Caracterización florística y estructural de tres fragmentos boscosos secundarios en Cartago, Costa Rica
Carlos O. Morales
Escuela de Biología, Universidad de Costa Rica, 11501-2060, San José, Costa Rica; oldem@biologia.ucr.ac.cr
Abstract: Floristic and structural characterization of three secondary forest fragments in Costa Rica. Observations on successional development and floristic composition were made in 25 plots (10x2 m) established in three contiguous secondary forest fragments of the Lankester Botanical Garden, Costa Rica. Human activities favored forest regeneration there by 1) protecting an abandoned farm and pasture area since 1970, and 2) planting and propagating plant species. Planting successfully growing native and introduced species can accelerate the succession process, avoid dominance of pioneer or invasive species, and increase species diversity faster than in common successional processes. Natural diversification slowed down as natural vegetation surrounding the secondary fragments was disappearing, with a consequent reduction or lost of seed input and other reproductive sources, such as spores and stalks. introduced plant species became naturalized in these forests, their reproduction is successful, and some of them are invasive. This study proposes general paths to help Neotropical forest restoration processes by combining natural succession and planting of native species. Rev. Biol. Trop. 57 (Suppl. 1): 69-82. Epub 2009 November 30.
Key words: secondary forest fragments, forest restoration, introduced plant species, natural succession, Lankester Botanical Garden, Costa Rica.
Grandes áreas de los trópicos han sido convertidas en pastizales para ganado. Con frecuencia, los pastizales abandonados no se transforman de nuevo en bosques. Se han estudiado las barreras que impiden la regeneración de bosques en bajuras húmedas. En pastizales fuertemente degradados de la Sierra Neva número de semillas, depredación de semillas, competencia con gramíneas, microclima hostil, condiciones de suelo inapropiadas e incendios (Aide & Cavelier 1994). En Panamá, terrenos agrícolas abandonados, antes cubiertos por bosques lluviosos, son invadidos por el pasto alóctono Saccharum spontaneum L. (Poaceae). Aquí, cuatro barreras para la regeneración natural son: competencia de Saccharum, limitada dispersión de semillas, incendios y suelos pobres en nutrientes. Hooper (2005) determinó que los incendios reducen significativamente la riqueza de especies de plantas durante la regeneración e inhiben la germinación de la mayoría de semillas, mientras que la competen cia de la gramínea reduce significativamente el crecimiento de plántulas nativas. Como otros, este estudio sugiere que facilitar la regenerada, Colombia, esas barreras fueron un bajo ción natural puede ser una opción de manejo factible y de bajo costo.
]]> La tala y la fragmentación de bosques están provocando cambios ecológicos complejos donde antes hubo masas continuas de selvas tropicales. La fragmentación de bosques puede reducir drásticamente la regeneración, en gran parte porque en los bordes de los fragmentos boscosos es más difícil el establecimiento de plántulas de árboles tolerantes a la sombra o característicos de bosques maduros. Sin embargo, en la Amazonia central se ha observado que el cierre del borde por regeneración del bosque y crecimiento del sotobosque del bosque primario puede amortiguar el aumento de temperatura y la reducción de humedad inducidos por el borde (Benítez-Malvido 1998, Gascon et al. 2000).A inicios de la década de 1970, algunos terrenos de cultivo y pastoreo de la finca El Silvestre, en Cartago, Costa Rica, que perteneció al caficultor y naturalista inglés Charles H. Lankester (1879-1969), fueron abandonados, de modo que allí se inició un proceso de regeneración que dura hasta el tiempo actual. En un plano de la finca, realizado por R.L. Rodríguez en 1973 (inéd.), basado en una fotografía aérea de 1971, se observan todavía los campos de cultivo y de pastoreo abandonados. En 1973, parte de esa finca se transformó en el Jardín Botánico Lankester. Los fragmentos secundarios existentes allí, que cubren cuatro a cinco de las casi 11 has. del jardín, representan una de las pocas áreas boscosas protegidas del área del Valle Central comprendida entre Paraíso de Cartago y la ciudad de San José (ca. 25 km de distancia).
Los fragmentos regenerados en el Jardín Lankester son el producto no sólo de un proceso de sucesión natural, sino también de la intervención humana, porque allí se cultivaron múltiples especies de plantas terrestres y epífitas, nativas e introducidas. Varios aspectos de la regeneración de estos fragmentos se desconocen, porque lamentablemente no se hicieron observaciones en las primeras etapas del proceso.
Una de las alternativas a la destrucción de los bosques tropicales es la recuperación de fragmentos boscosos en áreas de cultivo y pastoreo abandonadas y en matorrales, especialmente donde los suelos se han compactado, erosionado y empobrecido. Por ello, se considera relevante el establecimiento de pequeñas áreas boscosas en cerros, orillas de ríos y terrenos abandonados, tal como fue propuesto originalmente en Costa Rica por Fournier & Herrera (1979), no solamente en áreas rurales, sino también en áreas suburbanas. El interés oficial del Estado en el cuidado del ambiente tiene que coincidir con el interés privado de los propietarios de tierras, quienes estén dispuestos a dedicar parte de sus propiedades a la regeneración de bosques. En este contexto se justifica la investigación de los fragmentos secundarios del Jardín Botánico Lankester.
A partir de los resultados de este proyecto y de otros que podrán realizarse, se espera tener un punto de partida para establecer lineamientos sobre recuperación de bosques, lo que podrá mejorar la calidad ambiental, tanto en zonas rurales como suburbanas. El impacto consiste en señalar pautas para la recuperación de bosques con intervención humana. El objetivo principal del presente trabajo es estudiar la composición florística de los fragmentos boscosos secundarios del Jardín Botánico Lankester. Aquí es importante aclarar que se trata de un estudio taxonómico, más que ecológico. Los objetivos específicos son: 1) Comparar los resultados de este estudio con los de otros estudios sobre recuperación de bosques; 2) Evaluar el papel humano en el desarrollo de los fragmentos secundarios del Jardín Botánico Lankester, y 3) Proponer ideas prácticas para la recuperación de fragmentos boscosos, especial-mente en el Valle Central de Costa Rica.
Materiales y métodos
Área de estudio: Se ubica en Cartago, cantón Cartago, distrito Dulce Nombre, localidad Cóncavas, aprox. 9º51N, 83º53W, a ca. 1 350 m de altitud, en la zona de vida bosque premontano húmedo (Holdridge 1987), con una precipitación anual entre 1 000 y 1 300 mm y una temperatura diurna de 18º a 24º C.
Métodos: En tres fragmentos secundarios contiguos, sin barreras considerables, que suman unas 5 has., se marcaron 25 parcelas de 10x2 m. En cada fragmento las parcelas se ubicaron equidistantes, separadas al menos 5 m de distancia, orientadas perpendicularmente respecto a la mayor longitud del bosque. Se analizaron cinco parcelas en el área más reciente y más pequeña (Bosque Nuevo, ca. 1 ha., ver adelante), diez parcelas en el área central del Jardín (Bosque intermedio) y diez en el área periférica sudoeste (Bosque Periférico). En estas parcelas se cuantificaron y determinaron todas las especies posibles de plantas vasculares, incluso todas las familias y los géneros de plántulas que pudieron ser determinados. Además, se midió el diámetro a altura de pecho (DAP) y se midió o calculó la altura de las plantas leñosas (≥1 m de altura); para esto se colocó un tallo del bambú Phyllostachys aurea Carrière ex Rivière & C. Rivière de 4 m de largo en la base de cada planta >2 m; cada medio metro del tallo de bambú se marcó con cinta de colores. En plantas con más de 4 m de altura se hizo un cálculo de altura a distancia por comparación con el tallo de bambú. Aunque el área de parcelas es pequeña, se hicieron múltiples recorridos para completar el inventario florístico abarcando toda el área de los fragmentos. Se determinaron las proporciones de especies nativas y especies introducidas, así como de especies regeneradas y especies cultivadas, para describir y comparar florísticamente los tres fragmentos boscosos.
Resultados
I. Descripción de los bosques observados
]]> En el Jardín Botánico Lankester hallamos tres fragmentos boscosos secundarios, que son contiguos pero se distinguen tanto por su ubicación como por su desarrollo estructural. Por razones de comodidad y conveniencia, se han nombrado como sigue: 1) Bosque intermedio, 2) Bosque Periférico, 3) Bosque Nuevo.I.1. Bosque Intermedio
Se halla entre las colecciones de suculentas y bromeliáceas y el Bosque Nuevo, con una ubicación aproximadamente central en los terrenos del jardín. El árbol más abundante allí es Eugenia cartagensis O. Berg. Es un fragmento dominado por arbustos y árboles pequeños (3 a 7 m de alto), con algunos árboles medianos dispersos (15 a 20 m).
Un aspecto crucial en el desarrollo de este bosque es que allí se cultivaron plantas terrestres y epífitas. Así, en la recuperación del Bosque intermedio se evidencia un fuerte componente antropógeno. Géneros herbáceos y arbustivos como Dracaena (tres especies), Stromanthe, Cordyline, Crinum, Eucharis, y especies arbóreas como Grevillea robusta A. Cunn., Syzygium jambos (L.) Alston y Tabebuia ipe (Mart. ex K. Schum.) Standl., entre otras, son elementos alóctonos adaptados y naturalizados en el fragmento secundario. Otros géneros arbóreos fueron traídos de otras zonas del país y se han adaptado en esta área; tal es el caso de Pachira aquatica Aubl., especie que se regenera exitosamente, y Coussapoa sp. Algunas especies arbóreas introducidas se han adaptado bien y hasta se regeneran en este fragmento. Los casos más notables son Fraxinus uhdei (Wenz.) Lingelsh., Grevillea robusta, Tabebuia ipe y Syzygium jambos.
Especies más comunes en este fragmento
Arbóreas y arbustivas: Chamaedorea costaricana Oerst., Crossopetalum tonduzii (Loes.) Lundell, Croton hoffmannii Müll. Arg., Dracaena deremensis Engl., Dracaena fragrans (L.) Ker Gawl., Eriobotrya japonica (Thunb.) Lindl., Eugenia cartagensis, Grevillea robusta, Piper bredemeyeri Jacq., Syzygium jambos, Tabebuia ipe, Xylosma velutina (Tul.) Triana & Planch.
Trepadoras: Beaumontia grandiflora Wall., Monstera adansonii Schott, Paullinia sp., Philodendron spp.
Herbáceas: Aechmea sp., Anthurium sp., Calathea sp., Heliconia tortuosa Griggs, Impatiens walleriana Hook. f., Molineria capitulata (Lour.) Herb., Monstera deliciosa Liebm., Stromanthe stromanthoides (J.F. Macbr.) L. Andersson, Tradescantia zebrina hort. ex Bosse, Xiphidium caeruleum Aubl.
]]> Epífitas: La mayoría son cultivadas, especialmente Bromeliáceas y Orquidáceas. Algunas Bromeliáceas se reproducen vegetativamente con gran éxito; tres de ellas crecen bien tanto terrestres como epífitas. Aquí están representados los géneros Aechmea, Billbergia, Cryptanthus, Guzmania, Tillandsia y Vriesea, entre otros menos comunes. Las orquídeas más frecuentes son de Maxillaria, Oncidium, Pleurothallis (s.l.) y Warczewiczella. También se observan plantas de Coelogyne cf. ovalis Lindl. (nativa de Asia), Gongora, Oerstedella, Prosthechea, Stanhopea y Stelis. Entre las Aráceas se observan algunas especies de Anthurium; especialmente A. salvinii Hemsl. Los Pteridófitos (helechos y plantas afines) están representados por Hymenophyllum, Nephrolepis y Pleopeltis, entre otros.Regeneración: En este bosque la regeneración más activa es la de especies cultivadas, tanto terrestres como epífitas (los géneros ya mencionados).
I.2. Bosque periférico
Está ubicado en la periferia sudoeste del jardín. Se trata de un fragmento boscoso secundario con desarrollo atípico. Por un lado, hallamos árboles maduros dispersos, al final de un ciclo de vida de 30 o más años. Así, son elementos de la antigua finca, que existían antes de la fundación del jardín. Por otro lado, vemos un sotobosque que en amplios sectores está claramente dominado por una sola especie herbácea: Dieffenbachia oerstedii Schott (Araceae), que crece con gran densidad y evidentemente dificulta el desarrollo de especies leñosas arbustivas y arborescentes. En este fragmento también es evidente que en el pasado se han cultivado algunas especies, pero en un grado de intervención antropógena mucho menor que en el Bosque intermedio; solamente se destacan dos especies cultivadas: Zamia neurophyllidia D.W. Stev. (Zamiaceae), sin reproducción, y Cryosophila warscewiczii (H. Wendl.) Burret (Arecaceae), con plántulas observadas.
Especies más comunes en este bosque
Arbóreas y arbustivas: Annona cherimola Mill., Cecropia obtusifolia Bertol., Chamaedorea costaricana, Chamaedorea tepejilote Liebm., Coffea arabica L., Croton hoffmannii, Eugenia cartagensis, Hampea appendiculata (Donn. Sm.) Standl., Malpighia glabra L., Myrcia splendens (Sw.) DC., Neea psychotrioides Donn. Sm., Pachira aquatica, Parathesis sp., Quercus oocarpa Liebm., Randia aculeata L., Sapium glandulosum (L.) Morong, Syzygium jambos, Xylosma velutina, Zamia neurophyllidia, Zanthoxylum limoncello Planch. & Oerst. y varias Melastomatáceas indeterminadas.
Trepadoras: Hylocereus sp., Malvaviscus sp., Monstera adansonii, Passiflora biflora Lam., Paullinia sp., Philodendron spp., Syngonium sp.
Herbáceas: Anthurium sp., Calathea sp., Costus spp., Ctenanthe sp., Dieffenbachia oerstedii, Heliconia tortuosa, Impatiens walleriana, Lasiacis sp., Pilea sp., Renealmia cernua (Sw.) J.F. Macbr., Spigelia humboldtiana Cham. & Schltdl., Stromanthe stromanthoides, Tradescantia zebrina.
Epífitas: Bromeliaceae (Aechmea, Tillandsia, Vriesea) y Orchidaceae (Lepanthes, Oerstedella, Oncidium, Prosthechea, Stelis) son frecuentes, pero la diversidad es baja si se compara con la de los bosques más diversos en epífitas, sobre todo los nubosos, que fisonómicamente son similares. Son comunes Anthurium sp., algunos helechos (ej. Hymenophyllum), Peperomia spp. y Columnea sp. Solamente abundan briófitos (musgos y hepáticas foliosas) y líquenes.
Regeneración: En este bosque la regeneración de especies leñosas es abundante; sin embargo, la arácea Dieffenbachia oerstedii crece densamente en amplios sectores del sotobosque, lo que limita y probablemente impide el establecimiento de otras especies.
]]>I.3. Bosque nuevo
Se ubica entre el bosque intermedio y la colección de bambúes del jardín. Se trata de un fragmento evidentemente más joven (20 a 25 años al realizar este estudio; D.E. Mora, com. pers., 2000) que los dos referidos anteriormente, con abundante regeneración de especies leñosas y numerosos géneros típicos de matorrales y bosques secundarios jóvenes. Al comparar los tres fragmentos boscosos, en éste el efecto antropógeno directo ha sido el menor. Aun así, aquí también se observan con relativa frecuencia géneros y especies introducidos, que se han naturalizado y se reproducen exitosamente, vegetativa o sexualmente; entre ellos la herbácea Kalanchoë pinnata (Lam.) Pers. Aquí, con frecuencia los árboles son retorcidos y doblados; en muchos casos es evidente que esto se debe al efecto de grandes bejucos trepadores o a la orientación de la copa creciente hacia un claro (cuando existe fuerte competencia por luz, debido a que la vegetación es densa). No obstante, otros factores podrían tener influencia, tales como un suelo poco profundo o con una capa de arcilla compactada. Este fragmento muestra mayores características de bosque secundario natural que los otros fragmentos comparados: estructura más o menos homogénea, numerosas especies leñosas en regeneración (desde plántulas hasta ca. 4 m de altura), establecimiento de múltiples arbustos y árboles jóvenes, muy pocas epífitas (aunque abundan, desde luego, líquenes y musgos). Algunos bejucos, como Gouania lupuloides (L.) Urb. (Rhamnaceae) y Paullinia sp. (Sapindaceae), son frecuentes e incluso pueden alcanzar gran desarrollo, igual que en los otros bosques.
Especies más comunes en este bosque
Arbóreas y arbustivas: Ardisia sp., Crossopetalum tonduzii, Croton hoffmannii, Croton decalobus Müll. Arg., Eugenia cartagensis, Euphorbia hoffmanniana (Klotzsch & Garke) Boiss., Fraxinus uhdei, Malpighia glabra, Myrcia splendens, Parathesis glabra Donn. Sm., Randia aculeata, Triumfetta cf. semitriloba Jacq., Viburnum stellato-tomentosum (Oerst.) Hemsl., Xylosma velutina, Zanthoxylum limoncello.
Trepadoras: Gouania lupuloides, Malvaviscus sp., Paullinia sp., Philodendron sp.
Herbáceas: Anthurium sp., Anthurium salvinii, Kalanchoë pinnata, Pavonia rosea Schltdl., Spigelia splendens J.C. Wendl., Tradescantia zebrina.
Epífitas: Como podía predecirse, existen pocas especies en este bosque: Orchidaceae (la mayoría fueron fijadas con cuerdas en arbustos y árboles inmaduros en los últimos años: Gongora, Oncidium, Pleurothallis decipiens Ames & C. Schweinf.), Bromeliaceae (Aechmea, Tillandsia, Vriesea); Piperaceae (Peperomia), algunos helechos pequeños (e.g. Hymenophyllum). En la corteza de la mayoría de plantas inmaduras en regeneración, así como en arbustos y árboles en activo crecimiento en el sotobosque, se hallan solamente líquenes epífitos; en la etapa posterior inmediata se hallan líquenes y briófitos juntos.
Regeneración: Al comparar los tres bosques de este estudio, el Bosque Nuevo muestra mayor número y densidad de especies leñosas en regeneración.
II. Comparación de los tres fragmentos boscosos estudiados
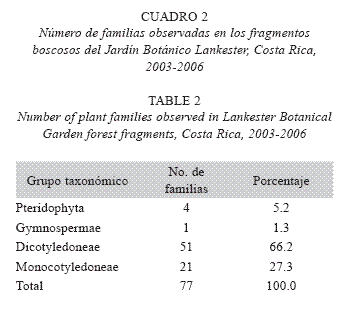
]]> Los parámetros de desarrollo de las plantas considerados en los tres fragmentos boscosos son diámetro y altura (Cuadro 1). El fragmento periférico muestra los datos de diámetro más dispersos y en el fragmento nuevo la altura es considerablemente menor que en los otros dos.El número de familias de los grupos más comunes de espermatófitos es el esperado, con predominio de las dicotiledóneas sobre las monocotiledóneas (Cuadro 2).
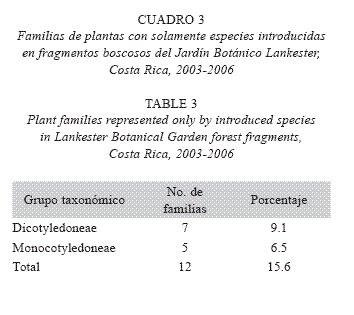
Algunas familias de plantas están representadas en el área de estudio solamente por especies introducidas (Cuadro 3), como se especifica a continuación:
Dicotyledoneae: Anacardiaceae (Mangifera indica L., de Asia oriental), Balsaminaceae (Impatiens walleriana, de zonas tropicales de África), Bignoniaceae (Spathodea campanulata P. Beauv., de África oriental, Tabebuia ipe, de la región del Chaco, Argentina), Crassulaceae (Kalanchoë pinnata, de África y Madagascar), Juglandaceae (Juglans olanchana Standl. & L.O. Williams, del norte de Mesoamérica), Oleaceae (Fraxinus uhdei, de América del Norte) y Proteaceae (Grevillea robusta, de Australia).
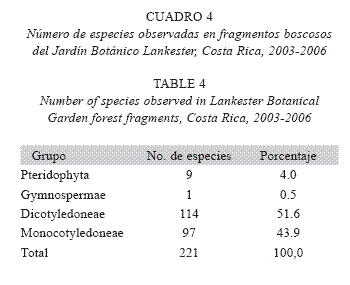
]]> Monocotyledoneae: Amaryllidaceae (Crinum x powellii hort. ex Baker, de África), Asteliaceae [Cordyline fruticosa (L.) A. Chev., Cordyline sp., de Asia oriental], Dracaenaceae (Dracaena deremensis, Dracaena fragrans, Sansevieria trifasciata Prain, de África), Hypoxidaceae (Molineria capitulata, de Asia sudoriental) e iridaceae (Neomarica sp., de Sudamérica).Un tercio de las familias registradas en estos fragmentos (n=26; 33.8%) posee especies introducidas de otras regiones del mundo; son 15 familias (19.5%) en Dicotiledóneas y 11 (14.3 %) en Monocotiledóneas. Además, el número total de especies observadas es relativamente bajo, con una notable abundancia de monocotiledóneas, cercana a la de dicotiledóneas (Cuadro 4). El número de especies herbáceas (n=140; 63.3%) supera ampliamente el de árboles y otros hábitos. Casi 20% de las especies registradas son introducidas (Cuadro 5).
Discusión
Comparación de los tres fragmentos boscosos: Se registró poca diferencia entre los diámetros de los fragmentos intermedio y Periférico, lo mismo que entre las alturas de estos bosques (Cuadro 1). Sin embargo, el diámetro en el Bosque Nuevo es notablemente menor que en los otros dos bosques. Lo observado se refleja en estos datos, porque el Bosque Nuevo es, estructuralmente, mucho menos desarrollado que los otros dos bosques.
El promedio de altura del Bosque Nuevo con la desviación máxima es casi igual al promedio del Bosque Periférico; sin embargo, ambos fragmentos son estructuralmente distintos. Esto puede deberse a la frecuencia de arbustos y árboles pioneros jóvenes de rápido crecimiento en el Bosque Nuevo, que en claros estrechos crecen con el tronco relativamente recto, en lugar de ser retorcido o arqueado (entre ellos Croton, Fraxinus y Myrcia). En las primeras etapas de su ciclo, estos árboles parecen concentrar sus energías en producir un tronco relativamente alto, que les permita alcanzar el dosel o el subdosel rápidamente. El Bosque Nuevo exhibe una regeneración de plantas leñosas mucho más activa que la de los otros fragmentos; esto se refleja en el número de plantas medidas en cinco parcelas (n=105), comparado con el número en diez parcelas del Bosque intermedio (n=73) y del Bosque Periférico (n=118). Sin intervención humana, estas plantas inmaduras estarán sometidas a un fuerte proceso de selección, de modo que ciertas especies presentes en la actualidad, que son exclusivamente pioneras, desaparecerán en próximos años, mientras que otras, inclusive epífitas y trepadoras, podrán establecerse en un bosque más denso, con numerosas especies de árboles maduros.
No se observaron diferencias florísticas notables entre los tres bosques, aunque el Bosque Nuevo es cuantitativamente diferente a los otros dos. Cabe señalar que este es el resultado esperado, porque no existen barreras físicas entre los tres fragmentos de bosque estudiados. El mayor número de familias es de dicotiledóneas, seguidas por las monocotiledóneas (Cuadro 2), puesto que ambos grupos dominan la vegetación tropical del mundo. La diversidad de helechos y afines (Pteridophyta) registrada en este estudio es más baja que la esperada en ambientes naturales equivalentes. Según observaciones personales, razones probables son: 1) Datos climáticos actuales indican que la zona es la más seca del Valle Central entre diciembre y abril (los pteridófitos tropicales son más diversos en zonas muy húmedas todo el año o la mayor parte del año); 2) Los helechos epífitos son más diversos en bosques primarios con varios estratos verticales y numerosos árboles maduros, y 3) Es difícil observar e identificar todos los helechos que crecen en árboles.
El número de especies introducidas parece relativamente bajo (ca. 20% del total). Sin embargo, las que lograron adaptarse se propagan con mucho éxito y forman parte del paisaje y de la composición florística del bosque.
Especies invasoras: Éstas pueden retardar notablemente la regeneración de las especies nativas y, consecuentemente, la diversificación del bosque (Standish et al. 2001). Por su éxito y su abundante propagación, en los fragmentos boscosos del Jardín Lankester pueden considerarse invasoras las siguientes especies introducidas: Dracaena fragrans (Dracaenaceae), Fraxinus uhdei (Oleaceae), Impatiens walleriana (Balsaminaceae), Molineria capitulata (Hypoxidaceae), Phyllostachys aurea (Poaceae) y Syzygium jambos (Myrtaceae).
A partir de estos datos podemos argumentar que la introducción de especies ornamentales debe hacerse cuidadosamente, para evitar la invasión de ecosistemas vulnerables. No obstante, este proceso no siempre puede ser controlado, como lo demuestra la invasión reciente (últimos cinco años) de la hierba africana Crassocephalum crepidioides (Benth.) S. Moore (Asteraceae), que se ha propagado muy rápidamente y ahora se halla con frecuencia en muchas áreas abiertas de Costa Rica; en un área abierta y protegida del Valle Central he contado hasta 80 plantas/ m2 (datos inéditos). En este país se ha estudiado muy poco el efecto de las especies introducidas sobre las nativas en el proceso de regeneración; una excepción es Syzygium jambos (Ávalos et al. 2006, Di Stefano et al. 1998). Por tanto, este es un tema de investigación de gran relevancia en el futuro cercano.
Actualmente es casi imposible regenerar bosques exclusivamente con las especies nativas de cada zona, porque las especies de los alrededores, tanto nativas como introducidas, se establecen inevitablemente por la acción de viento, lluvia y agentes bióticos (polinizadores y dispersores). Algunas especies alóctonas han tenido tanto éxito que se hallan hasta en bordes de bosques primarios; v.g., Impatiens walleriana, de África, una hierba terrestre que invade incluso el hábitat de las epífitas neotropicales, y Hedychium coronarium J. König (Zingiberaceae), de Asia, que forma grandes colonias sobre todo a orillas de ríos. En los últimos años he observado que la orquídea terrestre africana Oeceoclades maculata (Lindl.) Lindl. crece hasta en bosques primarios de Costa Rica (datos inéditos).
]]> Desarrollo de los fragmentos boscosos observados: La relación de plantas leñosas a herbáceas es ca. 1 a 3. Pese a la edad de estos fragmentos boscosos (ca. 35 años en Bosque intermedio y Periférico), las especies leñosas todavía están representadas como en las primeras etapas de la regeneración (primeros 10 a 15 años). Aunque la regeneración de ciertas especies de árboles [por ej., Crossopetalum tonduzii (Celastraceae), Croton decalobus (Euphorbiaceae), Eugenia cartagensis (Myrtaceae), Fraxinus uhdei (Oleaceae), Pachira aquatica (Bombacaceae), Viburnum stellato-tomentosum (Caprifoliaceae) y Xylosma velutina (Flacourtiaceae)] ha sido exitosa a lo largo del tiempo, no sucede lo mismo con la regeneración total del bosque ni con la diversificación de especies arbóreas, en contraste con lo que he observado en áreas regeneradas junto a bosques secundarios o primarios, que aportan abundantes propágulos para restaurar la vegetación natural. Es importante recordar que, con el paso del tiempo, el Jardín Lankester dejó de ser una finca rural, con bosques y matorrales cercanos, y se convirtió en un área verde en medio de urbanizaciones, pueblos y terrenos de cultivo abandonados. Por eso, aquí las fuentes de propágulos de especies de plantas de los antiguos bosques son escasas o nulas. Este aislamiento relativo explica, en gran medida, que los bosques del jardín no hayan alcanzado el desarrollo y la diversidad de otros bosques de edad similar. Varios investigadores (v.g. Barik et al. 1996, Twedt 2006) han observado que cuando las fuentes naturales de semillas son escasas o están relativamente lejos, la vegetación regenerada naturalmente puede ser pobre en especies leñosas.En un fragmento boscoso de nueve años en Tabarcia de Mora, Costa Rica (700-800 m), Fournier & Herrera (1985) registraron 26 familias y 52 especies de árboles. En un fragmento de premontano regenerado durante 20 años en Ciudad Colón (800 m), los mismos investigadores observaron 34 familias y 84 especies de árboles. También datos inéditos de Fournier ilustran el gran desarrollo alcanzado por este último fragmento en 20 años, en claro contraste con los fragmentos de 35 años estudiados en el Jardín Lankester. En este último caso la diversificación del bosque fue mucho más lenta que en los fragmentos estudiados por Fournier. Así se deduce que el aislamiento progresivo de un fragmento boscoso en recuperación puede retardar significativamente y hasta anular el proceso de diversificación vegetal.
La recuperación de un fragmento boscoso es un fenómeno fuertemente influido por la transformación del entorno del área restaurada. Esta transformación puede deberse a factores antropógenos o naturales, o a una combinación de ambos. En el caso del Jardín Lankester, la pérdida progresiva de cobertura vegetal en los alrededores convirtió a los fragmentos boscosos prácticamente en islas, en las que la diversificación se volvió cada vez más lenta y podría estancarse si no intervienen factores bióticos.
En el marco de este estudio se valora positivamente la intervención humana en la regeneración de bosques, ante todo por cultivo de especies que se establezcan exitosamente e impidan la dominancia indefinida de pioneras agresivas, tanto herbáceas como arbustivas. La diversidad de especies puede aumentar más rápidamente si la intervención humana realmente favorece el desarrollo del bosque.
Cuando se plantan especies para recuperar ambientes degradados, en algunos casos el estatus funcional del ecosistema no limita las primeras etapas de establecimiento y las condiciones abióticas juegan un papel crucial en este proceso (Maestre et al. 2006). Sin embargo, en pastizales fuertemente degradados de Colombia la dispersión de semillas está limitada a 10 m de distancia de un fragmento de bosque, pero esta barrera puede superarse por cultivo de semillas y de plántulas y por establecimiento de perchas para atraer dispersores (Aide & Cavelier 1994). La importancia del suelo se refleja en las observaciones de Guariguata (1990) en deslizamientos de tierra de las montañas de Puerto Rico; la parte inferior del deslizamiento tiene un suelo orgánico con abundantes restos vegetales, que generalmente posee mayor concentración de carbono y nutrimentos que la parte superior (con rocas meteorizadas expuestas y suelo mineral). A lo largo de cinco décadas, la regeneración fue siempre mayor en la parte inferior.
Especies epífitas y trepadoras: A menudo es difícil observar, recolectar y determinar estas plantas; además, la mayoría de investigadores carecen de equipo apropiado para subir a árboles. Sin embargo, las observaciones preliminares de este estudio confirman que, a lo largo del tiempo, también en las cortezas de árboles de bosques en regeneración ocurre un proceso de colonización, competencia y sustitución de diferentes tipos de plantas, primero líquenes y briófitos, después vasculares (Benzing 1990), y que este proceso está relacionado con la identidad taxonómica y la edad de las plantas hospederas. Por ejemplo, se observó que los árboles jóvenes de Sapium glandulosum (Euphorbiaceae) no portan ninguna epífita, mientras que algunos árboles viejos (30 o más años) de esta especie albergan líquenes, musgos, aráceas trepadoras, bromeliáceas e incluso algunas orquídeas. Sabemos muy poco sobre la sucesión ecológica de las epífitas, por lo que es relevante estudiar este tema más detalladamente, sobre todo en relación con factores microclimáticos.
Pautas para la recuperación de bosques en Costa Rica: Con el objetivo de que la intervención humana realmente pueda favorecer la recuperación de bosques, una cuestión clave es cómo manejar diferentes estadios del proceso.
Es posible adoptar una de las siguientes estrategias o una combinación de ellas:
A. Regeneración natural. Aquí la única intervención antropógena en el proceso es la protección física del área de recu peración mediante cercas y rondas contra incendios.
]]> B. Regeneración natural más cultivo. En áreas de crecimiento secundario (i.e., matorrales avanzados o bosques secunda rios incipientes) se cultivan diversas especies de plantas (especialmente árboles) de interés particular. Aquí el cultivo de espe cies puede tener uno o varios objetivos:1.Acelerar el proceso de recuperación del bosque,
2.Aumentar la diversidad del nuevo bosque
3.Reintroducir especies de la misma zona que han desaparecido localmente,
4.Cultivar especies de interés particular; v.g., maderables, ornamentales y medicinales. Sin embargo, en este contexto la explotación futura está excluida; de lo contrario, no es apropiado hablar de regeneración ni de recuperación de ecosistemas.
C. Regeneración por cultivo sucesivo. Este es un proceso de dominio antropógeno, sobre todo en terrenos agropecuarios abandonados, como pastizales o prados abandonados. En estas condiciones el proceso
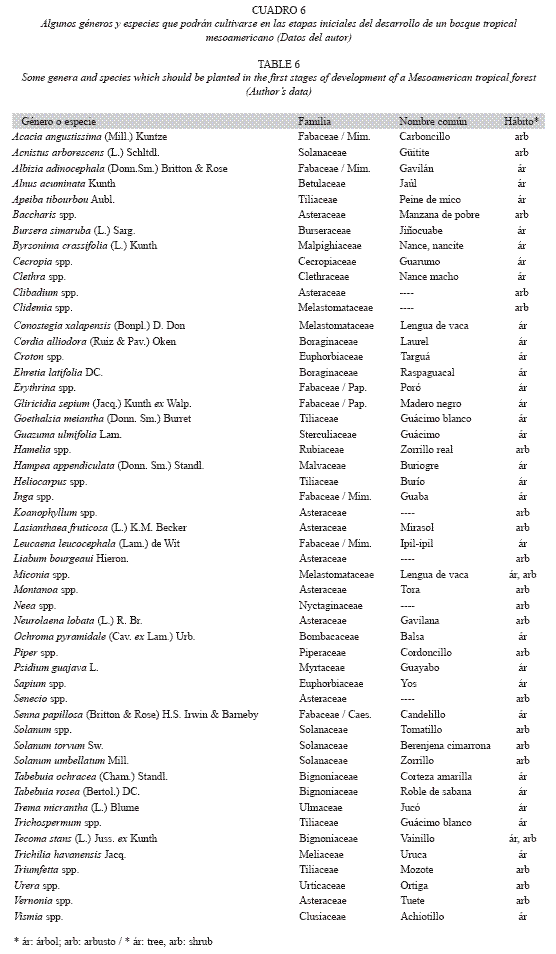
de regeneración natural puede ser muy lento; entonces, se inicia con el cultivo de plantas pioneras de la región (Cuadro 6). En diferentes etapas sucesivas se cultivan: 1) hierbas, arbustos y árboles pioneros, 2) árboles adaptados a bosques secundarios jóvenes, 3) árboles de bosques maduros y 4) trepadoras, epífitas y especies terrestres umbrófilas.
En general, la regeneración natural más cultivo (B) se revela como el proceso más rápido y eficiente (al menos en las condiciones de este estudio, en el Valle Central de Costa Rica). Si ya existe vegetación herbácea y arbustiva y existen parámetros (vigor del crecimiento vegetal) que indiquen que el suelo es fértil, con buena estructura y buen contenido de materia orgánica, puede iniciarse cuanto antes el cultivo de árboles. El proceso (C) parece más lento y difícil, pero existe muy poca información y prácticamente ninguna experiencia sobre regeneración controlada y completa del bosque en Costa Rica. El proceso (A) también es lento; además, diversos factores pueden retrasarlo; por ejemplo, dominancia indefinida de especies herbáceas, arbustivas o trepadoras pioneras. Aún así, el proceso (A) tiene ventajas notables respecto al (C), porque ya existen experiencias prácticas y la inversión de tiempo y dinero puede ser mucho menor.
En diversas latitudes se ha investigado si cercar áreas secundarias que tienen vegetación nativa es una protección suficiente. Se ha demostrado que, a corto plazo, la regeneración de árboles es significativamente mayor y la compactación del suelo es menor en áreas cercadas que en parcelas de control con influencia de ganado o animales silvestres, que incluso devoran los árboles inmaduros. El cercado es un paso inicial importante, pero se requiere de un manejo más activo para lograr la recuperación del bosque (Kuiters & Slim 2002, Spooner et al. 2002, Roovers et al. 2005). Twedt (2006) plantó grupos de árboles de crecimiento rápido junto con árboles de crecimiento lento y observó que esto mejoró la estructura del bosque. Al principio, las plántulas de los árboles de crecimiento rápido tuvieron que ser protegidas de la competencia con hierbas, pero después de cinco años la vegetación alrededor de estos árboles tenía más especies leñosas y de mayor altura que los sitios de control. incluso se ha observado que en los fragmentos boscosos regenerados la tasa anual de descomposición del mantillo del suelo es similar a la de bosques primarios, lo que indica una restauración eficiente de los ciclos de nutrimentos en los fragmentos protegidos (Borders et al. 2006).
En Puerto Rico, Zimmerman et al. (2000) observaron las barreras naturales que dificultan la regeneración de un bosque en un pastizal recientemente abandonado y demostraron que la siembra de semillas de especies pioneras conocidas puede acelerar la tasa de sucesión secundaria, pero algunas especies tendrán que ser plantadas en etapas posteriores de sucesión para poder superar las barreras más fuertes. Al inicio podrán sembrarse (o permitir la regeneración de) arbustos y árboles pioneros y tolerantes de áreas soleadas y suelos pobres (Cuadro 6). La zona de vida y la altitud son determinantes para seleccionar las especies. El ambiente edáfico y la sombra de estos elementos pioneros permitirán el establecimiento (natural o por cultivo) de especies arbóreas propias de bosques, que crecen más lentamente y llegan a ser más longevas.
Existen ejemplos de restauración ecológica exitosa incluso en zonas desérticas extratropicales (v.g. Wang et al. 2006). Por tanto, con condiciones ambientales más favorables se esperarían mejores resultados en la zona neotropical. En vista de que un cambio climático global podría transformar en el futuro las condiciones de los ecosistemas históricos, este hecho debe considerarse a mediano y largo plazo en los planes de restauración ecológica, tal como señalan Harris et al. (2006). Entre otros aspectos, sería difícil o imposible la restauración de ambientes donde ocurre un cambio drástico en la distribución geográfica y altitudinal de numerosas especies animales y vegetales.
Agradecimientos
A la Vicerrectoría de investigación de la Universidad de Costa Rica, por haber apoyado este proyecto (No. 111-A2-158) durante tres años.
]]> Resumen
Se realizaron observaciones sobre sucesión y composición florística en 25 parcelas de 10 m x 2 m en tres fragmentos boscosos secundarios, contiguos, del Jardín Botánico Lankester, Costa Rica. La intervención humana fue positiva en la regeneración de estos fragmentos por 1) protección de una antigua área de cultivo y pastoreo desde 1970, y 2) siembra y propagación de especies vegetales. Se comprobó que el cultivo de plantas nativas e introducidas de crecimiento exitoso puede acelerar el proceso de regeneración, impedir la dominancia de especies pioneras o invasoras y aumentar la diversidad de especies más rápidamente que en procesos comunes de sucesión. La diversificación natural se redujo a medida que la vegetación natural de los alrededores de los fragmentos fue desapareciendo, con una consecuente disminución o erradicación de semillas y otros propágulos, tales como esporas y tallos. Especies introducidas se naturalizaron en estos bosques, se propagan fácilmente y algunas son invasoras. Este estudio propone pautas generales para facilitar la regeneración de bosques neotropicales al combinar regeneración natural y cultivo de especies nativas.
Palabras clave: fragmentos de bosques secundarios, regeneración de bosques, plantas introducidas, sucesión natural, Jardín Botánico Lankester, Costa Rica.
Recibido 03-X-2007. Corregido 09-X-2008. Aceptado 18-XI-2008.
Referencias
Aide, T.M. & J. Cavelier. 1994. Barriers to lowland tropical forest restoration in the Sierra Nevada de Santa Marta, Colombia. Restor. Ecol. 2: 219-229. [ Links ]
Ávalos, G., K. Hoell, J. Gardner, S. Anderson & C. Lee. 2006. impact of the invasive plant Syzigium [sic] jambos (Myrtaceae) on patterns of understory seedling abundance in a Tropical Premontane Forest, Costa Rica. Rev. Biol. Trop. 54: 415-421. [ Links ]
]]>Barik, S.K., R.S. Tripathi, H.N. Pandey & P. Rao. 1996. Tree regeneration in a subtropical humid forest: effect of cultural disturbance on seed production, dispersal and germination. J. Appl. Ecol. 33: 1551-1560. [ Links ]
Benítez-Malvido, J. 1998. impact of forest fragmentation on seedling abundance in a Tropical Rain Forest. Conserv. Biol. 12: 380-389. [ Links ]
Benzing, D.H. 1990. Vascular epiphytes, general biology and related biota. Cambridge Univ., Nueva York, [ Links ]
Borders, B.D., J.C. Pushnik & D.M. Wood. 2006. Comparison of leaf litter decomposition rates in restored and mature riparian forests on the Sacramento River, California. Restor. Ecol. 14: 308-315. [ Links ]
Clarke, F.M., L.V. Rostant & P.A. Racey. 2005. Life after logging: post-logging recovery of a Neotropical bat community. J. Appl. Ecol. 42: 409-420. [ Links ]
]]>Di Stéfano, J.F., L.A. Fournier, J. Carranza, W. Marín & A. Mora. 1998. Potencial invasor de Syzigium [sic] jambos (Myrtaceae) en fragmentos boscosos: El caso de Ciudad Colón, Costa Rica. Rev. Biol. Trop. 46: 567-573. [ Links ]
Dunn, R.R. 2004. Recovery of faunal communities during tropical forest regeneration. Conserv. Biol. 18: 302-309. [ Links ]
Fournier, L.A. & M.E. Herrera. 1979. importancia científica, económica y cultural de un sistema de pequeñas reservas naturales en Costa Rica. Agron. Costar. 3: 53-55. [ Links ]
Fournier, L.A. & M.E. Herrera. 1985. Recuperación del bosque en el Premontano Húmedo y Muy Húmedo del cantón de Mora, Costa Rica. Rev. Biol. Trop. 33: 151-155. [ Links ]
Ganzhorn, J.U., J. Fietz, E. Rakotovao, D. Schwab & D. Zinner. 1999. Lemurs and the regeneration of Dry Deciduous Forest in Madagascar. Conserv. Biol. 13: 794-804. [ Links ]
]]>Gascon, C., G.B. Williamson & G.A.B. da Fonseca. 2000. Receding forest edges and vanishing reserves. Science 288: 1356-1358. [ Links ] [ Links ] </>
Hamann, A. & E. Curio. 1999. interactions among frugivores and fleshy fruit trees in a Philippine Submontane Rainforest. Conserv. Biol. 13: 766-773. [ Links ]
Harris, J.A., R.J. Hobbs, E. Higgs & J. Aronson. 2006. Ecological restoration and global climate change. Restor. Ecol. 14: 170-176. [ Links ]
Helle, P. 1985. Effects of forest regeneration on the structure of bird communities in northern Finland. Holarct. Ecol. 8: 120-132. [ Links ]
Holdridge, L.R. 1987. Ecología basada en zonas de vida. Trad. H. Jiménez Saa. iiCA, San José, Costa Rica. [ Links ] [ Links ] </>
Kuiters, A.T. & P.A. Slim. 2002. Regeneration of mixed deciduous forest in a Dutch forest-heathland, following a reduction of ungulate densities. Biol. Conserv. 105: 65-74. [ Links ]
Maestre, F.T., J. Cortina & R. Vallejo. 2006. Are ecosystem composition, structure, and functional status related to restoration success? A test from semiarid Mediterranean steppes. Restor. Ecol. 14: 258-266. [ Links ]
Mayfield, M.M., D. Ackerly & G.C. Daily. 2006. The diversity and conservation of plant reproductive and dispersal functional traits in human-dominated tropical landscapes. J. Ecol. 94: 522-536. [ Links ]
Nyhagen, D.F., S.D. Turnbull, J.M. Olesen & C.G. Jones. 2005. An investigation into the role of the Mauritian flying fox, Pteropus niger, in forest regeneration. Biol. Conserv. 122: 491-497. [ Links ]
]]>Roovers, P., H. Gulinck & M. Hermy. 2005. Experimental assessment of initial revegetation on abandoned paths in temperate deciduous forest. Appl. Veget. Sci. 8: 139-148. [ Links ]
Spooner, P., i. Lunt & W. Robinson. 2002. is fencing enough? The short-term effects of stock exclusion in remnant grassy woodlands in southern NSW [New South Wales, Australia]. Ecol. Manag. Restor. 3: 117-126. [ Links ]
Standish, R.J., A.W. Robertson & P.A. Williams. 2001. The impact of an invasive weed Tradescantia fluminensis on native forest regeneration. J. Appl. Ecol. 38: 1253-1263. [ Links ]
Tabarelli, M. & C.A. Peres. 2002. Abiotic and vertebrate seed dispersal in the Brazilian Atlantic forest: implications for forest regeneration. Biol. Conserv. 106: 165-176. [ Links ]
Twedt, D.J. 2006. Small clusters of fast-growing trees enhance forest structure on restored bottomland sites. Restor. Ecol. 14: 316-320. [ Links ]
]]>Wang, X.-P., X.-R. Li, H.-L. Xiao & Y.-X. Pan. 2006. Evolutionary characteristics of the artificially revegetated shrub ecosystem in the Tengger Desert, northern China. Ecol. Res. 21: 415-424. [ Links ]
Zimmerman, J.K., J.B. Pascarella & T.M. Aide. 2000. Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restor. Ecol. 8: 350-360. [ Links ]
]]>