Escape tactics and effects of perch height and habituation on flight initiation distance in two Jamaican anoles (Squamata: Polychrotidae)
William E. Cooper, Jr.
Department of Biology, Indiana University Purdue University Fort Wayne, Fort Wayne, IN 46805, USA; cooperw@ipfw.edu
Dirección para correspondencia
Abstract
Escape by Anolis lizards is influenced by microhabitats and fight initiation distance increases with predation risk. Differences in microhabitat use among ecomorphs affect escape behavior, but only two studies have reported ecomorphological differences in flight initiation distance among Greater Antillean species. I studied effects of predation risk and microhabitats on escape behavior by conducting field experiments using two species of anoles, Anolis lineatopus and A. grahami, on the campus of the University of the West Indies at Mona, Jamaica. Because ecomorphological variation of anoles has evolved independently within each island of the Greater Antilles, but relationships between ecomorphs and escape behaviors are poorly known, I characterized microhabitat use and escape tactics, and determined relationships between flight initiation distance and two risk factors, habituation to human presence and perch height, in Anolis lineatopus, a trunk-ground anole and A. grahami, a trunk-crown anole. Sample sizes for A. lineatopus and A. grahami were 214 and 93, for microhabitat use and escape destinations, 74 and 34 for human presence and 125 and 34 for perch height. The two species occurred in similar microhabitats and exhibited similar escape tactics, but exhibited key differences expected for their ecomorphs. Both species were sighted frequently on the ground and on trees, but A. lineatopus were more frequently on ground and were perched lower than A. grahami. Both species escaped from ground to trees and when on trees hid on far sides and escaped without changing climbing direction with equal frequency. The frequency of fleeing upward was greater for A. grahami than A. lineatopus. Both species exhibited habituation by having shorter flight initiation distances in areas with more frequent exposure to people. In both species flight initiation distance increased as perch height decreased because, lizards had to climb farther to be out of reach when perched lower. The relationship between flight initiation distance and perch height may apply to other anole ecomorphs that flee upward when low perched on trees. Rev. Biol. Trop. 58 (4): 1199-1209. Epub 2010 December 01.
Key words: ecomorphs, escape tactics, flight initiation distance, habituation, perch height, Polychrotidae, Squamata.
]]> ResumenEl escape de las largarijas Anolis está influenciado por el microhábitat y la distancia de iniciación de escape incrementa el riesgo de depredación. Las diferencias en el uso de microhábitats entre ecomorfos afecta el comportamiento de escape, pero sólo dos estudios han reportado diferencias ecomorfológicas en la distancia de iniciación de escape entre las especies de las Antillas Mayores. Se estudió el efecto de riesgo de depredación y la influencia del microhábitat en el comportamiento de escape, mediante la realización de experimentos de campo con Anolis lineatopus y A. grahami, en el campus de la Universidad West Indies en Mona, Jamaica. Debido a que las variaciones ecomorfológicas de Anolis han evolucionado independientemente en cada isla de las Antillas Mayores, la relación entre ecomorfos y el comportamiento de escape son pobremente conocidos. Se caracteriza el uso del microhábitat y las tácticas de escape, se determinan las relaciones entre la distancia de iniciación de escape y los dos factores de riesgo (habituación a presencia humana y altura a la que se posan) de Anolis lineatopus, una lagartija que habita en troncos-tierra y A. grahami, una lagartija de troncos-partes más altas. Los tamaños de muestra para A. lineatopus y A. grahami fueron: 214 y 93, para uso del microhábitat y destinos de escape 74, para presencia humana 34 y para perchas altas 125 y 34. Las dos especies se presentan en microhábitats similares y mostraron tácticas de escape parecidas, pero exhibieron diferencias claves esperadas para sus ecomorfos. Ambas especies fueron vistas con frecuencia en el suelo y en los árboles, pero A. lineatopus fue encontrada más frecuentemente en el suelo y debajo de A. grahami. Ambas especies escaparon del suelo a esconderse en los árboles y huían con igual frecuencia sin cambiar de dirección. La frecuencia de huir hacia arriba fue mayor para A grahami. Ambas especies mostraron habituación al tener distancias más cortas de iniciación de escape en zonas con exposición frecuente a la gente y la distancia de iniciación de escape incrementa cuando la altura de la percha disminuye, porque las lagartijas tienden a subir más al estar fuera de nuestro alcance cuando se posan en la parte baja. La relación entre la distancia de iniciación de escape y altura de la percha puede aplicar a otros ecomorfos de Anolis que huyen hacia arriba cuando están posados en las partes bajas de los árboles.
Palabras clave: ecomorfos, tácticas de escape, distancia de iniciación de escape, habituación, altura de la percha, Polychrotidae, Squamata.
Escape behavior has been studied extensively in lizards, effects of predation risk factors and costs of fleeing have been examined more thoroughly in lizards than in any other taxonomic group. Surprisingly little information is available about effects of factors that affect predation risk on the escape behavior of anoles. However, flight initiation distance (distance between the prey and an approaching predator when the prey begins to flee) has been shown to increase with several aspects of predation risk in one or more species of Anolis. Escape strategies and responses to predation risk may be affected by microhabitat use (Losos & Irschick 1996, Regalado 1998, Irschick & Losos 1999, Schneider et al. 2000, Irschick et al. 2005, Lattanzio 2009), which differ greatly among the nearly 400 species of anoles (Losos 2009).
Ecomorphs of Anolis lizards in the Greater Antilles differ in microhabitat use and in aspects of morphology that affect locomotion in their respective microhabitats (Williams 1972, 1983, Losos 2009). Such differences might affect methods and destinations of escape. Comparative studies to assess similarities and differences in escape behavior among ecomorphs have only recently begun (Cooper 2006a, Vanhooydonck et al. 2007), but few species have been studied and many risk factors that affect escape have been studied little or not at all.
Among risk factors that have been studied in anoles are conspicuousness/detectability, temperature, previous attack by a predator, perch height and possibly habituation to predators. Flight initiation distance was longer in A. cristatellus (Duméril & Bibron 1837), A. krugi (Peters 1876) and A. pulchellus (Duméril & Bibron 1837) that were in the open, fully exposed to the experimenter’s view, than in individuals that were partially concealed (Cooper 2006a), suggesting that detectability influences their escape decisions. Heatwole (1968) attributed the shorter flight initiation distance of A. stratulus (Cope 1862) than A. cristatellus to greater crypsis in the former. This interpretation is consistent with the finding that A. stratulus had the shortest flight initiation distance and was the least conspicuous among seven species of anoles in Puerto Rico [the twig anole A. valencianni (Duméril & Bibron 1837) was not studied, Cooper 2006a]. In Anolis lineatopus (Gray 1840) and A. gundlachi (Peters 1876) flight initiation distance decreases as temperature increases (Rand 1964, Cooper 2006a). This relationship reflects the slower maximum running speed and therefore, reduced escape ability of lizards at lower body temperature (Bennett 1980). Rapid learning about the dangerousness of a predator is implied by an increase in flight initiation distance by two Cuban anoles, A. sagrei (Duméril & Bibron 1837), and A. homolechis (Cope 1864) after they were captured a single time (Regalado 1998).
Perch height is an important risk factor that affected flight initiation distance in six of seven species of Puerto Rican anoles (Cooper 2006a). Flight initiation distance decreased as perch height increased in four arboreal species that escape by climbing out of reach, but increased as perch height increased in A. krugi and A. pulchellus, which are grass-bush anoles that escape by fleeing to the ground when initially perched higher (Cooper 2006a). In the trunk-crown anole A. stratulus perch height did not affect flight initiation distance, which is presumably another indication of its reliance on crypsis (Cooper 2006a). Consistent with the findings for Puerto Rican arboreal lizards, flight initiation distance decreased as perch height increased in the Hispaniolan trunk anole A. distichus (Cope 1862) (Schneider et al. 2000).
]]>Other indications that anoles assess risk in making decisions about antipredatory behavior are that duration of tonic immobility is greater in A. carolinensis (Voigt 1832) when a predator is closer and shorter if a refuge is near (Hennig et al. 1976). A single study failed find an effect of predation risk. Lattanzio (2009) reported that flight initiation distance is unaffected by predator approach speed in A. humilis (Peters 1863) and A. limifrons (Cope 1862). However, the methods were so severely flawed that this report should be disregarded. The biggest problem was that effects of occupation of different microhabitats and perch heights were ignored despite their strong effects on escape by anoles and other lizards (Regalado 1998, Schneider et al. 2000, Stankowich & Blumstein 2005, Cooper 2006b, Cooper & Wilson 2007, Vanhooydonck et al. 2007). Furthermore, in the only other anole for which the relationship has been studied, flight initiation distance is substantially greater for fast than slow approaches in A. lineatopus (Cooper 2006b, unpublished data).
I compare aspects of escape behavior in the trunk-ground anole A. lineatopus and the trunkcrown anole A. grahami (Gray 1845) from syntopic populations near Kingston, Jamaica. I examine differences in substrate use related to the ecomorphs, compare escape destinations and movements for lizards initially on the ground or on trees. In some lizards, flight initiation distance is shorter in areas where lizards are habituated to human presence (Burger & Gochfeld 1990, Cooper et al. 2003a, Cooper & Whiting 2007a, Cooper 2009a). I report the relationship between habituation to people and flight initiation distance for both anoles. I also examined the difference in perch height between species and ascertained the relationship between perch height and flight initiation distance in each species.
Materials and methods
Study site and general procedures: All data were collected on the campus of the University of the West Indies at Mona, Jamaica, during late March and the first half of April of 2005. The frequency of human presence and the abundance of people when present varied greatly across the campus. This way, three different areas were selected. In central areas near classrooms and other centers of student activity, people were usually present in hours when classes were taught and numbers of individuals present at a given time was higher than in other areas of campus. In peripheral areas adjacent to buildings, frequency and abundance of human presence was substantially lower, but vegetation, buildings and sidewalks were similar to those in classroom areas. In agricultural fields containing scattered trees, human beings were much less frequently present and buildings and sidewalks were absent.
To study escape responses, I simulated an approaching predator by walking toward adult lizards until they fled. Flight initiation distance did not differ between sexes in A. lineatopus (Cooper, unpublished data); sex was not recorded for A. grahami. Use of experimenters as predators is a widely accepted and successful method for studying escape behavior in the field (Stankowich & Blumstein 2005, Cooper 2009b,c). Using this method, approaches can be controlled and standardized much more easily than can approaches by real predators or mechanical models of predators. Real predators may be difficult to obtain, train and control, and mechanical models are difficult to maneuver in complex habitats. A strong indication of the validity of this method is that predictions of optimal escape theory about flight initiation distance are routinely confirmed for factors affecting both predation risk and costs of fleeing in studies using human simulated predators (Cooper 1999, 2000a, 2009a,b, Martín & López 2003, Cooper et al. 2003a).
Because human beings are not natural predators of anoles, escape responses specific to certain natural predators might not be elicited by human beings (Leal & Rodriguez-Robles 1997, Stuart-Fox et al. 2006). However, human approach readily elicits escape by anoles (Rand 1964, Heatwole 1968) and is the standard technique for studying their escape behavior in the field (Regalado 1998, Schneider et al. 2000, Cooper 2006a, Vanhooydonck et al. 2007).
]]> Data were collected only on warm, sunny days when lizards were fully active. I located lizards by searching visually while walking slowly through the UWI campus. When I detected a lizard, I very slowly moved into a position that allowed the lizard an unobstructed view of me standing 8-12m from it. I stopped walking for approximately 10 seconds and then approached the lizard directly, i.e., on a line leading to it. Prior to data collection I practiced a slow approach speed (60.6+3.3m/min, variability is reported throughout as SE). At intervals throughout the study I practiced the approach speed to prevent drift over time. I used a similar gait in all approaches and practiced the approach speeds 10 times for each speed used and each day tested the speeds to prevent drift. I approached at constant speed until a lizard began to flee. Except when the goal was to determine the directions moved and destinations of escaping lizards, then I stopped immediately and recorded the flight initiation distance to the nearest 0.1m. When lizards were on trees, flight initiation distance was measured as distance between the experimenter and the tree trunk on the ground.In each data set a particular lizard was tested only once, but the same lizard may have been included in more than one study. I avoided pseudoreplication by searching each area only once in a single direction during a set of observations and by moving at least 10m between trials while noting the position of the lizard tested previously to avoid approaching it again.
Designs and analyses: Effects of microhabitat use on direction of escape movements and eventual refuge sought were assessed within and between the two species of anoles. Upon sighting a lizard, I recorded its initial microhabitat and then approached it until it entered a refuge. I then recorded the type of refuge used. For lizards that were on trees initially, I recorded the upward and downward movements of each individual during its escape run and whether its final destination was on the side of the tree trunk visible to me or on the far side where the lizard was hidden.
The effect of frequency of human presence on flight initiation distance was studied both to assess the influence of habituation to potential predators that do not attack on escape by anoles and for possible use as a covariate in a study of effects of perch height. In the study of habituation, I recorded the flight initiation distance and location of each lizard tested, as well as the location (in relation to human presence). I observed no attempts by people to capture lizards. No differences in distance to nearest refuge were apparent among the three areas because lizards were on or near trees in all of them. Observations were restricted to lizards that were not interacting socially with other lizards, were no closer than 5m from the nearest visible lizard and were no closer than 10m from the nearest person. No trials were conducted when people were walking within 30m of the focal lizard. To examine effects of perch height on flight initiation distance, I approached lizards on tree trunks and recorded both variables to the nearest 0.1m.
To assess differences in frequency of occupation of microhabitats, refuges and escape directions, I conducted Chi-square analyses. Effects of risk factors on escape were assessed using parametric analyses performed with Statistica. Variances were tested for homogeneity using Levene’s tests. If variances were heterogenous, data were logarithmically transformed and the transformed data again were tested for heterogeneity. When variances remained significantly heterogeneous after transformation, Kruskal-Wallis nonparametric ANOVA was used. When main effects were significant, nonparametric paired comparisons were made using procedures described by Zar (1996).
Significance tests were two-tailed, with α=0.05. Effect sizes of human abundance and perch height were estimated. For analyses of variance, the effect sizes reported for significant effects is as η2 (Cohen 1992), which has range 0-1, and may be interpreted similarly to R2 (Pierce et al. 2004). For regression, the effect size is R2, which can readily be converted to r.
]]> ResultsEscape destination and movement directions: Anolis lineatopus. Lizards were equally often on trees and bushes (n=113, 53%) as on ground or low objects (<0.3m) such as stones, litter and boards (n=101, 47%) and this difference was not significant (Χ2 1=0.68, p>0.10). When lizards initially on the ground were approached until they took refuge, slightly over half of them fled to a tree or bush (n=53) and over one third hid beneath rocks or other objects on the ground (n=37); the remainder hid on the far side of rocks or other objects (n=11) but remained on the surface (Fig. 1). Proportions of lizards on the ground that used the three types of refuges differed significantly (Χ2 2=26.69, p <0.001). Significantly lower proportions of lizards hid on the far side of rocks or other low objects than fled to trees or bushes (Χ2 2=27.56, p<0.001) or hid beneath objects (Χ2 1=14.08, p<0.001). The proportion that fled to trees or bushes was somewhat greater than the proportion that hid under objects, but the difference was not significant (Χ2 1=2.84, p=0.09).
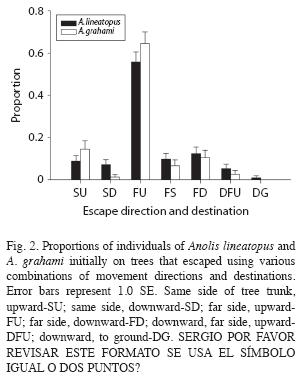
Escape by A. lineatopus initially on trees exhibited considerable variation in direction and resort to hiding (Fig. 2). Over four fifths of lizards (95 of 113) eventually fled out of sight on the far sides of tree trunks, significantly more than expected by random escape with respect to side of the trunk (Χ2 1=77.92, p<0.001). Nearly two thirds of these lizards fled upward (n=79). This percentage was significantly higher that the percentage of lizards that fled downward or at the same height (Χ2 1=11.04, p<0.001). Most lizards (n=107, 95%) exhibited simple escape behavior with movement directly to a destination where the lizard stopped, but a few lizards made more complex movements in which they moved downward initially while visible on the trunk, then moved to the far side of the trunk and fled upward (n=6, 5%). Because the predator did not pursue lizards when they fled, simple escape maneuvers were significantly more frequent than the more complex ones (Χ2 1=88.30, p<0.001). Only one lizard left a tree to hide in debris on the ground.
Anolis grahami. When initially sighted, 14 lizards (15%) were on the ground or low objects; three were on piles of logs (3%) and 76 were on trees (82%). A significantly higher percentage occurred on trees than elsewhere (Χ2 1=37.43, p<0.001). Among lizards on the ground, about four-fifths fled to trees or bushes when approached and one fifth hid under debris. All three lizards on logs moved to adjacent logs (n=2) or hid beneath the log they occupied.
Anolis grahami initially on trees (n=76) all remained there after escape movements, but the directions and eventual destinations of escape runs (climbs) were diverse (Fig. 2). Eighty-four percent of lizards on trees eventually fled out of sight on the far sides of trunks (n=64), significantly more than expected by random escape with respect to side of the trunk (Χ2 1=35.58, p<0.001). Sixty-one of 76 lizards (80%) fled upward immediately; one (1%) did so after initially moving downward. A significantly greater percentage of lizards fled upward than fled downward or stayed at the same height (Χ2 1=27.84, p<0.001). A large majority (97%) of 94 lizards exhibited simple escape behavior with movement directly to a destination where the lizard stopped, but three lizards made more complex movements in which they moved downward initially while visible on the trunk, moved to the far side of the trunk and then fled upward (3%). In this experiment in which the predator did not pursue lizards once they began to flee, simple escape maneuvers were significantly more frequent than more complex ones (Χ2 1=68.21, p<0.001).
The proportion of lizards initially on the ground was significantly greater for A. lineatopus than for A. grahami (Χ2 1=22.91, p<0.001) but the escape repertoires of these species were similar. The proportion of lizards detected on the ground that fled to trees was somewhat greater in A. grahami than A. lineatopus, but the difference was not significant (Χ2 1=2.51, p>0.10). Neither did the proportion of lizards initially on trees that fled to the far sides of tree trunks differ between species (Χ2 1=0.03, p>0.10). A slightly higher proportion of A. grahami than A. lineatopus on trees initially fled upward (Χ2 1=5.03, p=0.025). Proportions of lizards on trees that escaped without changing the direction of movement from downward to upward did not differ significantly between species (Χ2 1=0.80, p>0.10).
]]>Exposure to human beings: For A. lineatopus perched below 1.0m, flight initiation distance appeared to differ strongly among the three areas differing in frequency of exposure to people (F2,71=33.32, p<1.1x10-6, n=74) but variances were significantly heterogeneous (Levene’s F2,71=6.09, p=0.0036) and could not be rendered homogeneous by logarithmic transformation. Using a nonparametric Kruskal-Wallis ANOVA, flight initiation distance differed significantly among the three campus areas (Χ2 2=32.00, p<1.0x10-4, n=74). Flight initiation distance differing significantly among all pairs of groups (sample sizes were 23 in the area with few people, 24 for intermediate human presence and 27 for the area with greatest human presence), being longest where few people occurred and shortest where lizards were exposed to people most frequently (p<0.001 each, Fig. 3).
Flight initiation distance in A. grahami was shorter in the area with the highest frequency of human presence (0.5+0.2m, n=18) than in the area of intermediate frequency of human presence (1.2+0.3m, n=14). Using raw data this difference was significant (F1,29=10.02, p =0.0036) but variances differed significantly (Levene’s F1,29=5.24, p=0.030). Following square root transformation, variances were homogeneous (Levene’s F1,29=3.77, p=0.062). Using transformed data, flight initiation distance was significantly longer where people were present less frequently (F1,29=7.79, p=0.0092). No interspecific comparison of the effect of human abundance on flight initiation distance is reported due to the absence of data for A. grahami in areas with few people and the extremely small sample sizes of A. graham perched below 1.0m.
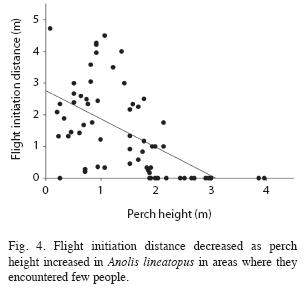
For A. lineatopus approached slowly in areas with few people, flight initiation distance decreased significantly as perch height increased (F1,62=32.67, p<1.0x10-6, R2=0.35, n=64, Fig. 4). In the regression equation FID=- 0.59PH+2.82m, where PH is perch height, the intercept is significantly greater than zero (t62=10.15, p<1.0x10-4). In similar regressions in the areas of intermediate and high human presence, R2=0.57 and R2=0.65, respectively).
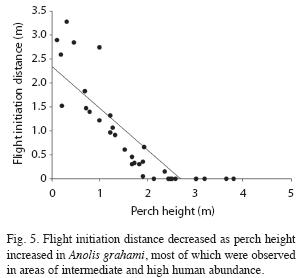
In A. grahami flight initiation distance decreased as perch height increased in areas of high human presence (F1,16=42.69, p=7.0x10-6, R2=0.73, n=18). The regression equation is FID=-0.85PH+1.78m, the intercept is significantly greater than zero (t16=8.52, p<1.0x10-4). Flight initiation distance also decreased as perch height increased in areas of low-intermediate human presence (F1,14=57.44, p=3.0x10-6, R2=0.80, n=16). The regression equation is FID=-0.90PH+2.79m; the intercept is significantly greater than zero (t14=11.24, p<1.0x10-4). Disregarding human presence, the correlation between flight initiation distance and perch height is -0.85 (Fig. 5) for all data and -0.90 for perch heights less than 3.0m. ]]>
Discussion
Microhabitats and escape directions and destinations: The higher proportion of A. grahami than A. lineatopus on trees matches expected habitat use for trunk-crown and trunkground anoles. Frequent observation of A. graham on ground may reflect separation between trees on campus. The higher perch height by A. grahami than A. lineatopus is also expected for a trunk-crown versus trunk-ground anole. The species had almost identical escape repertoires. The only qualitative difference was that all A. grahami remained on trees, but one individual of A. lineatopus perched low on a tree fled down to the ground and hid beneath debris.
Escape movements of the two species were similar for lizards occupying similar perches. The probabilities of taking refuge using objects, debris or fleeing to trees were equal for both species when on ground. When on trees both fled to far sides of trunks, a common anole escape tactic (Regalado 1998, Schneider et al. 2000) with equal probability. Both usually moved to the far side and up or down although some individuals remained visible while fleeing. Variability of change in perch height when on the far side of the trunk may be a strategy to decrease predictability of the location of hidden anoles. Another deceptive tactic in both species is moving down while visible but then moving to the far side of the trunk and climbing upward once out of view. For lizards on trees the proportions of lizards that fled up or down without changing vertical direction also was similar in the two species. The only quantitative difference in escape direction was that a slightly higher proportion of A. graham than A. lineatopus fled upward. It remains to be seen whether this is a consistent difference between trunk-ground and trunk-crown ecomorphs and might be a consequence of differences in perch height.
]]>Human presence: The shorter flight initiation distance by both species in areas of highest exposure of lizards to people is a novel finding for anoles and is consistent with reports of decreased flight initiation distance where diverse lizards are frequently exposed to people (phrynosomatids- Cooper 2009a, iguanids-Burger & Gochfeld 1990, Cordylids- Cooper & Whiting 2007b, teiids-Cooper et al. 2003a). Decreased flight initiation distance in the cited studies has been attributed to habituation. Alternatively, the risk allocation hypothesis predicts that prey decrease allocation of effort to antipredatory behavior as the presence/ abundance of predators increases (Lima & Bednekoff 1999) are often present. Habituation is more likely to apply in this case because people rarely if ever attack the lizards on the campus of the University of the West Indies at Mona. Taken together with the increase in flight initiation distance after anoles were captured and handled a single time, the effect of frequency of human presence on flight initiation distance suggests that anoles adjust their assessment of risk to their experience, whether positive or negative, with potential predators. The anoles in this study exhibited escape behaviors that were similar to those by the same species that I observed casually in other locations, suggesting that exposure to people on campus has altered escape quantitatively, but not qualitatively.
Perch height and escape by anole ecomorphs: In both anoles flight initiation distance decreased as perch height increased, as in other anoles and other lizards that escape up trees lizards (Phrynosomatidae-Cooper 2009a, Scincidae-Cooper 1997). Perch height is a special case of distance to refuge, which strongly affects escape decisions: flight initiation distance increases as distance to refuge increases (e.g., Iguanidae-Cooper 2003, Phrynosomatidae- Cooper 2000b, Scincidae-Cooper 1997, A. lineatopus-Cooper in review). The relationship between distance that must be climbed to reach safety and flight initiation distance may be widespread in anoles because perch height and flight initiation distance were negatively correlated in a phylogenetic analysis of 12 anole species (Vanhooydonck et al. 2007).
For species that escape by climbing trees beyond reach of terrestrial predators, distance to refuge decreases as perch height increases form ground level to the minimum safe height. Higher, escape is not required. The height above which lizards do not flee is indicated by the intercept of the regression of flight initiation distance on perch height to be about 3m. Although A. lineatopus and A. grahami sometimes escape downward, their predominant escape direction is upward. Their negative relationships between flight initiation distance and perch height are predicted by positive relationships between flight initiation distance and distance to refuge. The same applies to the trunk anole A. distichus (Schneider et al. 2000).
In grass-bush anoles that flee to ground, distance to refuge increases as perch height increases. Thus, increase in flight initiation distance as perch height increased in the grassbush species A. krugi and A. pulchellus (Cooper 2006a) also reflects increasing flight initiation distance as distance to refuge increases. Even in members of more arboreal ecomorphs, flight initiation distance increases as perch height increases when the lizards must escape downward (Regalado 1998). I predict that for lizards within 2-3m of ground on trees flight initiation distance increases as perch height decreases in trunk-ground, trunk-crown, trunk and crown giant ecomorphs, but increases as perch height increases in grass-bush anoles.
The only arboreal anole for which perch height and flight initiation distance were not related was the cryptic A. stratulus (Rand 1964, Cooper 2006a). It may rely more than other anoles on crypsis to avoid detection. Because use of crypsis would shorten flight initiation distance more for lizards at risk when perched low than for lizards perched higher, its effect would be to reduce or eliminate the influence of perch height on flight initiation distance.
The cryptic twig anole A. valencienni had the shortest flight initiation distance among 12 species studied by Vanhooydonck et al. (2007). Because twig anoles appear to rely strongly on crypsis maintained by immobility to avoid detection, I predict that the relationship between perch height and flight initiation is reduced or absent in twig anoles.
]]>Ecomorphs and flight initiation distance: In two studies with overlap of species, trunkcrown anoles permitted closer approach than trunk-ground and grass-bush anoles. The sole exception was A. cooki, a Puerto Rican trunkground anole with flight initiation distance similar to that of the local trunk-crown anoles (Cooper 2006a). This discrepancy is presumably a consequence of a protective effect of branches shielding A. cooki (Grant 1931) from the predator and habituation of some lizards to human presence (Cooper 2006a). The other trunk-ground species perched on bare trunks (Cooper 2006a). Flight initiation distance of a trunk anole was comparable to that of trunkcrown anoles (Cooper 2006a, Vanhooydonck et al. 2007). Data are lacking for crown giants, presumably due to infrequent encounters near the ground. The longer flight initiation distance of trunk-ground and grass-bush anoles than other ecomorphs requires confirmation in other Greater Antillean radiations. It may reflect greater predation for these lizards than more arboreal ecomorphs due to differences in predator suites. For grass-bush anoles, risk may be greater on the ground than for arboreal species when they climb out of reach.
Comparisons among studies and ecomorphs: Flight initiation distances for A. lineatopus are nearly identical in this study (0.83m) and in Vanhooydonck et al. (0.85m, 2007). This is surprising because their approach speed was faster than mine but flight initiation distance increases as approach speed increases in this species (Cooper 2006b). Data were collected from different populations that may differ in exposure to predation and human beings and in other ways affecting escape. Vanhooydonck et al. (2007) reported shorter flight initiation distance (0.60m) than I observed for A. graham (0.99m) despite similarity in perch height and their greater approach speed. I approached all individuals in the same way, but Vanhooydonck et al. (2007) held a clipboard attached to a 2m pole at the level of the lizard when approaching individuals perched above their heads. The effect of this difference in methods is unknown. Differences between populations in variables such as exposure to people and leaf cover on tree trunks might account for the difference. Differential response to individual investigators is another possibility (Cooper et al. 2003b). Quantitative comparisons of flight initiation distance between studies involving different populations and predators must be made cautiously.
Comparisons within and among ecomorphs may be affected by differences in local conditions, and flight initiation distance may or may not be affected in parallel ways in independent anole radiations on the four islands of the Greater Antilles. Phylogenetically informed analyses such as those conducted by Vanhooydonck et al. (2007) are essential for understanding relationships between flight initiation distance and ecomorphs. In future studies planned to compare escape by ecomorphs across islands, information about habitat variables and predator suites not normally collected in studies of escape behavior would be very useful.
Acknowledgments
I am grateful to Byron Wilson and Klaus Wolf for hospitality and logistical assistance.
References
Bennett, A.F. 1980. Thermal dependence of lizard behavior. Anim. Behav. 28: 752-762. [ Links ]
]]>Burger, J. & M. Gochfeld. 1990. Risk discrimination of direct versus tangential approach by basking black iguana (Ctenosaura similis): variation as a function of human exposure. J. Comp. Psychol. 104: 388-394. [ Links ]
Cohen, J. 1992. A power primer. Psychol. Bull. 112: 155- 159. [ Links ]
Cooper, W.E. Jr. 1997. Escape by a refuging prey, the broad-headed skink (Eumeces laticeps). Can. J. Zool. 75: 943-947. [ Links ]
Cooper, W.E. Jr. 1999. Tradeoffs between courtship, fighting, and antipredatory behavior by a lizard, Eumeces laticeps. Behav. Ecol. Sociobiol. 47: 54-59. [ Links ]
Cooper, W.E. Jr. 2000a. Tradeoffs between predation risk and feeding in a lizard, the broad-headed skink (Eumeces laticeps). Behaviour 137: 1175-1189.
Cooper, W.E. Jr. 2000b. Effect of temperature on escape behaviour by an ectothermic vertebrate, the keeled earless lizard (Holbrookia propinqua). Behaviour 137: 1299-1315. [ Links ] [ Links ]
Cooper, W.E. Jr. 2006a. Risk factors affecting escape behavior by Puerto Rican Anolis lizards. Can. J. Zool. 84: 495-504. [ Links ]
Cooper, W.E. Jr. 2006b. Dynamic risk assessment: prey rapidly adjust flight initiation distance to changes in predator approach speed. Ethology 112: 858-864. [ Links ]
Cooper, W.E. Jr. 2009a. Flight initiation distance decreases during social activity in lizards (Sceloporus virgatus). Behav. Ecol. Sociobiol. 63: 1765-1771. [ Links ]
Cooper, W.E. Jr. 2009b. Fleeing and hiding under simultaneous risks and costs. Behav. Ecol. 20: 665-671. [ Links ] [ Links ]
Cooper, W.E. Jr. Risk factors affecting escape behavior by the Jamaican lizard Anolis lineatopus (Polychrotidae, Squamata). In review. [ Links ]
Cooper, W.E. Jr. & M.J. Whiting. 2007a. Universal optimization of flight initiation distance and habitatdriven variation in escape tactics in a Namibian lizard assemblage. Ethology 113: 661-672. [ Links ]
Cooper, W.E. Jr. & M.J. Whiting. 2007b. Effects of risk on flight initiation distance and escape tactics in two southern African lizard species. Acta Zool. Sin. 53: 446-453. [ Links ]
Cooper, W.E. Jr. & D.S. Wilson. 2007. Sex and social costs of escaping in the striped plateau lizard Sceloporus virgatus. Behav. Ecol. 18: 764-768. [ Links ]
Cooper, W.E. Jr., V. Pérez-Mellado, T. Baird, T.A. Baird, J.P. Caldwell & L.J. Vitt. 2003a. Effects of risk, cost, and their interaction on optimal escape by nonrefuging Bonaire whiptail lizards, Cnemidophorus murinus. Behav. Ecol. 14: 288-293. [ Links ]
Cooper, W.E. Jr., J. Martin & P. Lopez. 2003b. Simultaneous risks and differences among individual predators affect refuge use by a lizard, Lacerta monticola. Behaviour 140: 27-41. [ Links ]
Heatwole, H. 1968. Relationship of escape behavior and camouflage in anoline lizards. Copeia: 109-113. [ Links ]
Hennig, C.W., W.P. Dunlap & G.G. Gallup Jr. 1976. The effect of distance between predator and prey and opportunity to escape on tonic immobility in Anolis carolinensis. Psychol. Rec. 26: 313-320. [ Links ]
Irschick, D. & J. Losos. 1999. Do lizards avoid habitats in which performance is submaximal? The relationship between sprinting capabilities and structural habitat use in Caribbean anoles. Am. Nat. 154: 293-305. [ Links ]
Irschick, D.J., E. Carlisle, J. Elstrott, M. Ramos, C. Buckley, B. Vanhooydonck, J. Meyers & A. Herrel. 2005. A comparison of habitat use, morphology, clinging performance and escape behaviour among two divergent green anole lizard (Anolis carolinensis) populations. Biol. J. Linn. Soc. 85: 223-234. [ Links ]
Lattanzio, M. 2009. Escape tactic plasticity of two sympatric Norops (Beta Anolis) species in Northeast Costa Rica. Amph.-Rept. 30: 1-6. [ Links ]
Leal, M. & J.A. Rodriguez-Robles. 1997. Signalling displays during predator-prey interactions in a Puerto Rican anole, Anolis cristatellus. Anim. Behav. 54: 1147-1154. [ Links ]
Lima, S.L. & P.A. Bednekoff. 1999. Temporal variation in danger drives antipredator behavior: the predation risk allocation hypothesis. Am. Nat. 153: 649-659. [ Links ]
Losos, J.B. 2009. Lizards in an evolutionary tree: Ecology and adaptive radiation of anoles. University of California, Berkeley, California, USA. [ Links ]
Losos, J.B. & D.J. Irschick. 1996. The effect of perch diameter on escape behaviour of Anolis lizards: Laboratory predictions and field tests. Anim. Behav. 51: 593-602. [ Links ]
Martín, J. & P. López. 2003. Changes in the escape responses of the lizard Acanthodactylus erythrurus under persistent predatory attacks. Copeia: 408-413. [ Links ]
Pierce, C.A., C.A. Bliock & H. Aguinas. 2004. Cautionary note on reporting eta-squared values from multifactor ANOVA designs. Educ. Psychol. Meas. 64: 916-924. [ Links ]
Rand, A.S. 1964. Inverse relationship between temperature and shyness in the lizard Anolis lineatopus. Ecology 45: 863-864. [ Links ]
Regalado, R. 1998. Approach distance and escape behavior of three species of Cuban Anolis (Squamata, Polychrotidae). Carib. J. Sci. 34: 211-217. [ Links ]
Schneider, K.R., J.S. Parmerlee Jr. & R. Powell. 2000. Escape behavior of Anolis lizards from the Sierra de Baoruco, Hispaniola. Carib. J. Sci. 36: 321-323. [ Links ]
Stankowich, T. & D.T. Blumstein. 2005. Fear in animals: a meta-analysis and review of risk assessment. Proc. Roy. Soc. Lond., Ser. B, Biol. Sci. 272: 2627-2634. [ Links ]
Stuart-Fox, D., M.J. Whiting & A. Mousalli. 2006. Camouflage and colour change: antipredator responses to bird and snake predators across multiple populations in a dwarf chameleon. Biol. J. Linn. Soc. 88: 437-446. [ Links ]
Vanhooydonck, B., A. Herrel & D. Irschick. 2007. Determinants of sexual differences in escape behavior in lizards of the genus Anolis: a comparative approach. Integr. Compar. Biol. 47: 200-210. [ Links ]
Williams, E.E. 1972. The origin of faunas. Evolution of lizard congeners in a complex island fauna: a trial analysis. Evol. Biol. 6: 47-89. [ Links ]
Williams, E.E. 1983. Ecomorphs, faunas, island size, and diverse end points in island radiations of Anolis, p. 326-370. In R.B. Huey, E.R. Pianka & T.W. Schoener (eds.). Lizard Ecology: Studies of a Model Organism. Harvard University, Cambridge, Massachusetts, USA. [ Links ]
Zar, J.H. 1996. Biostatistical analysis. Prentice Hall, Upper Saddle River, New Jersey, USA. [ Links ]
Correspondencia a: William E. Cooper, Jr. Department of Biology, Indiana University Purdue University Fort Wayne, Fort Wayne, IN 46805, USA; cooperw@ipfw.edu.
Received 23-XI-2009. Corrected 10-VI-2010. Accepted 15-VII-2010.
]]>