Variación espacial y temporal en la composición de la dieta de peces invertívoros en un río neotropical, Venezuela
Mario Ortaz, Ricardo Martín & Adriana López-Ordaz
Instituto de Biología Experimental, Universidad Central de Venezuela. Apartado postal 48170, Caracas 1041A, Venezuela; maortaz@hotmail.com, ri14782@yahoo.es, adri_lopezordaz@yahoo.es
Dirección para correspondencia
Instituto de Biología Experimental, Universidad Central de Venezuela. Apartado postal 48170, Caracas 1041A, Venezuela; maortaz@hotmail.com, ri14782@yahoo.es, adri_lopezordaz@yahoo.es
Dirección para correspondencia
Abstract
Spatial and temporal variation in diet composition of invertivore fishes in a tropical stream, Venezuela. Invertivores fishes are an important component of neotropical streams and they represent a link between aquatic invertebrates and piscivorous species. This study evaluated the breadth diet and interspecific food overlap of nine invertivores fish species during three consecutive hydrological phases: falling (December/07, January/08, February/08 and ]]> i) and Morisita (IM) were used to estimate the breadth diet and interspecific food overlap, respectively. All estimations were made using the numeric ]]> Knodus deuteronoides, Creagrutus bolivari and C. melasma) and five in LW (Thoracocharax stellatus, Moenkhausia lepidura, Cheirodon pulcher, Ctenobrycon spilurus and Aphyocharax alburnus), and one Cyprinodontiformes (Poecilia ]]> ), which was also found in HW. In HW aquatic insects were the main resource consumed by fishes while plant material and terrestrial arthropods were secondary resources. In LW the fishes ingested all of these items in addition to zooplankton (Copepoda, Cladocera and larval stages of Decapoda). However, there was a temporal replacement with a predominance of zooplankton in falling and low water. In general, the breadth diet decreased during the falling water in both sections and increased in rising water. However, the average breadth diet was higher in HW. The interspecific food overlap was high in HW while low values were more frequent in LW and ]]>
Key words: trophic ecology, Characiformes, invertivores fishes, neotropical stream, Venezuela.
Resumen
La ictiofauna invertívora es un componente importante en los ríos neotropicales y representa un enlace entre los invertebrados acuáticos y los piscívoros. En el presente trabajo se evaluó la amplitud y sobreposición interespecífica de dieta de nueve especies de peces invertívoros durante tres fases hidrológicas consecutivas: descenso (diciembre/07, enero/08, febrero/08, marzo/08), aguas bajas (abril/08) y ascenso de aguas (junio/08) en dos secciones de un río venezolano ubicadas a distinta altitud (subcuenca alta, SA y subcuenca baja, SB). Los peces se recolectaron con una red de cerco (0.5cm entrenudos) entre las 8:00 y 11:00 horas. La dieta de cada especie se evaluó usando un índice de importancia relativa (IIR) que incluyó el número, peso y frecuencia de ocurrencia de los items consumidos. Para estimar la amplitud y sobreposición interespecífica ]]> i) y Morisita (IM), respectivamente. Todas las estimaciones se realizaron con la frecuencia numérica de las presas consumidas. Se capturaron nueve especies, ocho Characiformes, tres en SA (Knodus deuteronoides, Creagrutus bolivari y C. melasma) y cinco en SB (Thoracocharax stellatus, Moenkhausia ]]> , Ctenobrycon spilurus y Aphyocharax alburnus) y un Cyprinodontiformes (Poecilia reticulata), capturada en SA. En SA el principal recurso consumido fueron insectos acuáticos mientras que el material vegetal y los artrópodos terrestres fueron recursos secundarios. En SB, las especies consumieron estos items además de zooplancton (Copepoda, Cladocera y estadios larvales de ]]>
]]> Palabras clave: ecología trófica, Characiformes, peces invertívoros, río neotropical, Venezuela.
La ictiofauna invertívora es un componente conspicuo e importante de los ríos neotropicales ya que está conformado por un gran número de especies que ejercen una permanente presión depredadora sobre sus presas y representan una importante fuente de alimento para los piscívoros (Zaret & Rand 1971, Winemiller 1990, Flecker 1992, Ortaz 1992, Taphorn 1992, Machado-Allison 2005, Winemiller 2005, Pouilly et al. 2006, Trujillo-Jiménez & Toledo-Beto 2007).
En muchos ríos, los invertebrados presas están representados por una diversa ]]> et al. 1980, Wootton 1990, Allan 1995, Wetzel 2001, Barbee 2005). La disponibilidad de esta materia orgánica varía tanto a escala espacial como temporal, y está regulada por la disponibilidad de nutrientes, el tipo, desarrollo y fenología de la vegetación ribereña, las diferencias hidrogeomorfológicas del canal y las fluctuaciones en la descarga hidraúlica, entre otros (Vannote et al. 1980, Merritt & Cummins 1984, Welcomme 1985, Lowe-McConnell 1987).
De manera general, se considera a la época de aguas bajas como la de menor disponibilidad de alimento para los peces invertívoros. Sin embargo, esto no necesariamente ocurre a todo lo largo de una cuenca ya que dependerá de las diferencias que existan en el gradiente altitudinal. Al respecto, en muchos ríos de tierras bajas se ]]> donde disminuye la disponibilidad del bentos comenzaría al inicio de descenso de aguas y su menor valor debiera ocurrir cuando el río presenta su menor nivel hidrométrico. Adicionalmente, pueden ocurrir cambios importantes ]]>
Con base en lo anterior, en el presente trabajo se evaluó en un río neotropical en el que existe un evidente gradiente altitudinal (Ortaz 2001, Ortaz & Rocha 2001, Martín 2007) si en la ictiofauna invertívora ocurre un cambio importante, a escala espacial y temporal, tanto en la composición de dieta como en su amplitud y sobreposición ]]>
Materiales y métodos
Area de estudio: Se evaluaron dos secciones del cauce principal del río Orituco, ubicado en el centro-norte de Venezuela. Una de ellas correspondió a la subcuenca alta del río (SA, c.a 800msnm) cerca del límite ]]>
Entre diciembre/07 y junio/08 se realizaron seis muestreos de campo, con una frecuencia mensual, que incluyeron las fases hidrológicas de descenso de aguas (diciembre/07, enero/08, febrero/08 y marzo/08), aguas bajas (abril/08) e inicio de ascenso de aguas (junio/08), las cuales se definieron de ]]> la información meteorológica disponible. Para confirmar la fase hidrológica de cada sección, en cada muestreo se midió el ancho del canal (precisión: 0.01m), el perfil de profundidad (precisión: 0.01m) y la velocidad superficial de la corriente (m/s), con el método del flotador, con lo cual se calculó la descarga hidráulica (m3/s) (Wetzel & Likens 2000).
]]> Para la recolecta de los peces se empleó una red de cerco (0.5cm de abertura entre nudos). El esfuerzo de pesca en cada sección varió entre cuatro y diez lances, siendo seis el número más frecuente. La captura se realizó entre las 8:00 y 11:00 horas. En cada muestreo se realizó primero el trabajo en SA y al día siguiente en SB. Los peces recién capturados se preservaron en frío hasta su muerte, posteriormente en una solución de formol ]]>
Métodos: Los peces se identificaron según Géry (1977) y Taphorn (1992). Para cada ejemplar se estimó: la longitud estándar (precisión: 0.05mm) y el peso corporal húmedo (precisión: 0.01g). Posteriormente bajo lupa se extrajo el tracto digestivo. Se describió la composición de la dieta ]]> et al. 2006). Los insectos acuáticos consumidos se identificaron según Merritt & Cummins (1984), Roldán (1996) y Fernández & Domínguez (2001).
]]> Se estimó la amplitud de dieta con el índice de Levin (Bi =1/ΣPij2) (Chesson 1978, Pouilly et al. 2003, Pouilly et al. 2006). Pij=proporción numérica de la presa “j” en la dieta de la especie “i” y N=número de tipos de presas consumidas. La sobreposición trófica interespecífica se estimó con la modificación de Horn (1966) del índice de Morisita IM=2Σ(Pij Pkj)[Σ (Pij2+Pkj2)]-1 ]]> (Zaret & Smith 1984, Smith 1985). Pij=proporción numérica del ítem “j” en la dieta de la especie “i” y Pkj=proporción numérica del ítem “j” en la dieta de la especie “k”.
Resultados
]]> Composición de la dieta: Las especies analizadas fueron: Knodus deuteronoides (SA) (n=224), Creagrutus bolivari (SA) (n=116), Creagrutus melasma (SA) (n=60), Poecilia reticulata (SA) (n=49), Thoracocharax stellatus (SB) (n=112), Moenkhausia lepidura (SB) ]]> Cheirodon pulcher (SB) (n=38), Ctenobrycon spilurus (SB) (n=38) y Aphyocharax alburnus (SB) (n=28).
La dieta de las especies en SA estuvo constituida por invertebrados acuáticos (insectos, gastrópodos y ostrácodos), artrópodos terrestres y material vegetal alóctono. Los insectos representaron el grupo ]]> K. deuteronoides). Los órdenes Coleoptera, Diptera, Ephemeroptera y Trichoptera fueron los más consumidos por los carácidos evaluados y sólo Diptera y Ephemeroptera fueron consumidos en una proporción importante por P. reticulata (Cuadro 1). Los artrópodos terrestres fueron un ]]> C. bolivari y K. deuteronoides, mientras que en C. melasma y P. reticulata su consumo fue insignificante. El consumo de material vegetal alóctono fue alto en las cuatro especies y a lo largo del lapso evaluado. K. deuteronoides consumió el mayor número de items (13), seguida por C. bolivari (11), C. melasma (8) y P. reticulata (5). El consumo promedio de insectos acuáticos fue parecido entre los carácidos y bajo en P. reticulata. El consumo promedio de invertebrados terrestres fue muy bajo en C. melasma, representó cerca del 15% del consumo total ]]> C. bolivari y K. deuterodonoides y no fue una presa depredada por P. reticulata. El consumo promedio de material vegetal alóctono fue alto en P. reticulata y menor en los carácidos.
La dieta de los invertívoros ]]> Cuadro 1). En general, aunque en SB la dieta estuvo constituida por un número parecido de items al encontrado en SA (12 en SB vs 13 en SA), hubo un cambio importante en su composición ya que sólo el 47% de las presas consumidas fueron comunes para ambas secciones. En SB no se consumieron ]]> consumo como por el número de especies que lo depredaron. Otro componente bentónico que ]]> K. deuteronoides y sólo en un mes. En SB la depredación sobre el zooplancton fue importante en buena parte del registro, el cual nunca formó parte de la dieta en SA. El consumo promedio de zooplancton nunca fue inferior al 25 % en cuatro de las cinco especies evaluadas y este fue parecido en A. alburnus y C. pulcher. Sólo en T. stellatus no se ]]> A. alburnus los copépodos llegaron a representar un porcentaje alto. El consumo de preadultos de Decapoda fue bajo en las especies evaluadas. Los artrópodos terrestres representaron en SB un recurso relativamente poco importante en la mayoría de las especies con excepción de T. stellatus para la cual representó su ]]> T. stellatus. De hecho en especies como C. pulcher llegó a representar hasta la totalidad de su dieta (junio/08). M. lepidura fue la especie que consumió el mayor número de tipos de presas y en casi la totalidad del registro consumió, de manera ]]> C. pulcher y T. stellatus fueron las más estrechas mientras que las de A. alburnus y C. spilurus mostraron una condición intermedia.
]]> Fig. 1). Por el contrario, en SB hubo una progresiva disminución en el consumo de insectos acuáticos desde el inicio de descenso de aguas hasta aguas bajas con un posterior incremento en ascenso de aguas. El zooplancton representó en SB la principal comunidad depredada en ]]> T. stellatus.
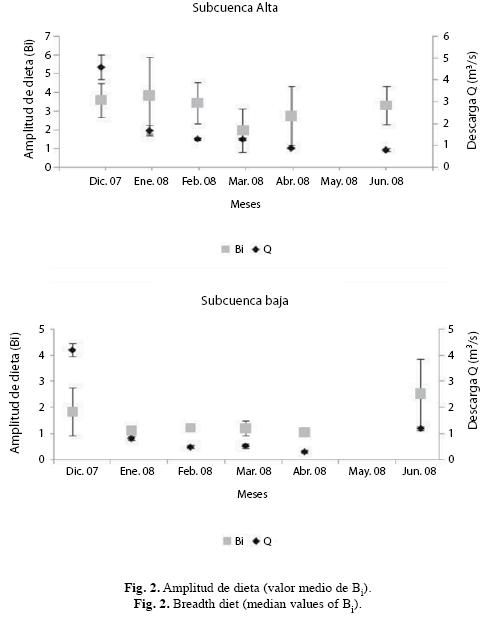
Amplitud de dieta: En C. bolivari, la menor amplitud se obtuvo en aguas bajas (abril/08) y la mayor en descenso de aguas (enero/08). La menor amplitud no correspondió con una reducción general del consumo de presas y, por el contrario, el promedio de presas consumidas en este mes fue elevado (X: 7.1 presas/pez), y solo superado por el obtenido en marzo (X: 8.2 presas/pez). En este caso sólo un tipo de presa (Trichoptera) representó el 72% de todas las consumidas, que en este mes incluyó a Ephemeroptera (9.8%), Artrópodos terrestres (5.5%), Coleoptera (4.5%), Diptera (3.2%), Odonata (2.5%) y Hemiptera (2.5%). El mayor valor ]]> C. melasma, el menor valor se obtuvo en marzo/08 (descenso de aguas) y el mayor en junio/08 (ascenso de aguas) (Cuadro 2). La tendencia general obtenida consistió en un descenso progresivo de diciembre/07 hasta marzo/08, con excepción de febrero/08 y a partir de marzo/08 incrementó hasta ascenso de ]]> K. deuteronoides la tendencia ]]> En esta especie la menor amplitud de dieta se obtuvo en marzo/08 (descenso de aguas) y la mayor en abril/08 (aguas bajas). En el mes de menor amplitud de dieta el consumo de presas fue elevado (X: 7.1 presas/pez) y el consumo de las presas fue poco proporcional ya que sólo Diptera representó el 67% del total y el resto quedó repartido entre Ephemeroptera (12 ]]> y un último grupo constituido por Coleoptera, Hemiptera y Odonata (6% restante). La mayor amplitud correspondió, igualmente, con un elevado consumo de presas (X: 5.1 presas/pez) pero más uniforme ya que la variación de la proporción de las cuatro presas más depredadas fue de sólo 9 % (Trichoptera: 16%, Diptera: 21%, Artrópodos terrestres: 22% y Ephemeroptera: 25%) mientras que el resto representaron en conjunto el 16% del total. P. ]]> fue el invertívoro con el consumo más bajo de invertebrados acuáticos y terrestres y, entre diciembre/07 y febrero/08, su dieta consistió exclusivamente de material vegetal alóctono y sólo entre marzo/08 y junio/08 depredó insectos acuáticos pero sin excluir al material vegetal. La menor amplitud de dieta se obtuvo en marzo/08, en el que fue alto el consumo de insectos acuáticos (X: 4.5 presas/pez). Este bajo valor coincidió con el consumo de un solo tipo de presa (Diptera). La ]]> junio: 3 presas/pez) y un incremento en la variedad de presas que incluyó Diptera (50%), Ephemeroptera (41%) y Trichoptera (9%).
La amplitud de dieta obtenida en C. bolivari fue superior a la del resto de las especies de SA, con excepción de lo obtenido en abril/08.
En C. spilurus, la menor amplitud de dieta se obtuvo en enero/08 y la mayor en diciembre/07 (ambas en descenso de aguas). Esta especie no se capturó entre marzo/08 y junio/08. La baja amplitud en enero/08 correspondió con un elevado consumo de Cladocera, quien representó el 99.5% de su dieta, mientras que el resto de las presas representaron el 0.5 % (Decapoda: 0.1%, Copepoda: 0.2%, Diptera: 0.1% y Ostracoda: 0.1%). La ]]> A. alburnus la menor amplitud se obtuvo en marzo/08 (aguas bajas) y la mayor en junio/08 (ascenso de aguas). Esta especie no se capturó en diciembre/07 y abril/08. La reducción de la amplitud en marzo/08 correspondió con el consumo de sólo 2 tipos de presas y en proporciones muy distintas (Copepoda: 99.4% del total y Cladocera: 0.6%), mientras que la mayor amplitud no correspondió con un incremento importante en el número de ]]> M. lepidura, la mínima amplitud se obtuvo en febrero/08 (descenso de aguas) y la máxima en junio/08 (ascenso de aguas). La baja amplitud no correspondió con una disminución del número de tipos de presas consumidas sino con la variación en sus proporciones, que en este caso fue elevada para Cladocera (91.66%) y muy baja para el resto (Ostracoda: 4.16%, Diptera: 2.77% y Artrópodos terrestres: 1.41%). La mayor amplitud ]]> C. pulcher se obtuvo en aguas bajas y se debió al consumo de un solo tipo de presa (Cladocera). Esta especie no se capturó en todo el registro evaluado. La mayor amplitud se obtuvo en descenso de aguas y fue consecuencia del consumo de una mayor variedad de presas con ]]> T. stellatus fue la más especializada, como en C. pulcher, pero en un mayor número de meses (enero/08, abril/08 y junio/08), su amplitud de dieta fue mínima debido al consumo de un solo tipo de presa. Su mayor amplitud se obtuvo al inicio de descenso de aguas y fue consecuencia del consumo de una mayor variedad de presas con intensidades de consumo variables (Hemiptera: 6.8%, Diptera: 12.2%, ]]>
En la mayor parte del registro, la amplitud de dieta de M. lepidura fue superior, con excepción de T. stellatus en diciembre/07 y A. alburnus en febrero/08. En general, la variación en A. alburnus ]]> M. lepidura fue parecida y consistió en una disminución progresiva desde descenso hasta aguas bajas, con excepción de un comportamiento opuesto en febrero/08, para luego incrementar en ascenso de aguas.
La variación general de la amplitud de dieta fue parecida en varias especies y consistió en una disminución a medida que progresaron las fases de descenso y aguas bajas, con un ]]> C. bolivari en SA este mínimo ocurrió en aguas bajas, mientras que en el resto de las especies de esta sección y en M. lepidura en SB este ocurrió en descenso de aguas. En el resto de las especies de SB la menor amplitud se obtuvo tanto en descenso como en aguas bajas. Otro resultado parecido en ambas secciones fue el incremento importante en la amplitud de dieta en ascenso de aguas lo cual ocurrió en todas las especies, con ]]> T. stellatus. La amplitud promedio de dieta resultó mayor en SA (Fig. 2).
{kind=link}
{kind=link}
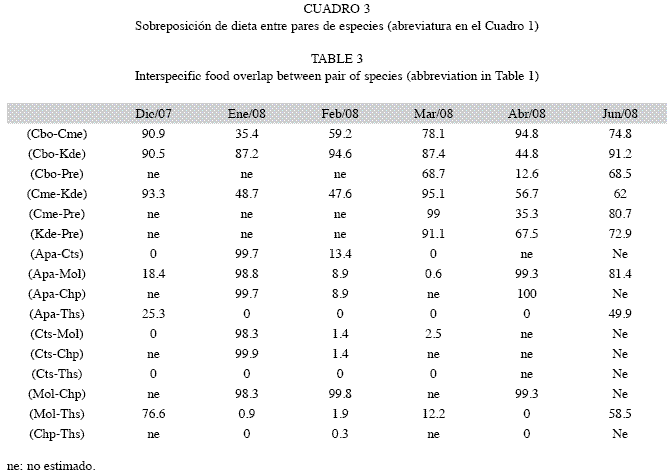
Sobreposición interespecífica de dieta: Los valores de sobreposición de dieta en SA resultaron altos (>50%) en buena parte del registro (Cuadro 3). En esta sección, la sobreposición tendió a ser alta entre los carácidos (C. bolivari, C. melasma y K. deuteronoides) y baja entre éstos y P. reticulata, de hecho el menor valor se obtuvo entre C. bolivari y P. reticulata en aguas bajas (abril/08). La sobreposición no fue recurrentemente alta en un único par de especies y por el contrario se obtuvieron valores altos en todos los pares comparados. Los bajos valores se obtuvieron en enero/08 y abril/08 (IM media: 57.1% y IM media: 52%, respectivamente), y el mayor se obtuvo al inicio del descenso de aguas (diciembre/07) (IM media: 91.6%) (Fig. 3). La variación en los pares C. bolivari - C. melasma y C. bolivari - K. deuteronoides fue parecida entre diciembre/07 y febrero/08 y opuesta en el resto del período. En el par C. melasma - K. deuteronoides y en el período diciembre/07 - marzo/08 la variación fue más parecida a la que ocurrió entre C. bolivari - C. melasma en el mismo lapso, mientras que en abril/08 y junio/08 varió tal y como ocurrió en C. bolivari - K. deuteronoides. En C. bolivari - C. melasma la sobreposición fue alta al inicio de descenso (diciembre/07) y en aguas bajas (abril/08) y sólo en enero/08 fue menor del 50%. Entre C. bolivari - K. deuteronoides fueron frecuentes los valores altos (>80% en todos los meses evaluados), con excepción de aguas bajas (abril/08) (<50%). Al igual que entre C. bolivari y C. melasma, la sobreposición fue alta al inicio de descenso de aguas (diciembre/07) pero también en descenso (febrero/08) y ascenso de aguas (junio/08). La sobreposición entre C. melasma y K. deuteronoides tendió a ser menor ya que fueron más frecuentes los valores menores al 60%, sin embargo, también fue alta al inicio de descenso (diciembre/07) y en aguas bajas (marzo/08). En P. reticulata y debido a que no consumió invertebrados entre diciembre/07 y febrero/08, sólo se pudo estimar la sobreposición de dieta con los carácidos capturados en el resto del período. Su dieta tendió a ser diferente a la de C. bolivari por su baja sobreposición y fue parecida a la de C. melasma y K. deuteronoides en marzo/08 y junio/08 y baja en abril/08. No se observó una relación entre la sobreposición y el porcentaje de presas comunes ya que, por ejemplo, en meses en los que éste fue bajo, como entre C. bolivari y C. melasma en diciembre/07, enero/08, febrero/08 y abril/08, se obtuvieron por igual valores bajos y altos de sobreposición. Lo mismo ocurrió en otros pares como C. melasma y K. deuteronoides, en los que el porcentaje de presas comunes fue parecido en parte del lapso evaluado (ca 40%), y sin embargo la variación de la sobreposición de dieta fue grande. Aunque no se obtuvo relación entre estas variables, el porcentaje de presas comunes siempre fue alto entre C. bolivari y K. deuteronoides y bajo entre P. reticulata y los carácidos. Tampoco se observó una relación entre la sobreposición de dieta y el promedio de presas consumidas/especie, ya que, por ejemplo, el mayor promedio obtenido en marzo/08, no correspondió con una alta sobreposición de dietas. Igualmente, los bajos promedios de presas consumidas no correspondieron necesariamente con bajos valores de sobreposición.
{kind=link}
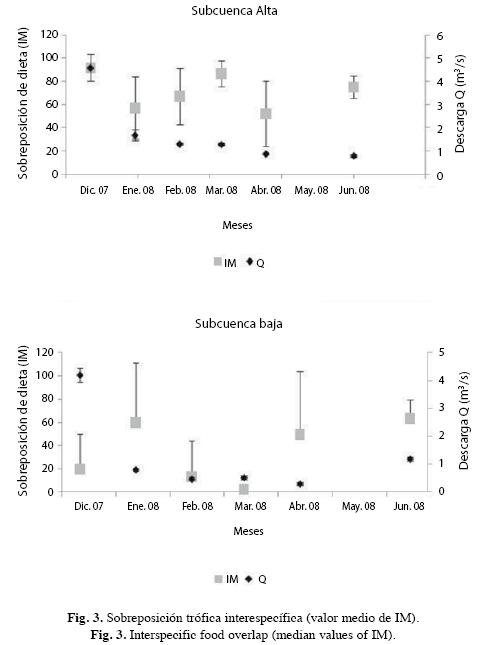
En SB se obtuvieron, al igual que en SA, valores de sobreposición superiores al 50%, sin embargo lo frecuente fueron valores bajos. En enero/08 (descenso de aguas) se obtuvieron valores altos en la mayoría de los pares comparados así como entre A. alburnus y M. lepidura en abril/08 (aguas bajas) y junio/08 ]]> A. alburnus y M. lepidura. La variación de la sobreposición de dieta entre diciembre/07 y marzo/08 fue parecida en los pares A. alburnus - C. spilurus, A. alburnus - M. lepidura y C. spilurus - M. lepidura (Cuadro 3) y fue baja al inicio de descenso de aguas (diciembre/07), incrementó en enero/08 y disminuyó hasta marzo/08. La sobreposición entre A. alburnus y M. lepidura fue alta en ]]> M. lepidura - C. pulcher, todos los valores fueron altos, sin embargo en este caso el número de individuos analizados de C. pulcher en abril/08 fue bajo y probablemente no representó el comportamiento de la población. La dieta de T. stellatus fue la más ]]> M. lepidura, esta especie sobrepuso su dieta en más del 50% pero sólo al inicio de descenso y ascenso de aguas.
En SA ca del 80% de los estimados de IM fueron superiores al 50% mientras que en SB esto se obtuvo sólo en el ]]> Fig. 3). En SB tampoco se obtuvo una relación entre la sobreposición de dieta y el porcentaje de presas comunes y el promedio de presas consumidas. En casi todo el período evaluado se encontró una tendencia opuesta en la variación temporal de la sobreposición trófica interespecífica promedio entre secciones.
Discusión ]]>
En descenso y aguas bajas, las presas más consumidas en SB, con excepción de T. stellatus, no fueron los insectos acuáticos sino el zooplancton. Esto podría ser consecuencia de la disminución en la disponibilidad de insectos acuáticos bentónicos por condiciones ambientales adversas ]]> A. alburnus, C. spilurus y M. lepidura en ascenso y aguas bajas mientras que en descenso no hubo un consumo de zooplancton. En SA, condiciones como la mayor variedad de ]]>
Los cambios observados indican que dentro de un mismo gremio trófico y en el mismo ambiente puede ocurrir una variación importante en el tipo de presa consumida y en su intensidad de depredación. Estos cambios ocurren tanto a una escala espacial (altitudinal) como temporal (entre fases hidrológicas) y en buena medida son inducidos inicialmente por los cambios en la descarga ]]> T. stellatus quien consumió fundamentalmente insectos terrestres. Una especie abundante en SB fue M. lepidura la cual si bien consumió principalmente presas autóctonas, ]]> T. stellatus, a los invertebrados terrestres y de hecho, fue la que sobrepuso más su dieta con ésta (IM media: 18%) y compartió el mayor porcentaje de presas comunes (X: 29%).
Si bien los invertebrados son un recurso de uso frecuente, su importancia varía dependiendo del ambiente. Pouilly et al. ]]>
]]> En SB, M. lepidura fue la que mostró la dieta más amplia debido a la mayor variedad de tipos de presas ingeridas y a la mayor intensidad y proporcionalidad en su consumo. Este resultado fue equivalente al obtenido en SA para K. deuteronoides y C. bolivari. C. pulcher fue un caso interesante y un ]]> et al. 2006).
En ambas secciones la amplitud promedio de dieta disminuyó a medida que progresaron las fases de descenso y aguas bajas, con un incremento importante en ascenso de aguas, que en SB consistió en un reemplazo del zooplancton por los insectos acuáticos. Además, los valores fueron ]]> estimado de amplitud de dieta. Por ejemplo, la amplitud en C. bolivari en abril/08 disminuyó considerablemente, y no se debió a una reducción del consumo de presas, ya que inclusive éste fue proporcionalmente mayor al de otros meses en los que su dieta fue más amplia. La causa de esta reducción fue el consumo menos equitativo de las ]]> K. deuteronoides en marzo/08, mes en el que, aunque el promedio de presas consumidas fue el más alto del registro (X: 7 presas/pez), éste fue desproporcionado. El elevado consu-mo de sólo un tipo de presa también explicó la reducción de la dieta de C. melasma y A. alburnus en marzo/08 ]]> C. spilurus en enero/08 (Cladocera: 99.5%) y de M. lepidura en febrero/08 (Cladocera: 91.7%). La reducción extrema en C. pulcher en abril/08 y en T.stellatus en enero/08, abril/08 y en junio/08 se debió al consumo de un solo tipo de presa (cladóceros en C. pulcher y ]]> T. stellatus). Esto indica que una reducción en la amplitud de dieta no necesariamente estará asociada a una disminución del consumo de presas sino a una variación en las proporciones de consumo.
Los resultados mostraron que en SA los invertívoros dependen de la misma comunidad de presas (insectos ]]>
La sobreposición de dieta obtenida puede sub o sobreestimar la real debido a que no considera la posible ]]> K. deuteronoides y P. reticulata en marzo/08 fue IM: 0.9114. Sin embargo, K. deuteronoides compartió con P. reticulata sólo uno de los siete tipos de presas consumidas (14% de ellas), mientras que P. reticulata compartió el único ]]> una opción podría ser incorporar al cálculo de sobreposición, una variable, como el porcentaje de presas comunes consumidas, que permitiría obtener valores corregidos de sobreposición y un valor por especie. A continuación se detalla el procedimiento:
]]> IMspA–spB=0.9114.
IMspA=0.9114x14%=0.1276
(14% representa el porcentaje de tipos de presas que la especie A comparte con la especie B).
IMspB=0.9114x100%=0.9114
(100% representa el porcentaje de tipos de presas que la especie B comparte con la especie A).