107.11g of biomass. Density of macroinvertebrates was favored with increasing height, and biomass showed the opposite pattern (K-W=10.1, d.f.=1, p<0.05), due to the addition of large crustaceans (Macrobrachium), and the taxa diversity was higher in the middle stretch of the river (H’=3.16). The Gaira stream runs through a mid-sized river basin, for this reason we found mainly bedrock (epilithon=50.5%), gravel and sand (43.7%). The functional unit with more habitat and food resources that contains a higher abundance of leaf litter macroinvertebrates was foliage followed by epilithon, fine sediment and gravel-sand (K-W=25.3, d.f.=3, p<0.05). The biomass values of these organisms were higher in leaves followed by gravel-sands, epilithon and sediment (K-W=15.3, d.f.=3, p<0.05). Autochthonous biomass input by different functional feeding groups can be considered very low, but they define the functionality of the stream, being represented almost exclusively by shredders (Macrobrachium, 73%), present only in the lower reaches, followed by shredder Leptonema with 15%, located mostly in the upper reaches and predatory stoneflies of the genus Anacroneuria to 6.56%, which dominated in the middle stretch of stream. Excluding Macrobrachium from the analysis, there was dominance of Anacroneuria in the lower reaches. Between rainfall and drought events, biomass of functional feeding groups was higher in rain (W=10.1, d.f.=1, p<0.05), favoring the growth of decapods, but the abundance was much higher during drought events. Rev. Biol. Trop. 59 (4): 1537-1552. Epub 2011 December 01.
Resumen
Palabras clave: macroinvertebrados acuáticos, grupos funcionales alimentarios, unidades funcionales, río tropical de montaña, biomasa.
La inmensa diversidad de la biota de los ríos tropicales, dificulta obtener un conocimiento completo sobre la estructura de sus comunidades, siendo la diversidad biológica uno de los temas centrales de las investigaciones, junto con la descripción y distribución de sus especies (Jackson & Sweeney 1995). La mayoría de estudios sobre la biota de ríos de Sur América, se ha utilizado como herramienta para el diagnóstico de la calidad ambiental del agua, sin tener en cuenta el papel trófico y otros aspectos ecológicos y funcionales de estas especies (Greathouse & Pringle 2006). Por lo tanto se debe generar información que integre elementos de la estructura de las comunidades y la función de estos organismos en los ecosistemas fluviales. De esta manera se podrán obtener modelos ecológicos que permitan hacer mejores generalizaciones conceptuales sobre la estructura y funcionamiento de estas corrientes tropicales (Tomanova et al. 2006).
El objetivo del presente trabajo consistió en evaluar la estructura trófica a nivel de GFAs de la comunidad de macroinvertebrados presentes en tres localidades dentro del gradiente altitudinal del río Gaira y durante un año de muestreos, con el fin de determinar: (1) la distribución de los GFAs a escala de microhábitats o unidades funcionales (UF) y entre tramos ubicados en un gradiente altitudinal del río y (2) el efecto de los periodos de lluvia y sequía sobre la composición de los GFAs. Estos elementos espacio-temporales se consideran modeladores de la estructura y función en ecosistemas torrentosos como los de la SNSM.
Materiales y métodos
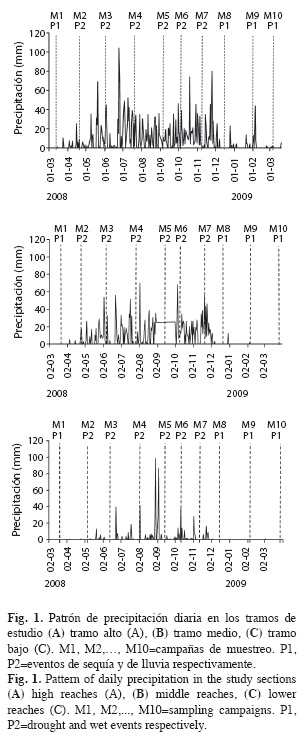
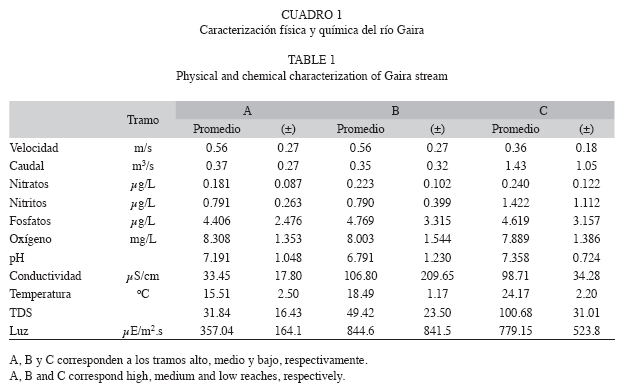
Se realizaron 10 muestreos (M1-M10), entre marzo 2008 y abril 2009. Este periodo abarcó eventos de sequía y de lluvias (P1 y P2 respectivamente) (Fig. 1A-C). El primer muestreo (M1) se realizó entre el 20 y 23 de marzo, el segundo (M2) entre el 27 de abril y siete de mayo, el tercero (M3) entre el 20 y 17 de junio, el cuarto (M4) fue durante el 26 de julio y el dos de agosto, el quinto (M5) entre el 13 y el 15 de septiembre, el sexto (M6) durante el tres y el seis de octubre, el séptimo (M7) del 11 al 14 de noviembre, el octavo (M8) entre el 13 y 16 de diciembre, el noveno (M9) durante el 31 de enero y el tres de febrero y el décimo muestreo entre el 27 de marzo y primero de abril. El río Gaira, presenta valores altos de velocidad de la corriente, bajos niveles de nutrientes, el pH es ligeramente básico y la temperatura muestra una variación importante entre los tramos evaluados (Cuadro 1).
Muestreo: Se seleccionaron tres tramos de 200m, de acuerdo a las consideraciones espaciales de Frisell et al. (1986), con el criterio de suficiente heterogeneidad microgeomorfológica y el sistema de rápidos y remansos desarrollado por Dunne & Leopold (1978), que consiste en tomar un tramo de la quebrada que presente un conjunto de rápidos y remansos (mínimo tres de cada uno) por cada tramo. La heterogeneidad de microhábitatas en estos tramos fue subdividida en cuatro tipos de UFs, con distintos componentes bióticos y abióticos, que representan a microambientes en los cuales se desarrollan los GFAs. Las UFs seleccionadas fueron: hojarasca (HOJ), epiliton (EPI), gravas y arena (GRA) y sedimento fino (SED) (modificado de Gutiérrez 2006).
]]>En cada muestreo realizado y con la ayuda de una red surber con ojo de malla de 200μm (área de muestreo=0.09m2), se recolectaron los macroinvertebrados acuáticos presentes en las distintas UF, de acuerdo a los procedimientos modificados de Merrit & Cummins (1996). En cada tramo y muestreo se recolectó un total de tres muestras por UF. El material extraído por la red, fue almacenado en bolsas de polietileno y almacenado en alcohol etílico al 90%. En el laboratorio, las muestras fueron revisadas bajo el estereoscopio y cuando fue necesario se realizaron montajes para observación al microscopio. El material biológico fue determinado hasta el nivel taxonómico más específico posible (en la mayoría de los casos hasta género), utilizando las claves taxonómicas y descripciones de Lopretto (1995), Merritt & Cummins (1996) y Fernández & Domínguez. (2001). Debido a la limitación taxonómica para poder llegar hasta el nivel de especies con larvas, se acogieron las propuestas de Dolédec et al. (2000) y Gayraud et al. (2003), quienes demuestran que la resolución hasta nivel de especie no es indispensable en estudios de diversidad funcional, considerando al género como el máximo nivel necesario. Todo el material identificado fue corroborado y reposa en la colección de referencia del laboratorio de invertebrados acuáticos de la Universidad Nacional de Colombia.
Los GFA considerados en este trabajo fueron: colectores-filtradores (C-F), colectoresrecolectores (C-R), raspadores (R), depredadores (D) y fragmentadores (T). La asignación de los GFA de macroinvertebrados se realizó de acuerdo con la literatura especializada para táxones neotropicales (Tomanova et al. 2006, Chara et al. 2010) y con la ayuda de expertos. La biomasa se estimó mediante regresiones de longitud-peso de cada taxon identificado, utilizando las regresiones propuestas por Benke et al. (1999) y Cressa (1999).
Resultados
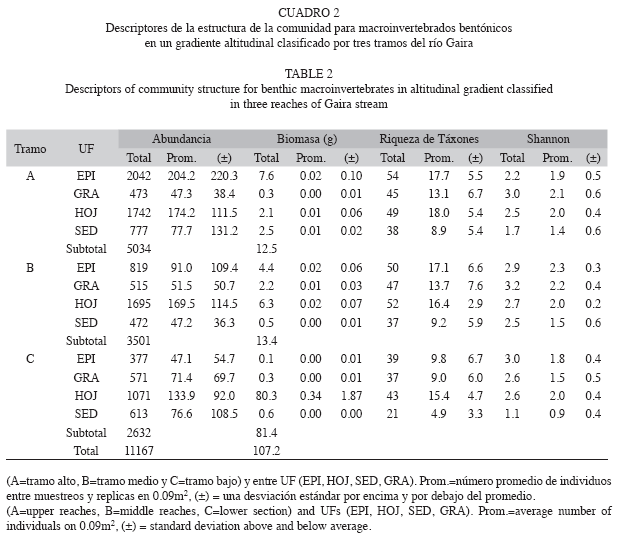
Patrón de abundancia, biomasa y diversidad: Durante las diez campañas de muestreo realizadas en los tres tramos, se recolectó un total de 11 167 individuos, distribuidos en 12 ordenes, 52 familias y 109 táxones de macroinvertebrados, los cuales aportaron una biomasa total de 107.11g de peso seco/32.4m2 de área muestreada con la red surber (3.306g/m2) (Cuadro 2). A nivel de tramos, la mayor abundancia se presentó en el tramo alto (A), seguido del tramo medio (B) y tramo bajo (C) con 5 034, 3 501 y 2 632 individuos respectivamente (Cuadro 2). El tramo con mayor aporte de biomasa fue el tramo bajo, seguido de los tramos medio y alto con 81.4, 13.4 y 12.48g respectivamente (K-W=10.1, gl=1, p=0.0015).
Un patrón similar al presentado por la abundancia, se encontró en la riqueza de táxones, por cuanto el tramo alto presentó el mayor valor con 81 táxones, seguido de B y C con 75 y 63 táxones, pudiéndose determinar una relación directa entre la altura y la riqueza de táxones. Los valores promedio de diversidad de táxones, expresados mediante el índice de Shanon-Wiener (H’), fueron mayores en el tramo medio con 3.16, seguidos de los tramos bajo y alto con 2.65 y 2.58 respectivamente. A nivel de las UFs, HOJ presentó la mayor abundancia con 4 508 individuos (promedio=450.8±196.4), seguida de EPI con 3 237 individuos (promedio=693.6±1348.8), SED con 1 862 individuos (promedio=354.6±483.2) y GRA con 1 559 individuos (promedio=524.1±980.8). Esta variación en el patrón de abundancias no fue significativa (K-W=1.37, gl=2, p=0.5).
Los valores de biomasa total también fueron mayores en HOJ con 88.7g (promedio=8.9±9.5), seguida de EPI con 12.5g (promedio=1.3±0.8), GRA con 5.1g (promedio=0.5±0.4) y SED con 0.9g (promedio= 0.09±0.07). La variación en la abundancia y biomasa por tramos es reportada en el Cuadro 2. La UF con una mayor diversidad de Shannon fue GRA, seguida de HOJ, EPI y SED con 3.3, 2.9, 2.8 y 2.01 respectivamente (Cuadro 2).
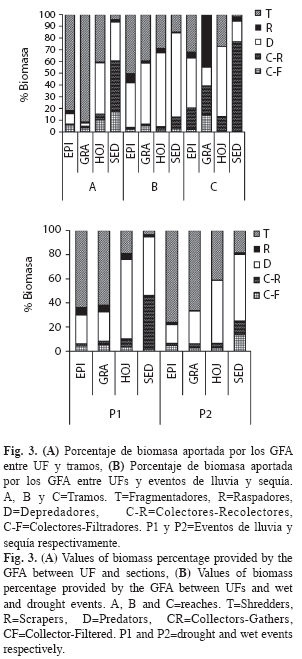
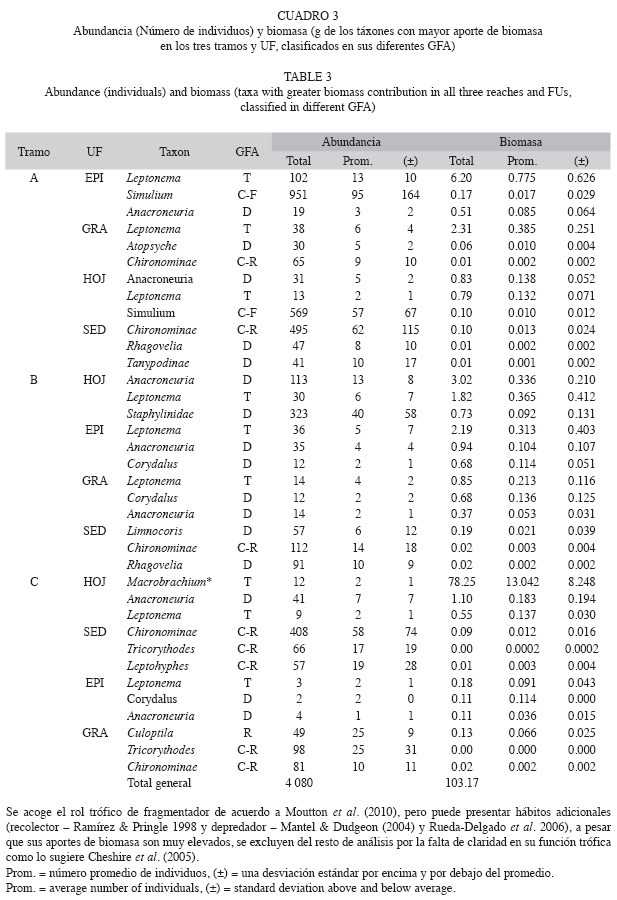
Variación espacial de los GFA: A escala de río, los colectores-recolectores dominaron en riqueza y abundancia con 29 táxones y 5 228 individuos (H’=1.95), seguidos por los depredadores con 28 táxones y 1 426 individuos (H’=2.29), los raspadores con 23 táxones y 1 510 individuos (H’=1.99), los fragmentadores con 12 táxones y 543 individuos (H’=1.64) y los colectores-filtradores con 9 táxones y 2 443 individuos (H’=0.73).
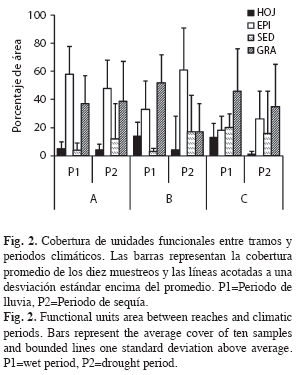
A escala de UFs, se presenta una variabilidad en el patrón de distribución de los diferentes GFA (W=14.5, gl=1, p=0.0023) (Fig. 3A). Los fragmentadores y raspadores tienden a concentrarse en sustratos rocosos y arenosos definidos por EPI y GRA. Los depredadores se distribuyen en todos los microhábitats, principalmente en HOJ y SED y con bajas proporciones a mayor altura. Los colectores suelen encontrar mayor oferta de recursos en el sustrato fino determinado por SED (Fig. 3A), posiblemente la alta frecuencia de los depredadores en este sustrato se deba a la oferta de colectores y su preferencia por este tipo de habitat.
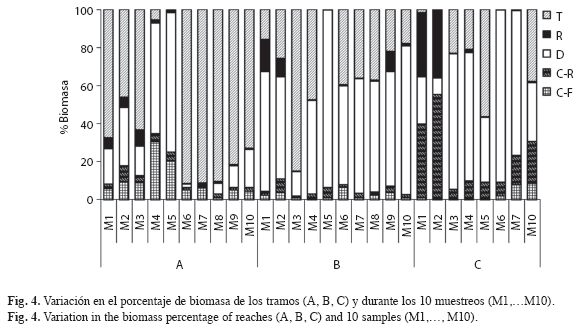
En el tramo alto se presenta una tendencia sucesional marcada, con el incremento en la proporción de depredadores y raspadores al inicio y final de la sequía (M8 a M10 y M1 a M5); ésta, disminuye de manera importante con la entrada de las lluvias en M5, favoreciendo el aumento y posterior dominancia de los fragmentadores entre M6 y M10 (Fig. 4). En el tramo medio, la dominancia de depredadores seguida de los fragmentadores es más homogénea, relacionado con bajas proporciones de raspadores y colectores. En el tramo bajo, el periodo seco es más prolongado y con menores niveles de precipitación en la época de lluvias (Fig. 1). En este tramo hay una mayor proporción de colectores-recolectores y raspadores durante los muestreos de periodo seco (M7 a M10 y M1 a M2). Los fragmentadores dominan desde el inicio de las lluvias, representado por M3 a M6 (Fig. 4), debido a que en este periodo se presentó un aumento en la frecuencia de grandes pupas de Leptonema, ubicadas debajo de rocas.
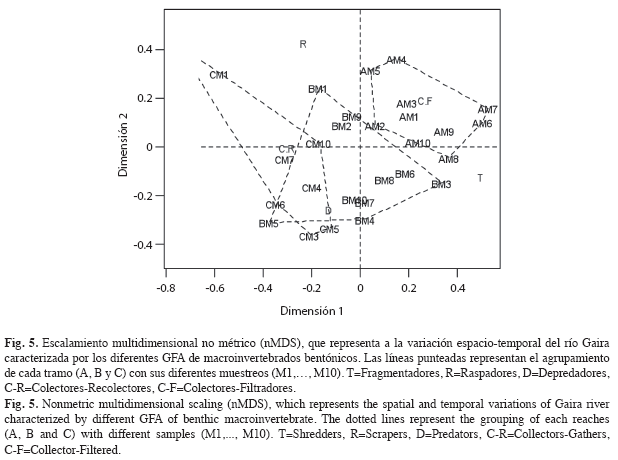
La caracterización de los tramos a lo largo de los diez muestreos, se determina por altos valores de biomasa para los fragmentadores y colectores-filtradores en el tramo alto, mientras que los tramos medio y bajo fueron caracterizados por la biomasa de depredadores y colectores-recolectores. Los raspadores incrementaron de manera importante en el muestreo 1 (P1) del tramo medio (Fig. 4). El análisis de PERMANOVA indica que los tramos evaluados presentan una diferencia en cuanto al rol trófico de sus GFA representativos (Seudo F=7.53, g.l. 2, p=0.005) (Fig. 5), reafirmando la importancia de los fragmentadores en los tramos de cabecera (A y B) y de los depredadores y colectores-recolectores en el tramo bajo.
Discusión
Cobertura de las UFs: La heterogeneidad espacial, descrita por las UFs, se encuentra influenciada por la condición geomorfológica del río, principalmente por ser una cuenca de tamaño mediano (máximo orden=6), de alta pendiente y bajo potencial de almacenamiento de materiales, condiciones que promueven a una dominancia importante de sustratos rocosos (EPI), principalmente en los tramos altos y medio (Allan & Castillo 2007).
Patrón de abundancia, biomasa y diversidad: Los datos reportados para el río Gaira, demuestran que a pesar de su corto recorrido, es un ecosistema fluvial de alta diversidad y abundancia faunística, con 11 167 individuos agrupados en 109 táxones, cifras que superan a varios estudios de la región tropical, así como a estudios para la región andina realizados por Chara et al. (2010), quienes encontraron 2 019 individuos, 52 familias y 71 táxones presentes en ocho quebradas de la cuenca del río la Vieja. Gutiérrez (2006), en una quebrada de la Sabana de Bogotá y a escala de tramo, reporta 82 táxones. Para la región amazónica colombiana, Rueda-Delgado et al. (2006), reportan en HOJ 5 456 individuos en 39 táxones. Para el sector medio y bajo del río, en el presente estudio se citan 3 501 y 2 632 individuos, superando al único reporte precedente, en esos sectores por Rueda (2005), quien encontró 1 076 y 1 950 individuos en 27 familias. En ríos del trópico de Vietnam se ha logrado encontrar una mayor riqueza de táxones a la del presente estudio, con reportes de hasta 230 géneros y 91 familias (Nguyen et al. 2001, Hoang & Bae 2006). Para ríos de Brasil, Landeiro et al. (2010) encontraron una cifra inferior, representada por 8 365 individuos distribuidos en 35 táxones.
La mayor biomasa fue dominada por Macrobrachium, Leptonema, Anacroneuria y Coridalus (Cuadro 3), los cuales caracterizan a los tramos bajo, alto y medio respectivamente por su aporte en peso. Ramírez & Pringle (1998), reportan que para algunos ríos de Puerto Rico la mayor abundancia y biomasa está representada por insectos efemerópteros (Thraulodes) con el 10% y entre el grupo de los "no-insectos" Macrobrachium representa el 40% del aporte total de biomasa.
Variación espacial de los GFA: Los colectores-recolectores y los depredadores dominaron en la riqueza general del río. En términos de abundancia se encuentran los colectores-recolectores, seguidos de los colectores-filtradores, pero la biomasa fue dominada por los fragmentadores y depredadores. Autores como Chara et al. (2010) plantean que aunque los colectores sean abundantes pero de poco tamaño, su importancia ecológica no supera a otros grupos con menos abundancia pero que aportan mayor biomasa, como los fragmentadores y los depredadores.
Otros trabajos en ríos tropicales reportan una baja proporción de fragmentadores (Ramírez & Pringle 1998, Greathouse & Pringle 2006, Gutiérrez 2006, Rueda-Delgado et al. 2006, Wantzen & Wagner 2006), atribuyendo este patrón a que su función es reemplazada por raspadores y recolectores. Cheshire et al. (2005) atribuyen la baja proporción de fragmentadores en estudios como los anteriores, al mal uso de la clasificación funcional de los macroinvertebrados, pues son trabajos en los que se suele utilizar la clasificación de GFA de zonas templadas como la de Merritt & Cummis (1996), siendo que el rol trófico y las historias de vida de estos organismos difiere en varios casos para nuestras regiones. En este sentido se sugiere la revisión de contenidos estomacales o la utilización de estudios que hayan realizado este procedimiento en el trópico. Quizá el primer reporte válido para el trópico en el que se pondera poco a los aportes de biomasa por los fragmentadores, lo desarrollaron Tomanova et al. (2006) en ríos de Bolivia y Chara et al. (2010) para Colombia. Estos últimos destacan a Leptonema como un fragmentador importante por su gran tamaño. Otros autores resaltan la dominancia de los colectores debido a que su caracterización se desarrolla con base en lasabundancias y no en la biomasa (eg. Hoang & Bae 2006 en el Trópico de Vietnam, Dudgeon 1994, en Hong Kong y Dudgeon & Bretschko 1996, en Nueva Guinea), remitiéndose a referencias de GFA de regiones templadas.
]]> La biomasa de GFA que se encontró en las diferentes UF, demuestra que no se puede hacer una generalización a escala de río o de tramo, como lo sugiere Vannote et al. (1980), pues se subestima el patrón de distribución y preferencia de microhábitats por los GFA, asumiendo que quizá no sea posible generalizar el rol trófico de los macroinvertebrados a meso y macroescalas.GRA y SED a pesar de presentar altas densidades de colectores-recolectores, presentan baja biomasa de organismos, posiblemente por su baja estabilidad mecánica al efecto de la corriente, como lo sugiere Gutiérrez (2006) en SED. Autores como Huryn & Wallace (1987) y Ramírez & Pringle (1998), sugieren una menor oferta de hábitat para comunidades de macroinvertebrados en sustratos de remansos (pools), en donde dominan los sedimentos. Para ríos tropicales, Pringle (1996) sugiere que estos tipos de hábitat soportan mayor presión de los peces, camarones y macroinvertebrados depredadores. Se sabe además que los SED y GRA son sustratos que ofrecen una buena oferta de refugios para sostener la mayor densidad de macroinvertebrados (Arunachalam et al. 1991).
Los resultados demuestran que el río Gaira, por su ubicación y tamaño, mantiene una dependencia funcional alóctona, en donde la biomasa viva que aportan estos GFs es mínima, comparada al suministro orgánico proveniente de la vegetación ribereña, que ingresa, se almacena y se moviliza a lo largo del río. Es así como las comunidades de macroinvertebrados, en respuesta a su ambiente, incrementan su rol trófico en sus proporciones de fragmentadores, para aprovechar el suministro particulado alóctono típico de estos ríos de cabecera (Allan & Castillo 2007, Vannote et al. 1980). Los depredadores, juegan un importante papel como el segundo rol trófico en importancia, en relación con la alta abundancia de grupos como los colectores y fragmentadores, estos últimos tienden a disminuir sus tamaños con el incremento de depredadores. En este sentido, se puede afirmar que el tramo alto es caracterizado por macroinvertebrados acuáticos fragmentadores, el tramo medio por el papel trófico de los depredadores y fragmentadores y el tramo bajo por una mayor complejidad funcional representada por los depredadores, colectoresrecolectores y raspadores.
Agradecimientos
Los autores agradecen a la División de Investigación de la Universidad Nacional de Colombia (DIB) por la financiación del proyecto "Descriptores funcionales de un sistema fluvial de montaña" con el cual se pudo llevar a cabo este trabajo. También se agradece a los grupos GIBEA y GILIN de la Universidad del Magdalena, así como a Gaspar Polanco, por su apoyo en el desarrollo exitoso del presente trabajo, tanto en la fase de campo como en la de laboratorio. Agradecimientos especieales al señor Micky Weber por permitirnos hacer el estudio de manera exitosa en la Hacienda la Victoria.
Allan, J.D. & M.M. Castillo. 2007. Stream ecology, estructure and function of running waters. Chatman & Hall, Nueva York, Nueva York, EEUU. [ Links ]
Arunachalam, M., K.C. Madhusoodan, J. Vijverberg, K. Kortmulder & H. Suriynarayanans. 1991. Substrate selection and seasonal vitiation in densities of invertebrates in stream pools of a tropical river. Hydrobiologia 213: 141-141. [ Links ]
Benke, A.C., A.D. Huryn, L.A. Smock & J.B. Wallace. 1999. Length-mass relationships for freshwater macroinvertebrates in North America with particular reference to the southeasten United States. J. N. Am. Benthol. Soc. 18: 308-343. [ Links ]
Britto, E.F., T.P. Moulton, M.L. Souza & S.E. Bunn. 2006. Stable isotope analysis indicates microalgae as the predominant food source of fauna in a coastal forest stream of Southeast Brazil. Aust. J. Ecol. 31: 623-633. [ Links ]
Chara, A.M., J.D. Chara, M. Zúñiga, G.X. Pedraza & L.P. Giraldo. 2010. Clasificación trófica de insectos acuáticos en ocho quebradas protegidas de la ecorregión cafetera colombiana. U. Scient. 15: 27-36. [ Links ]
Cheshire, K., L. Boyero & R.G. Pearson. 2005. Food webs in tropical Australian streams: shredders are not scarce. Freshwater Biol. 50: 748-769. [ Links ]
Colwell, R.K. & D.C. Lees. 2000. The mid-domain effect: geometric constraints on the geography of species richness. Trends Ecol. Evol. 15: 70-76. [ Links ]
Cressa, C. 1999. Dry mass estimates of some tropical aquatic insects. Rev. Biol. Trop. 47: 133-141. [ Links ]
Dolédec, S., J.M. Olivier & B. Statzner. 2000. Accurate description of the abundance of taxa and their biological traits in stream invertebrate communities: effects of taxonomic and spatial resolution. Arch. Hydrobiol. 148: 25-43. [ Links ]
Drake, J.A. 1984. Species aggregation: the influence of detritus in a benthic invertebrate community. Hydrobiologia 112: 109-115. [ Links ]
Dudgeon, D. 1994. The influence of riparian vegetation on macroinvertebrate community structure and functional organization in six New Guinea streams. Hydrobiologia 294: 65-85. [ Links ]
Dudgeon, D. & G. Bretschko. 1996. Allochthonous inputs in land-water interactions in seasonal streams: tropical Asia and temperate Europa, p. 161-179. In F. Schiemer & K.T. Boland (eds.). Perspectives in tropical limnology. Academic, Amsterdam, Holanda. [ Links ]
Dunne, T. & L. Leopold. 1978. Water in Environmental Planning. W.H. Freeman, San Francisco, California, EEUU. [ Links ]
Espinal, L.S. & E. Montenegro. 1963. Formaciones vegetales de Colombia. Memoria explicativa del mapa ecológico. Inst. Geogra. Agustín Codazzi, Bogotá, Colombia. [ Links ]
Fernández, H.R. & E. Domínguez. 2001. Guía para la determinación de artrópodos bentónicos sudamericanos. Universitaria de Tucumán, Universidad Nacional de Tucumán, Argentina. [ Links ]
Frayter, V., E. Jiménez, R. Pabón & V. Rivera. 2000. Plan de manejo integral de la cuenca hidrográfica del río Gaira. Tesis de Grado, Universidad del Magdalena, Santa Marta, Colombia. [ Links ]
Frisell, C.A., W.J. Liss, C.E. Warren & M.D. Hurley. 1986. A hierarchical framework for stream classification: viewing streams in a watershed context. Environ. Manag. 10: 199-214. [ Links ]
Gayraud, S., B. Statzner, P. Bady, A. Haybachp, F. Scholl, P. Usseglio-Polatera & M. Bacchi. 2003. Invertebrate traits for the biomonitoring of large European rivers: an initial assessment of alternative metrics. Freshwater Biol. 48: 1-20. [ Links ]
Greathouse, E.A. & C.M. Pringle. 2006. Does the river continuum concept apply on a tropical island? Longitudinal variation in a Puerto Rican stream. Can. J. Fish. Aquat. Sci. 63: 134-152. [ Links ]
Grubaugh, J.W., J.B. Wallace & E.S. Houston. 1996. Longitudinal changes of macroinvertebrate communities along an Appalachian stream continuum. Can. J. Fish. Aquat. Sci. 53: 896-909. [ Links ]
Gutiérrez, J.D. 2006. Caracterización del metabolismo y de la oferta de recursos de materia orgánica para la fauna de macroinvertebrados bentónicos en una quebrada de montaña de orden menor. Tesis de Doctorado, Universidad Nacional de Colombia, Bogotá, Colombia. [ Links ]
Hoang, D.H. & Y.J. Bae. 2006. Aquatic insect diversity in a tropical Vietnamese stream in comparison with that in a temperate Korean stream. Limnology 7:45-55. [ Links ]
Huryn, A.D. & J.B. Wallace. 1987. Local geomorphology as a determinant of macrofaunal production in a mountain stream. Ecology 68: 1932-1942. [ Links ]
Jackson, J.K. & B.W. Sweeney. 1995. Research in tropical streams and rivers: introduction to a series of papers. J. N. Am. Benthol. Soc. 14: 2-4. [ Links ]
Landeiro, V.L., N. Hamada, B.S. Godoy & A.S. Melo. 2010. Effects of litter patch area on macroinvertebrate assemblage structure and leaf breakdown in Central Amazonian streams. Hydrobiologia 649: 355-363. [ Links ]
Lau, D.C., K.M. Leung & D. Dudgeon. 2009. Are autochthonous foods more important than allochthonous resources to benthic consumers in tropical headwater streams? J. N. Am. Benthol. Soc. 28: 426-439. [ Links ]
Lopretto, E. 1995. Crustacea Eumalacostraca, p. 250-265. In C.E. Lopretto & G. Tell (eds.). Ecosistemas de aguas continentales: Metodologías para su estudio. Ediciones del Sur, La Plata, Argentina. [ Links ]
MacArthur, R.H. 1972. Geographical Ecology. Harper & Row, Nueva York, EEUU. [ Links ]
McCoy, E.D. 1990. The distribution of insects along elevational gradients. Oikos 58: 313-332. [ Links ]
Mackay, R.J. &. J. Kalf. 1969. Seasonal variation in standing crop and species diversity of insects communities in a small Quebec stream. Ecology 50: 101-109. [ Links ]
Mantel, S.K., M. Salas & D. Dudgeon. 2004. Foodweb structure in a tropical Asian forest stream. J. N. Am. Benthol. Soc. 23: 728-755. [ Links ]
Merritt, R.W. & K.W. Cummins. 1996. An introduction to the aquatic insects of North America. Kendall Hunt, Dubuque, Iowa, EEUU. [ Links ]
Minshall, G.W., K.W. Cummins, R.C. Petersen, D.A. Bruns & C.E. Cushing. 1985. Developments in stream ecosystem theory. Can. J. Fish. Aquat. Sci. 42: 1045-1055. [ Links ]
Moulton, T.P., S.A. Magalhaes-Fraga, E.F. Brito & F.A. Barbosa. 2010. Macroconsumers are more important than specialist macroinvertebrate shredders in leaf processing in urban forest streams of Rio de Janeiro, Brazil. Hydrobiologia 638: 55-66. [ Links ]
Nguyen, V.V., D.H. Hoang, T.K. Cao, X.Q. Nguyen, Y.J. Bae. 2001. Altitudinal distribution of aquatic insects from Tam Dao National Park in northern Vietnam, p. 123-133. In Y.J. Bae (ed.). The 21st century and aquatic entomology in East Asia, Jeonghaengsa, Seoul, Corea. [ Links ]
Pringle, C.M. 1996. Atyid shrimps (Decapoda: Atydae) influence the: spatial heterogenety of algal communities over different scales in tropical montane streams, Puerto Rico. Freshwater Biol. 35: 125-140. [ Links ]
R. Development Core Team. 2010. R: A language and environment for statistical computing 2.11.1. R Foundation for Statistical Computing, Vienna, Austria. (disponible en línea: http://www.R-project.org). [ Links ]
Rahbek, C. 1995. The elevational gradient of species richness: a uniform pattern? Ecography 18: 200-205. [ Links ]
Ramírez, A. & C.M. Pringle. 1998. Structure and production of a benthic insect assemblage in a neotropical stream. Rev. Biol. Trop. 17: 443-463. [ Links ]
Reice, S.R. 1980. The role of substratum in benthic macroinvertebrate microdistribution and litter decomposition in a woodland stream. Ecology 61: 580-590. [ Links ]
Rodríguez, J.A. 2006. Dinámica de la materia orgánica en la cabecera de una quebrada de montaña. Tesis de Maestría, Universidad Nacional de Colombia, Bogotá, Colombia. [ Links ]
Rodríguez-Barrios, J., R. Ospina-Torres, J.D. Gutiérrez & H. Ovalle. 2007. Densidad y biomasa de macroinvertebrados acuáticos derivantes en una quebrada tropical de montaña (Bogotá, Colombia). Caldasia 29: 397-412. [ Links ]
Rueda-Delgado, G. 2005. Lineamientos de un Programa de aseguramiento de la oferta hídrica del río Gaira a partir de la evaluación de la Integralidad biológica de la cuenca. DADMA. Universidad del Magdalena, Santa Marta, Colombia. [ Links ]
Rueda-Delgado, G., K.M. Wantzen & M. Beltrán. 2006. Leaf-litter decomposition in an Amazonian floodplain stream: effects of seasonal hydrological changes. J. N. Am. Benthol. Soc. 25: 233-249 [ Links ]
Sanders, N.J. 2002. Elevational gradients in ant species richness: area, geometry, and Rapoport’s rule. Ecography 25: 25-32. [ Links ]
]]>Silveira, M.L. & T.P. Moulton. 2000. Modelling the food web of a stream in Atlantic forest. Acta Limnol. Bras. 12: 63-71. [ Links ]
Stout, J. & J.H. Vandermeer. 1975. Comparison of species richness for stream· inhabiting insects in tropical and mid-latitude: streams. Am. Nat. 109: 263-280. [ Links ]
Tamaris-Turizo, C. & H.J. López. 2006. Aproximación a la zonificación climática de la cuenca del río Gaira. Inotrópica 3: 69-76. [ Links ]
Tamarís, C., R. Turizo & M. del C. Zúñiga. 2007. Distribución espacio-temporal y hábitos alimentarios de ninfas de Anacroneuria (Insecta: Plecoptera: Perlidae) en el río Gaira (Sierra Nevada de Santa Marta, Colombia). Caldasia 29: 375-385. [ Links ]
Tomanova, S., E. Goitia & J. Helešic. 2006. Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia 556: 251-264. [ Links ]
]]>Vannote, R.L., G.W. Minshall, K.W. Cummins, J.R. Sedeli & C.E. Cushing. 1980. The river continuum concept. Can. J. Fish. Aquat. Sci. 37: 130-137. [ Links ]
Wantzen, K.M. & R. Wagner. 2006. Detritus processing by invertebrate shredders: a neotropical–temperate comparison. J. N. Am. Benthol. Soc. 25: 216-232. [ Links ]
Correspondencia a: Javier Rodríguez-Barrios. Grupo de Biodiversidad y Ecología Aplicada, Instituto de Investigaciones Tropicales, Universidad del Magdalena, Carrera 32 No 22-08, Santa Marta, Colombia. PBX: 4217940; jarodriguezba@gmail.com
Rodulfo Ospina-Tórres. Laboratorio de Insectos Acuáticos, Universidad Nacional de Colombia-Bogotá, Carrera 45 No 26-85, Bogotá, Colombia, PBX 3165000; rospinat@unal.edu.com
Rodrigo Turizo-Correa. Grupo de Limnología Neotropical, Instituto de Investigaciones Tropicales, Universidad del Magdalena Carrera 32 No. 22-08, Santa Marta, Colombia. PBX 4217940; turizortc@yahoo.es
Recibido 25-X-2010. Corregido 06-III-2011. Aceptado 04-IV-2011.
]]>
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}