Abstract
Morphology of the species Hyphessobrycon, heterorhabdus group (Characiformes: Characidae) in Colombia. Hyphessobrycon is the most numerous and morphologically complex genus of Characidae, and includes 18 species reported for Colombia, from which thirteen belong to the heterorhabdus-group different methods have been proposed for species identification within this genus. This study used these species to undertake a morphogeometric analysis by the Box Truss and Thin Plate Splin (TPS) methods; 13 homologous landmarks type I and three type II were used. The result of cluster analysis indicated that these species are represented by two big groups: robust and thin. The uniform and non uniform components, and the principal warps (WP) and partial warps (Wparc), described the shape changes related to body depth, involving relative displacements of the appendicular skeleton and the cephalic region. All species were characterized by allometric growth with the exception of Hyphessobrycon mavro, who presented an isometry between the landmarks 6-7. Two new measures are proposed for species discrimination in the group (snout-supraoccipital spine distance and supraoccipital spine-pelvic fin). Rev. Biol. Trop. 59 (2): 709-725. Epub 2011 June 01.
Key words: box truss, landmarks, tropical fish, thin plate spline (TPS).
Resumen
El género Hyphessobrycon incluye dieciocho especies reportadas para Colombia de las cuales 13 son miembros del grupo heterorhabdus; con las mismas, se realizó un análisis morfogeométrico por medio del Método de Cerchas (Box Truss) y Láminas Delgadas (TPS). Se usaron 13 hitos homólogos tipo I y tres tipo II. El resultado del análisis cluster, llevado a cabo con cada una de las metodologías indicó que estas especies están representadas en dos grandes grupos: robustas y delgadas. Las deformaciones parciales, los componentes uniformes, no uniformes, Warps principales y Warps parciales describieron cambios de la forma relacionados con la profundidad del cuerpo e involucraron desplazamientos relativos del esqueleto apendicular y de la región cefálica. Todas las especies se caracterizan por presentar crecimiento alométrico excepto Hyphessobrycon mavro con isometría entre los hitos 6-7. Se plantean dos nuevas medidas discriminantes (hocico-supraoccipital y supraoccipital-aleta pélvica) para las especies del grupo.
Palabras clave: cerchas, hitos, pez tropical, thin plate spline (TPS).
]]> Hyphessobrycon Durbin (en Eigenmann 1908) es un género de pequeños peces carácidos que cuenta con alrededor de 125 especies válidas (García-Alzate et al. 2010a) se encuentran distribuidas en todas las cuencas del norte de América del sur. Además, se ha planteado el ]]> heterorhabdus, definido inicialmente por presentar una banda oscura en la parte lateral del cuerpo (García-Alzate et al. 2008a). También, se ha planteado como grupo monofilético, con base en dos caracteres osteológicos: 13 a 14 epipleurales y metapterigoide sin proyección ventral y no en contacto con el ]]>
García-Alzate et al. (2010a) reportan 15 especies de Hyphessobrycon para Colombia, actualmente el número de especies asciende a 22: H. bentosi Durbin, H. columbianus Zarske & ]]> H. condotensis Regan, H. ecuadorensis Eigenmann, H. erythrostigma Fowler, H. panamensis Durbin, H. saizi Géry, H. sweglesi Géry, H. sebastiani García-Alzate et al., H. agulha Fowler, H. acaciae García-Alzate et al., H. amaronensis García-Alzate et al., H. diancistrus Weitzman, H. heterorhabdus (Ulrey), H. metae Eigenmann & Henn, H. mavro García-Alzate et al., H. niger ]]> et al., H. ocasoensis García-Alzate & Román-Valencia, H. oritoensis García-Alzate et. al., H. poecilioides Eigenmann, H. proteus Eigenmann, H. taguae García-Alzate et al., las últimas 13 pertenecientes al grupo heterorhabdus.
]]> Dentro del género Hyphessobrycon los análisis tradicionales de la forma han sido ampliamente usados para discriminar diferencias entre especies, pero han sido poco útiles para analizar la variación entre grandes grupos, es así que los análisis de forma diferentes al tradicional como es el método de morfometría geométrica, representa un ]]> Hyphessobrycon (García-Alzate et al. 2010b), además, permite la distinción entre forma y talla, comparar organismos mediante sus estructuras homólogas, cuantificar la variabilidad de la forma en múltiples escalas espaciales, y estudiar las posibles correlaciones entre la variabilidad y otros parámetros morfológicos y/o ambientales (Rohlf 2000, Zelditch et al. 2004, Ruiz-Calderón & Cipriani 2006). Así, el objetivo de este trabajo fue realizar un análisis morfogeométrico de las especies del género Hyphessobrycon grupo heterorhabdus presentes en Colombia.
Materiales y Métodos
Se analizaron 612 ejemplares adultos provenientes de algunas de las instituciones y museos registrados en la siguiente página web: http://www.asih.org/codons.pdf entre las que se incluyen: AUM, IAvHP, ICNMHN, MBUC-V, MCNG, MCP y MUSM, con la adición de: ]]>
El análisis ]]> H. acaciae, H. agulha, H. amaronensis, H. ocasoensis, H. poecilioides, H. proteus e H. taguae a partir de la metodología propuesta por Rolff (1996). Otras especies del grupo heterorhabdus fueron ]]> Fig. 1). Entonces, se utilizó la opción macro del lente para adquirir ]]> Fig. 1).
La talla de cada ]]>
]]> La disparidad entre las configuraciones alineadas fue descrita con un análisis de láminas finas, conocido como TPS (Thin Plate Spline) (Bookstein 1991, Zelditch et al. 2004). En este análisis cada una de las configuraciones (CI) fue comparada con la configuración consenso (CC) de toda la muestra en dos pasos: 1) Al sobreponer CI a CC y 2) Y al deformar el plano de CI hasta hacer coincidir las posiciones de todos sus hitos con las posiciones de los ]]> affine y el segundo en uno no uniforme o no affine; las deformaciones asociadas a este componente fueron resultado de la adición de los efectos de todos los Wparc (Warp Parciales) calculados para cada ]]> et al. 2004, Rohlf et al. 1996).
Por lo tanto, se describió la forma y variabilidad en cada especie a partir de contrastar las configuraciones individuales (individuos) con la CC de la especie en el programa tpsSplin (Rohlf ]]> et al. 2004); el análisis de los WP describieron la magnitud de las deformaciones (locales o globales) con un factor de escala de 0.22.
Para probar si las ]]> 0=isometría) fue evaluada de cuatro formas: la prueba generalizada F de Goodall, Lambda de Wilks, análisis de permutación y regresión lineal simple; todos ellos llevados a cabo en el programa tpsRegr (Rohlf 2003).
Así mismo, se realizó un diseño de cerchas mediante la metodología propuesta por Strauss & Bookstein (1982) y posteriormente seguida por García-Alzate et al. (2010b); donde se incluyen las 13 especies del grupo heterorhabdus, la ]]> et al. 2008), donde se obtuvieron 28 coordenadas y se realizaron tres análisis: 1) Deformaciones parciales con los datos transformados a distancias procrustes, 2) Análisis de componentes principales (ACP), donde las coordenadas fueron transformadas con el método de Burnaby para especies con ]]> et al. 2008), para evaluar la similitud de formas entre las especies por medio del algoritmo UPGMA.
]]> Material examinado: Hyphessobrycon agulha: Colombia: IAVH-P 8333, (22) (26.2-47.2mm de LE); Leticia, afluente rio Purité, Amazonas, 25 mar. 2001. IAVH-P 8335, (32) (32.6-47.9mm de LE); Leticia, afluente quebrada Matamatá, Amazonas, 18 mar. 2001. IAVH-P 8345, ]]> H. diancistrus: ver García-Alzate et al. 2008c. H. heterorhabdus: Colombia: ICNMHN 5063, (5) (26.4-31.4mm de LE); Río Puré, Letícia, Amazonas, 8 ene. 2000. Brasil: ]]> H. metae: Colombia: ICNMHN 12274, (8) (24.0-30.0mm de LE). IUQ 2340, (2) (22.1-24.1mm de LE); Brazuelo caño Guayapa, playa rica, Vista Hermosa, Meta, 9 jul. 2008. IUQ 2341, (1) (23.1mm de LE); caño 1km antes de las Brisas, Vista Hermosa, Meta, 10 jul. 2008. IUQ 2342, (4) (21.7-27.3mm de LE), 1km antes del caño Guapaya en la vía playa rico, Vista Hermosa, Meta, 9 jul. 2008. MPUJ 3801, (7) (21.6-24.4mm de LE). MPUJ 3848, (15) (21.6-27.7mm de LE). Venezuela: MCNG 32469, (18) (21.9-26.0mm de LE), Parque Nacional ]]> H. ocasoensis: ver García-Alzate & Román-Valencia 2008a. H. poecilioides: ver García-Alzate et al. 2010c. H. proteus: ver García-Alzate et al. 2010c. H. amaronensis: ver García-Alzate et al. 2008c. H. taguae: ver García-Alzate et al. 2008c. H. mavro: ver García-Alzate et al. 2010c. H. niger: ver García-Alzate et al., 2010c y H. acaciae: ver García-Alzate et al. 2010c.
Resultados
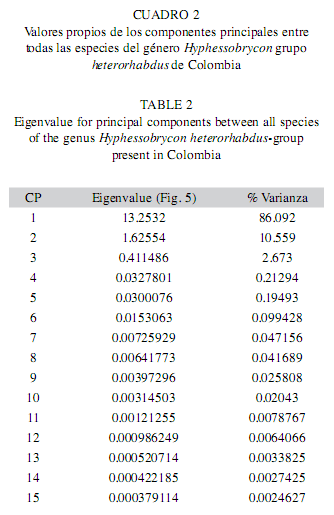
Disparidad general: El valor ]]> Cuadro 1).
]]> La mayor explicación sobre las deformaciones de las especies es dada por el componente uniforme (>50%), mientras cambios locales (componente no uniforme) presentaron una menor influencia sobre la forma general de las especies. En H. agulha las deformaciones del componente uniforme describieron el desplazamiento anterior de la región cefálica, mientras la región dorsal presenta un desplazamiento hacia el extremo posterior, explica el 89% ]]> Fig. 2). Mientras que en H. amaronensis el componente uniforme describió el desplazamiento de la región de la aleta hacia el extremo posterior del cuerpo, entre tanto, la zona ventral se desplazó hacia la región anterior, explicó el 61% de la variación de la forma. El ]]>

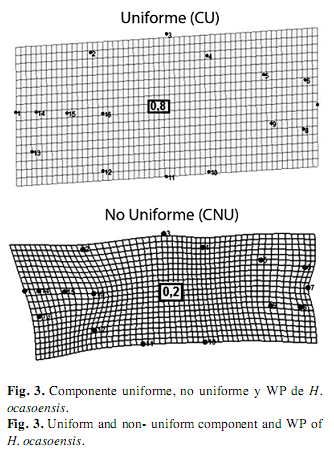
En H. ocasoensis, el componente uniforme describió cambios en la región de la aleta dorsal hacia la parte anterior, mientras la región ventral se desplazó hacia el extremo posterior, lo cual describe el incremento de la profundidad del cuerpo en la especie, el componente no uniforme describió el movimiento hacia la parte posterior de la región opercular, lo cual reveló el 20% de variación de la forma en ésta especie (Fig. 3), al mismo tiempo en H. poecilioides, el componente uniforme describió el desplazamiento de los hitos de la zona anterior hasta la región posterior del cuerpo, y hacia la región anterior de los de la zona ventral, este componente expresó el 79% de la variación de la forma; por otro lado, el componente no uniforme señaló una contracción de la región media ventral en donde explica el 21% de la variación de la forma.
]]>
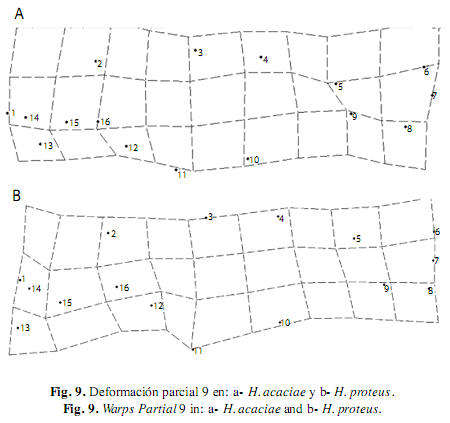
En H. proteus, el componente uniforme solo describió el 38% de la variación total en el cual se observó desplazamientos no muy marcados desde la espina supraoccipital al inicio de la aleta dorsal y hacia la parte anterior del cuerpo, entre tanto con el componente no uniforme se presentaron desplazamientos de los hitos de la región dorsal hacia arriba y de los hitos de la región ventral de tal manera que resulta en una expansión, a su vez se visualizó una contracción entre estos hitos y originó un acercamiento entre la base de las aletas y explicó el 62 % de la variabilidad total. Así mismo, en H. taguae, la deformación descrita por el componente uniforme resultó del desplazamiento anterior y hacia arriba de los hitos de la zona dorsal del cuerpo que dan origen a la expansión de la parte media del cuerpo y revelo el 82% de la variabilidad total, mientras que el componente no uniforme describió una expansión de la cabeza y una contracción en el extremo ventral del pedúnculo caudal y explicó 18% de la variación de la forma de esta especie.
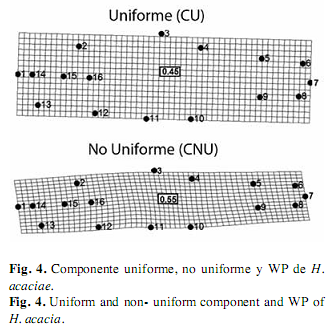
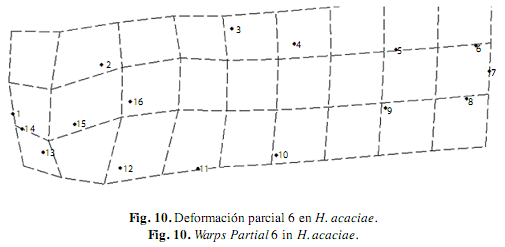
En H. acaciae la deformación explicada por medio del componente uniforme resultó del desplazamiento de los hitos de la zona dorsal hacia atrás y de los hitos de la zona ventral hacia delante, estos movimientos dieron origen a la expansión de la región media del cuerpo, este componente explicó el 55% de la variación morfológica de esta especie, y el componente no uniforme describió la contracción longitudinal del cuerpo debido al desplazamiento hacia arriba de todos los hitos de la zona ventral, lo cual explicó el 45% de la variación (Fig. 4).
Se obtuvieron 13 WP para cada especie, los cuales describieron la magnitud de los ejes de deformación general y local para cada una. La variación morfológica interespecificas presentó alta similaridad (>50%), sólo un pequeño porcentaje explicó la variación morfológica, desde los cuales se puede plantear diferencias de los principales ejes de deformación de las especies.
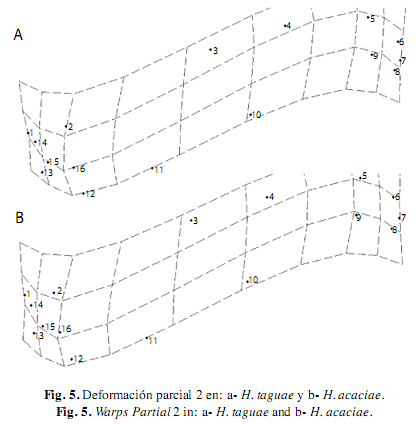
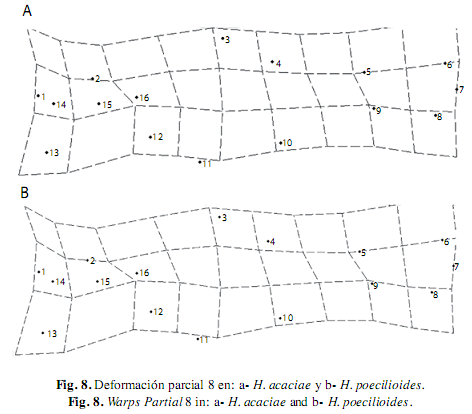
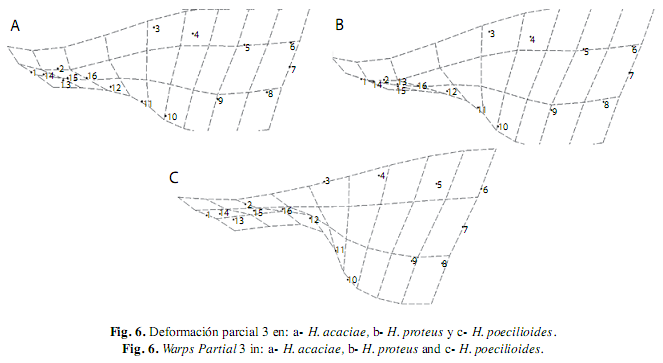
El WP 1 posee el menor autovalor, describe una deformación generalizada en todas las especies, la curvatura del cuerpo y el acortamiento de la cabeza a partir de la expansión de la parte media para todas las especies analizadas, excepto en H. poecilioides e H. amaronensis, en la que el WP 1 detalló la contracción sobre la región caudal. El WP 2 contrasta la exposición de la parte anterior y posterior del cuerpo, en todas las especies se observó mayor exposición de la parte anterior debido a la expansión de la región dorsal (hitos 3 y 4) y de la contracción dorsal de la región pélvica (hito 11); excepto en H. taguae e H. acaciae en las que el WP 2 presentó una mayor expansión en la región caudal en contraste con la menor exposición de la parte anterior del cuerpo (Fig. 5). El WP3 describió la contracción dorsal de la región caudal (hitos 8, 9, 10 y 11) en contraste a la expansión de la región cefálica, excepto en H. poecilioides, H. proteus y H. acaciae en donde la contracción dorsal del WP 3 se mostró en la región cefálica, mientras se contempló la expansión de la región caudal (Fig. 6).
]]>
{kind=link}