Sistemas reproductivos de las plantas en tres hábitats de la planicie costera de Paraguaná, Venezuela
Luis José Lemus-Jiménez1 & Nelson Ramírez2
1 Universidad Nacional Experimental "Francisco de Miranda", Programa de Ingeniería Agronómica, Departamento de Ambiente y Tecnología Agrícola, Coro, Estado Falcón, Venezuela; llemus@unefm.edu.ve.
2 Universidad Central de Venezuela, Facultad de Ciencias, Instituto de Biología Experimental, Centro de Botánica Tropical, Aptdo. 48312, Caracas. 1041-A, Venezuela.Fax. 58-212-753-58-97; nramirez@reacciun.ve.
2 Toda la correspondencia debe ser dirigida a N. Ramírez.
Recibido 21-XI-2002. Corregido 22-XI-2004. Aceptado 01-VIII-2005.
Abstract: Breeding systems of the Paraguaná coastal plain plants, Venezuela. Breeding systems were evaluated for 51 plant species according to life form, pollination system, vegetation type, and phenology, in the coastal plain of Paraguaná Peninsula, Venezuela. Sexual systems were no associated to life form, pollination system, vegetation type, and phenology. The frequency distribution of sexual system was 82.3% hermaphroditism, 15.6% monoecy, and 1.9% dioecy. All sexual systems had a peak during the lowest rainfall. Genetic system distribution was 64.8% self-compatibility (including partially self-compatibility) and 35.2% self-incompatibility. Among self-compatible species, 45.1% were autogamous (19.6% not autogamous). The genetic systems were associated significantly to: (1) plant life form: self-compatible species tend to be herbaceous and self-incompatible plants tend to be woody species; (2) vegetation type: self-compatible species were predominant in the three vegetation types, but in the mangrove the frequency of self-compatible and self-incompatibles was similar; and (3) pollination system: most of the self-compatible species were polyphilous. Mating systems: xenogamous and autogamous species were associated only with plant life forms. Xenogamous plants were mostly woody species and autogamous plants were mostly herbaceous species. The high incidence of hermaphroditism, self-compatibility, and autogamy are related to herbaceous life form, polyphilous pollination system, and climatic conditions, together the insularity of the Paraguaná peninsula. Rev. Biol. Trop. 53(3-4): 415-430. Epub 2005 Oct 3.
Key words: Breeding system, coastal plain, life form, arid zone, Venezuela.
Los sistemas reproductivos resultan de la combinación específica de varias características como compatibilidad genética, sexualidad, variación temporal en la maduración de los sexos o barreras morfológicas florales (Bawa y Beach 1981). De estos atributos, la unisexualidad y la autoincompatibilidad genética representan los caracteres más eficientes en promover la polinización cruzada. En este sentido, la información de los sistemas reproductivos de especies de plantas es importante ya que determinan el flujo de genes y por consiguiente la diferenciación genética entre poblaciones (Ashton 1969, Scariot et al. 1991). Janzen (1970) y Lloyd (1980) postularon que en ambientes donde existen intensas interacciones bióticas y abióticas, aparentemente, se favorece la selección hacia aquellos elementos donde exista un incremento de la recombinación genética mediante el entrecruzamiento. El entrecruzamiento es alcanzado principalmente por la autoincompatibilidad en especies hermafroditas (Bawa 1974, Ruiz-Zapata y Arroyo 1978, Bullock 1985, Arroyo et al. 1993) y por la dioecia (Bawa 1974, Sobrevila y Arroyo 1982, Tanner 1982, Bawa et al. 1985, Bullock 1985). Ambas estrategias están íntimamente asociadas al habito perenne de las plantas (Baker 1955, Ashton 1969, Bawa 1974, Freeman et al. 1980, Ruiz-Zapata y Arroyo 1978, Fox 1985, Bullock 1985, Bawa et al. 1985, Arroyo et al. 1993, Thomas y La Frankie 1993, Ramírez 1993, Oliveira y Gibbs 2000, van Dulmen 2001), polinización generalista en especies dioicas (Bawa y Opler 1975, Bawa 1980, Givnish 1982, Bawa et al. 1985, Fox 1985, Muenchow 1987, Bawa y Beach 1981, Sobrevila y Arroyo, 1982, Ibarra-Manriquez y Oyama 1992, Ramírez 1993), dispersión por frugívoros especializados en especies dioicas (Bawa y Opler 1975, Bawa 1980, Givnish 1982, Muenchow 1987, Steiner 1988, Ibarra-Manriquez y Oyama 1992, Ramírez 1993), y ubicación geográfica de las comunidades: subtropicales (Ej. Tomlinson 1974) y tropicales (Bullock 1985).
]]> En contraste, una alta proporción de especies autocompatibles está asociada al hábito herbáceo, impredictibilidad de polinización, grado de aislamiento geográfico y a la condición colonizadora de las especies. Por ejemplo, altos porcentajes de especies autocompatibles han sido reportadas en un bosque tropical, asociado a la impredictibilidad de los polinizadores (Arroyo y Cabrera 1977, Sobrevila y Arroyo 1982); en la vegetación presente en las islas, asociado al aislamiento geográfico (McMullen 1987, Percival 1974); en la vegetación de morichales, asociado al aislamiento biológico (Ramírez y Brito 1990); y en especies herbáceas (Ramírez y Seres 1994, Jaimes y Ramírez 1999). De acuerdo a la estructura de la vegetación y la frecuencia de formas de vida la incidencia de estrategias que promueven el entrecruzamiento incrementa desde comunidades herbáceas a comunidades boscosas (Bawa 1974, Ruiz-Zapata y Arroyo 1978, Chan 1981, Sobrevila y Arroyo 1982, Bawa et al. 1985, Bullock 1985, MacMullen 1987, Xena et al. 1988, Ramírez y Brito 1990, Arroyo et al. 1993, Ramírez y Seres 1994, Jaimes y Ramírez 1999). En este sentido, las zonas áridas costeras de la Península de Paraguaná, muestran diferentes tipos de vegetación los cuales varían de acuerdo a la frecuencia de las formas de vida (Lemus-Jiménez y Ramírez 2002). Esta característica permite evaluar la importancia de las formas de vida en diferentes habitas bajo condiciones climáticas similares.Por otra parte, la historia geológica de la Península de Paraguaná muestra que a comienzos del cuaternario (pleistoceno), la península constituía una isla, incomunicada de tierra firme y durante el holoceno (época actual) perdió su insularidad con la formación del Istmo de los médanos, hace aproximadamente 4000 años (Audemard 1996). Actualmente en la Península las condiciones climáticas y edáficas están caracterizadas por altas temperaturas, fuerte velocidad del viento, constante arrastre de finas partículas de arena, fuerte radiación y suelos mayormente arenosos calcáreos. Aunado a esto, la fenología de floración está ampliamente superpuesta (Lemus-Jiménez y Ramírez 2002), la fauna de animales antófilos es baja y el grado de especialización de los sistemas de polinización es principalmente generalista (Lemus-Jiménez y Ramírez 2003). De acuerdo a la historia geológica de la Península, las características climáticas predominantemente adversas, la alta incidencia de especies herbáceas, fenología de floración superpuesta y polinización poco especializada, deberían encontrarse una alta incidencia de especies autocompatibles, principalmente autogamas, así como una baja frecuencia de especies dioecia. La autocompatibilidad y la autogamia han sido encontrados como los sistemas reproductivos mas frecuentes en las especies de plantas de las islas oceánicas de los Galápagos (McMullen 1987), asociados a baja diversidad de visitantes florales (McMullen 1993). El presente estudio evalúa los sistemas reproductivos de las especies de plantas presentes en la zona costera de la Península de Paraguaná y la relación con la forma de vida, tipo de hábitat, sistema de polinización y fenología de floración de las especies.
Métodos
Áreas de estudio: El estudio fue realizado en la Península de Paraguaná, ubicada en la región noroccidental de Venezuela. Se seleccionaron tres tipos de vegetación según Huber y Alarcon (1988): Manglar costero (11º40 N, 69º49 W); Herbazal psamófilo (11º50 N, 69º48 W) y Herbazal litoral (11º38 N, 69º44 W). Las tres áreas están a una elevación menor de 10 m de condiciones semiáridas, con vegetación de porte bajo, muchas de ellas rastreras, fijadoras de dunas. La precipitación media anual es de 330.5 mm y la velocidad media anual del viento es de 6.4 km/h (Ministerio del Ambiente y de los Recursos Naturales Renovables).
Selección de las especies de plantas: formas de vida y hábitat: En cada tipo de hábitat fueron establecidas al azar tres áreas de 200 m2, las cuales fueron divididas en parcelas de acuerdo al método descrito por Lemus-Jiménez y Ramírez (2002). La forma de vida y la abundancia de las especies fue determinado, y de acuerdo a la abundancia se estableció estadísticamente la preferencia de las especies de plantas a cada hábitat (ver Lemus-Jiménez y Ramírez 2002).
Sistema de reproducción: Las plantas fueron clasificadas como hermafroditas, monoicas y dioicas de acuerdo a la sexualidad floral y de los individuos de las poblaciones de cada especie. Además se determinó la presencia de dimorfismo floral (heterostília).
Los niveles de incompatibilidad y autogamia al igual que la presencia de agamospermia fueron estudiadas para 51 especies de plantas pertenecientes a los tres tipos de habitas de la Península de Paraguaná. El sistema de reproducción fue establecido de acuerdo a cuatro pruebas controladas en plantas hermafroditas y monoicas, siguiendo la metodología descrita por Ruiz y Arroyo (1978) y Sobrevila y Arroyo (1982). Para un total de 20 flores por individuo y de 10-20 individuos por especie fueron realizadas las siguientes pruebas: (1) Polinización Automática. Las yemas florales fueron aisladas y posteriormente fue cuantificado el número de frutos y semillas producidas espontáneamente sin ningún tipo de tratamiento. (2) Autopolinización. Las flores abiertas, aisladas previamente a la antesis, fueron autopolinizadas con polen de la misma flor o de otras flores del mismo individuo, y aisladas hasta la formación de frutos y semillas. (3) Polinización cruzada. Yemas florales fueron emasculadas (eliminación de estambres en las yemas) y aisladas, y posteriormente durante la antesis fueron polinizadas con polen proveniente de diferentes individuos de la misma especie. Las flores fueron aisladas nuevamente para cuantificar la formación de frutos y semillas. (4) Agamospermia. Yemas florales fueron emasculadas y aisladas hasta la formación de frutos y semillas.
La condición de autoincompatibilidad y autogamia fue establecida mediante la prueba estadística de independencia "G" (Sokal y Rohlf 1969), a un nivel de significancia de p<0.01. De acuerdo a la producción de frutos y semillas producidas por tratamiento, las especies de plantas fueron clasificadas como autocompatibles y autoincompatibles. Las plantas autocompatibles fueron clasificadas como autocompatibles autogamas y autocompatibles no autogamas. El sistema de apareamiento fue establecido de acuerdo a la condición sexual y sistema genético de reproducción. Dos categorías fueron establecidas: especies xenogamas, representadas por especies dioicas, autoincompatibles y autocompatibles no autogamas, y especies autogamas representadas por especies autocompatibles autogamas.
Fenología: Los sistemas sexuales, sistemas genéticos de reproducción y sistemas de apareamiento fueron expresados mensualmente de acuerdo a la información fenológica de floración de las especies de plantas previamente publicada (Lemus-Jiménez y Ramírez 2002). Para los análisis estadísticos fueron consideradas dos categorías discretas: sequía y lluvia.
Sistema de polinización: El sistema de polinización fue establecido de acuerdo al estudio previo realizado por Lemus-Jiménez y Ramírez (2003) y la biología floral de las especies fue establecidas de acuerdo a la metodología descrita por Grases y Ramírez (1998). Las especies de plantas fueron clasificadas de acuerdo al carácter de la visita recibida. Las categorías consideradas y ligeramente modificadas de Faegri y van der Pijl (1979) fueron: 1.- polifílico, especies de plantas polinizadas por diferentes órdenes taxonómicos de polini-zadores. 2.- oligofílico, especies polinizadas por especies de insectos pertenecientes a más de una familia de un mismo orden taxonómico. 3.- monofílico, plantas polinizadas por una o más especies diferentes de una misma familia taxonómica. En este estudio, las avispas y hormigas fueron consideradas como grupos diferente porque difieren notablemente en sus hábitos alimenticios y morfología de sus partes bucales con relación a sus parientes las abejas (Faegri y van der Pijl 1979, Proctor et al. 1996).
]]> Las dependencia e interacciones entre el sistema sexual, el sistema genético de reproducción y el sistema de apareamiento de acuerdo a las formas de vida, tipo de hábitat, sistema de polinización y fenología (sequía y lluvia), fueron analizadas por la prueba de log-linear (StatSoft 2001), usando tablas de dos o tres factores. Además se determinó la interacción, la cual aporta resultados análogos al usado en el análisis de varianza.Resultados
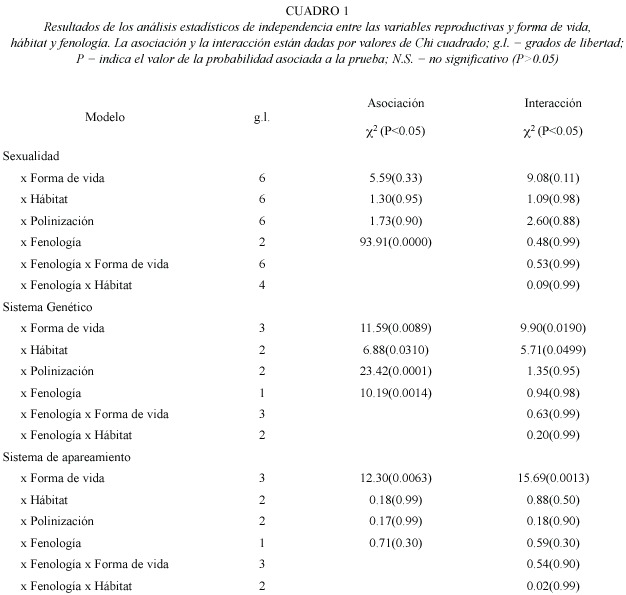
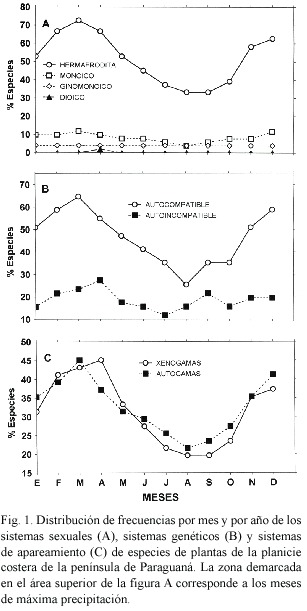
Sistemas sexuales: La distribución de los sistemas sexuales de 51 especies estudiadas fueron: 42 (82.3%) hermafroditas, la cual incluye una especie distílica, 6 (11.7%) monoicas, 2 (3.9%) ginomonoicas y 1 (1.9%) dioica. Los sistemas sexuales no estaban asociados con las diferentes formas de vida (Cuadro 1). El hermafroditismo fue dominante entre todas las formas de vida. La monoecia estaba representada entre las hierbas perennes, hierbas anuales y arbustos (Cuadro 2). Los sistemas sexuales son independientes de tipo de hábitat (Cuadro 1). Las plantas hermafroditas fueron dominantes en los tres tipos de hábitat. Las especies monoicas ocupan el segundo lugar de importancia en el herbazal litoral y herbazal psamófilo (Cuadro 2). Una sola especie dioica fue encontrada en las tres áreas de estudio. Los sistemas sexuales no estaban asociados al sistema de polinización (Cuadro 1). Los sistemas de polinización polifílico, oligofílico y monofílico están principalmente representados entre las especies hermafroditas (Cuadro 2). El sistema de polinización anemofílico fue observado exclusivamente en la especie ginomonoica (Cuadro 2). La distribución de los sistemas sexuales no estaba asociada con la fenología de floración de las especies. Sin embargo, el porcentaje de especies hermafroditas y monoicas decrece desde el inicio a la extrema sequía, y la dioecia ocurrió durante el periodo de mínima precipitación (Fig. 1a). La interacción no significativa indica que la distribución de las formas sexuales no esta afectada significativamente por la forma de vida, hábitat y sistema de polinización (Cuadro 1).
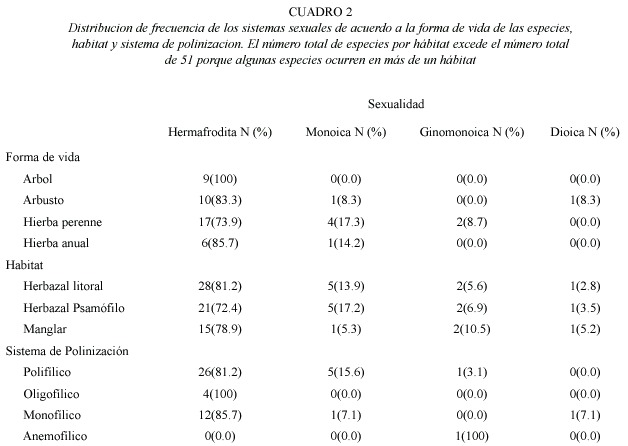
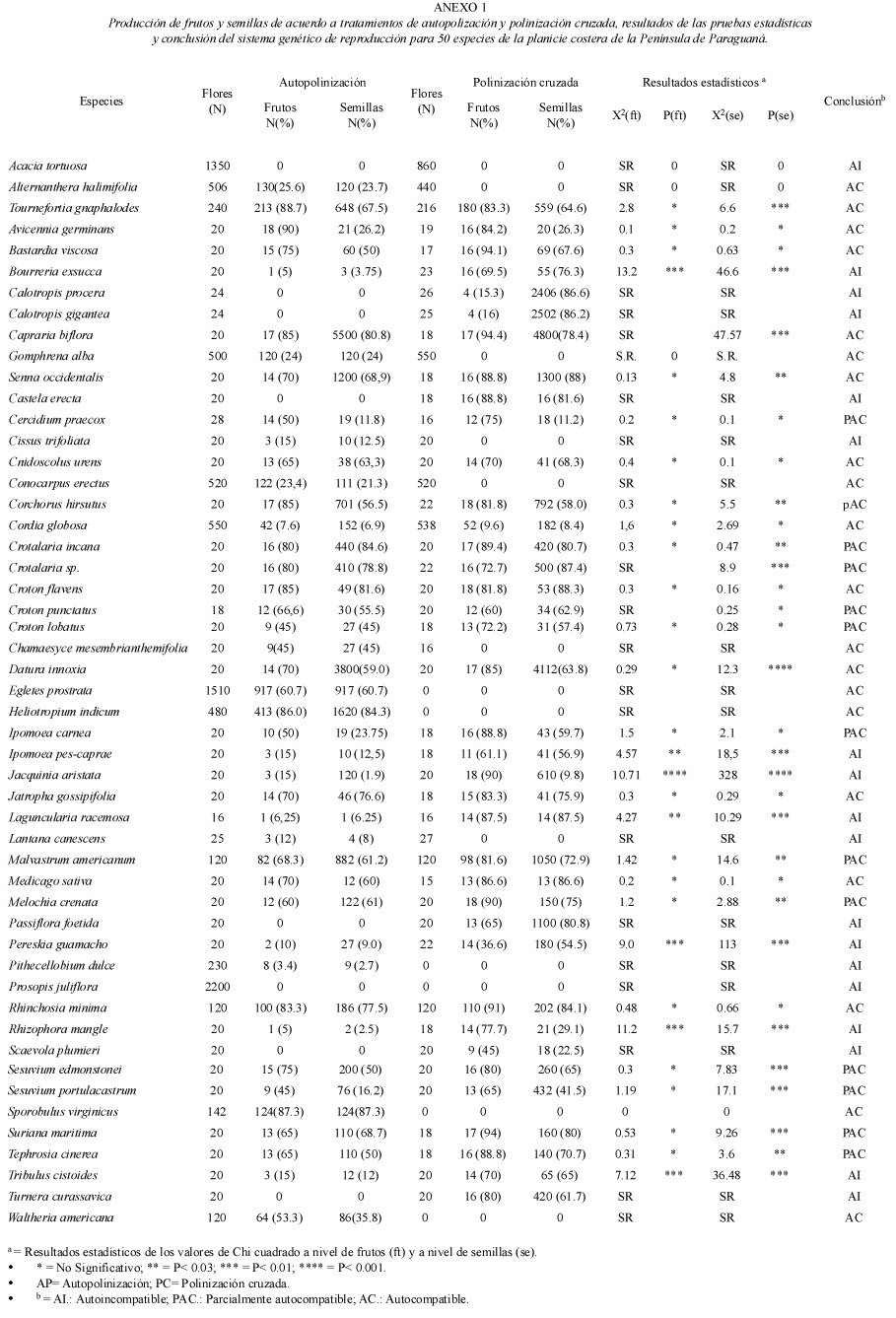
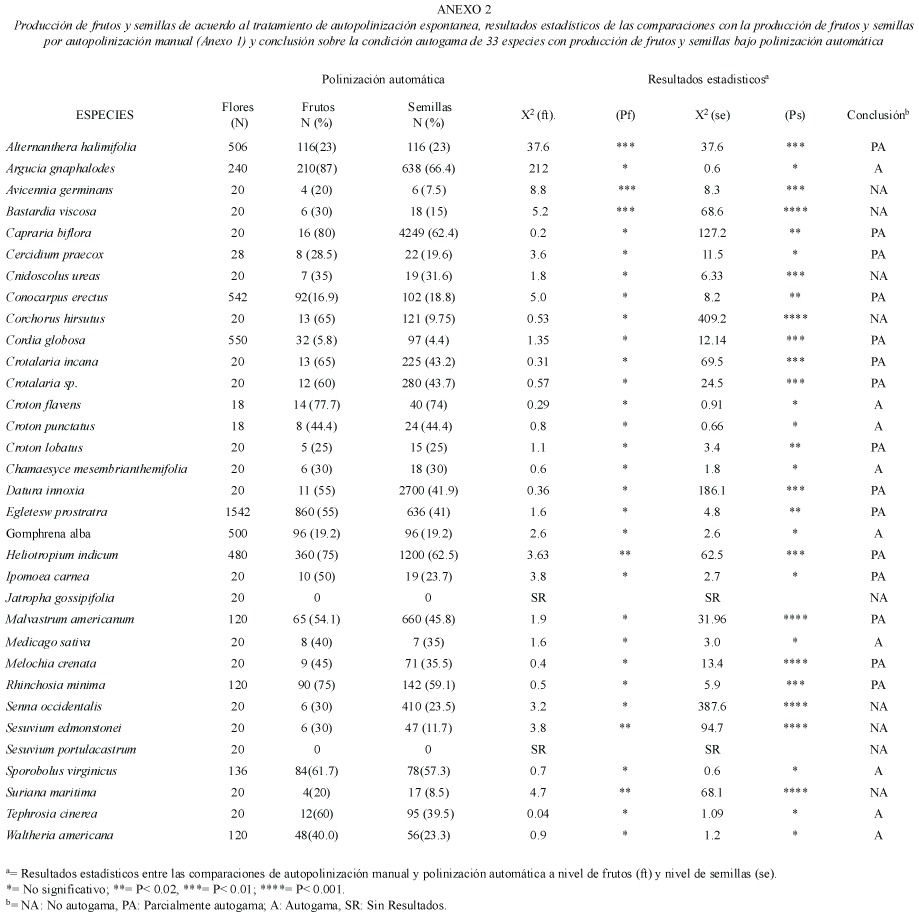
Sistema genético de reproducción: Los resultados de los cruces experimentales de 51 especies de plantas estas detallados en el Anexo 1. Por otra parte, los resultados de las pruebas de autogamia para 33 especies de plantas que presentaron cierta producción de frutos y semillas por polinización espontánea están reseñados en el Anexo 2. La frecuencia de los sistemas genéticos de reproducción fue la siguiente: 18 (35.3%) autoincompatibles y 33 (64.7%) autocompatibles (incluye las parcialmente autocompatibles). Entre las especies autocompatibles, 23(45.1%) resultaron autógamas y 10 (19.6%) no autógamas. No se observó la presencia de agamospermia entre las especies estudiadas. Los sistemas genéticos estaban asociados a las formas de vida (Cuadro 1). El sistema genético mas importante entre los elementos herbáceos fue la autocompatibilidad, mientras que para los elementos arbustivos y arbóreos fue la autoincompatibilidad (Cuadro 3). Los sistemas genéticos están asociados al tipo de hábitat (Cuadro 1), y destaca que la proporción de elementos autocompatibles e autoincompatibles fue muy similar en la vegetación de manglar (Cuadro 3).
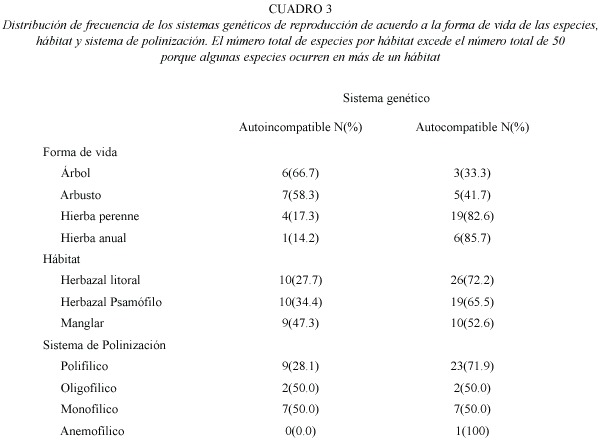
Los sistemas genéticos están asociados al sistema de polinización (Cuadro 1). Las especies autocompatibles mostraron principalmente sistemas de polinización polifílicos (Cuadro 3), mientras los sistemas de polinización monofílicos y oligofílicos fueron observados en iguales proporciones para especies autoincompatibles y autocompatibles (Cuadro 3). Los sistemas genéticos están asociados a la fenología de floración de las especies (Cuadro 1). Las especies autocampatibles son mas abundantes durante el periodo lluvioso. La proporción de especies autocompatibles incrementa desde la extrema sequía hasta el periodo de lluvia, posteriormente decrece (Fig. 1b). En contraste, el porcentaje de especies autoincompatibles incremento al comienzo y mediados de la estación seca (Fig. 1b).
Sistemas de apareamiento: Los sistemas de apareamiento están asociados a las formas de vida de las especies (Cuadro 1). En las especies leñosas, árboles y arbustos, predomina la xenogamia, mientras que para las especies herbáceas fue más abundante la autogamia (Cuadro 4). Los sistemas de apareamiento no están asociado con el tipo de hábitat (Cuadro 1); el porcentaje de especies autógamas y xenógamas fue similar en el herbazal litoral y herbazal psamófilo, pero la proporción de especies xenógamas fue mayor que las especies autógamos en el manglar costero (Cuadro 4). Los sistemas de apareamiento no están asociados al sistema de polinización (Cuadro 1). Los sistemas de polinización oligofílico y monofílico fueron mas abundantes en especies xenógamas, mientras que sistemas de polinización polifílicos ocurrieron en frecuencias similares para especies autogamas y xenogamas. El sistema de polinización anemofílico fue observado exclusivamente en especies autogamas (Cuadro 4). La distribución de los diferentes sistemas de apareamiento no esta significativamente asociado a la fenología de floración de las especies (Cuadro 1). Las especies xenogamas y autogamas incrementaron su frecuencia al comienzo de la estación seca, las cuales decrecen en forma similar hasta la extrema sequía (Fig. 1c).

Discusión
La frecuencia de especies dioicas a nivel comunitario ha sido reportada entre 3.1% y 27% para estudios realizados principalmente en especies leñosos (Ashton 1969, Bawa 1974, Bawa et al. 1985, Fox 1985, Ibarra-Manríquez y Oyama 1992, Thomas y La Frankie 1993). En comunidades donde se han estudiado una alta proporción de especies herbáceas la incidencia de dioecia es relativamente baja como en la flora de Puerto Rico (Flores y Schemske 1984); arbustal mesotérmico (Ramírez 1993) y morichal (Ramírez y Brito 1990). Debido a que la frecuencia de los sistemas sexuales no esta asociado a la forma de vida, tipo de vegetación, sistema de polinización ni fenología de floración en la península de Paraguaná, la baja proporción de especies dioicas podría estar asociada a las limitaciones edáficas y climáticas, así como a la historia geológica de la península de Paraguaná.
]]> La frecuencia de plantas dioicas es baja en zonas insulares (Croat 1979, McMullen 1987, Anderson et al. 2001) e islas biológicas (Ramírez y Brito 1990). La relativa insularidad de la Península de Paraguaná y la abundancia de especies herbáceas podrían también estar limitando la incidencia de dioecia De manera similar al dioicismo, la autoincompatibilidad ha sido asociada frecuentemente con el habito arbóreo (Bawa 1974, Opler y Bawa 1978, Ruiz-Zapata y Arroyo 1978, Bawa et al. 1985, Bullock 1985, Xena et al. 1988, Arroyo et al. 1993). El porcentaje de especies autoincompatibles encontrados para árboles en la Península de Paraguaná (35.3%) es diferente a los obtenidos para otras comunidades vegetales leñosas (Bawa 1974, Chan 1981, Bawa et al. 1985, Bullock 1985, Xena et al. 1988). La asociación entre forma de vida y sistema genético de reproducción evidencia la tendencia a incrementar la proporción de especies autoincompatibles desde hierbas anuales a árboles, y la situación inversa para la proporción de especies autocompatibles. Esta tendencia afecta la proporción de especies autoincompatibles de acuerdo al tipo de vegetación, la cual incrementa desde el herbazal litoral al manglar. En este sentido, la relación entre plantas autoincompatibles y el hábito perenne puede estar asociado con el hecho de que un ciclo de vida largo promueve la oportunidad de varios eventos reproductivos a lo largo de su ciclo de vida, y garantiza la producción de semillas con cierto grado de variabilidad genética (Levin 1975, Grant y Grant 1981).La proporción de especies monoicas (15.6%) en la planicie costera de Paraguaná es similar a los valores reportados para otras zonas tropicales como en bosques decíduos (Jaimes y Ramírez 1999); bosques semidecíduos (Bullock 1985), Bosques lluvioso (Tanner 1982), islas biológicas (Ramírez y Brito 1990), bosque seco ribereño (Xena et al. 1988) y para la flora de Puerto Rico (Flores y Schemske 1984). En este estudio se destaca la importancia del sistema sexual monoico entre las especies herbáceas. La monoecia ha sido considerada como un sistema sexual que promueve la fertilización cruzada por la separación de las funciones sexuales (Schemske et al. 1978, Maynard-Smith 1979, Bawa y Beach 1981, Willson 1983), frecuentemente asociada a la dicogamia (Lloyd y Webb 1986, Kress y Beach 1994). Sin embargo, la mayoría de las especies monoicas en la planicie costera de Paraguaná son autocompatibles y además, muchas presentan altos niveles de autogamia. Probablemente, las condiciones medioambientales extremas pueden favorecer la autogamia en especies monoicas por medio de adaptaciones como la adicogamia y la cercanía entre flores masculinas y femeninas.
La autocompatibilidad es el sistema reproductivo favorecido en plantas pioneras y en especies colonizadoras de las islas continentales (Baker 1955, Baker y Cox 1984, Bawa 1982, Grant y Grant 1981, Lloyd 1980, 1982, McMullen 1987, Thompson y Barrett 1981) o en islas biológicas (Ramírez y Brito 1990). En las comunidades insulares generalmente se observa una alta proporción de especies arbustivas y herbáceas autocompatibles (McMullen 1987, Takebayashi y Morrell 2001). De estos estudios se puede sugerir que la autocompatibilidad en las especies de la planicie podría estar asociada a las características ecológicas y ambientales del área evaluada como son altas temperaturas anuales, escasas precipitaciones, fuerte viento unido a la condición de insularidad mantenida hasta época reciente. En este sentido, se ha reportado que bajo condiciones ambientales muy severas predominan especies de plantas anuales y autocompatibles (Moldenke 1975). Por otra parte, la forma de vida de la plantas, predominantemente herbácea, afecta positivamente la incidencia de especies autocompatibles. Es importante agregar que una alta proporción de especies autocompatibles se reproducen durante el periodo lluvioso, cuando las condiciones favorecen el desarrollo de plantas herbáceas en la península de Paraguaná. Este patrón fenológico puede ser considerado como un parámetro adicional que promueve la alta incidencia de autocompatibilidad en la Península de Paraguaná. Por otra parte, la alta incidencia de autogamia en las especies de la Península de Paraguaná podría estar asociado a la baja diversidad de polinizadores, la abundancia de sistemas de polinización polifílicos y el carácter poliléctico o polifílico de los agentes polinizadores (Lemus-Jiménez y Ramírez, 2003). En este sentido, Sobrevila y Arroyo (1982) han señalado que la alta proporción de especies autocompatibles en un bosque nublado esta asociado a la acción impredecible e insuficiente de los agentes polinizadores. La autogamia asegura el éxito reproductivo en ambientes con un limitado número y diversidad de polinizadores (Lloyd 1980, 1982, Grant y Grant 1981, Thompson y Barrett 1981, Bawa 1982, Baker y Cox 1984).
Finalmente, la alta frecuencia de especies autocompatibles, muchas de las cuales autogamas, está principalmente asociado a la condición herbácea de muchas de las especies presentes en el área de estudio, aunado a la insularidad, condiciones climáticas extremas y la condición polifílica del sistema de polinizadores.
Agradecimiento
Este trabajo forma parte de la tesis doctoral del primer autor, realizada en la Universidad Central de Venezuela y financiada por la Universidad Experimental Francisco de Miranda. Los autores agradecen las sugerencias y recomendaciones de dos árbitros anónimos. Agradecemos a R. Wingfield por la identificación de las especies de plantas. Agradecemos a S. Fernández, S. Musset, N. Pettit y M. Mustiola su colaboración en el trabajo de campo.
Resumen
Los sistemas reproductivos fueron evaluados para 51 especies de plantas en relación a la forma de vida, sistema de polinización, tipo de vegetación y fenología de floración en la planicie costera de la península de Paraguaná, Venezuela. Los sistemas sexuales resultaron independientes de la forma de vida, sistema de polinización, tipo de vegetación y fenología. La distribución de frecuencia de los sistemas sexuales fue 82.3% hermafroditas, 15.6% monoicas y 1.9% dioicas. Todos los sistemas sexuales mostraron un pico de floración durante el mínimo de precipitación. La frecuencia de los sistemas genéticos de reproducción fue 35.3% de especies autoincompatibles y 64.7% de especies autocompatibles, incluyendo especies parcialmente autocompatibles. Entre las especies autocompatibles, 45.1% resultaron autogamas y 19.6% no autogamas. Los sistemas genéticos de reproducción estaban asociados significativamente a: (1) forma de vida de las plantas, especies autocompatibles tienden a ser especies herbáceas y especies autoincompatibles tienden a ser árboles y arbustos; (2) tipo de vegetación, en los tres tipos de vegetación predominan las especies autocompatibles, pero en la vegetación de manglar la proporción de especies autocompatibles y autoincompatibles fue similar; y (3) sistema de polinización, muchas de las especies autocompatibles tenían sistemas de polinización polifílicos. Los sistemas de apareamiento, xenogamia y autogamia, solo estaban asociados significativamente con la forma de vida de las plantas. La xenogamia predominó para especies leñosas, mientras que la autogamia predominó en especies herbáceas. La alta frecuencia de hermafroditismo, autocompatibilidad genética y autogamia son relacionados con la abundancia de especies herbáceas en las áreas de estudio, la polinización polifílica y a las condiciones climáticas extremas unida a la insularidad de la península de Paraguaná.
Palabras clave: Sistema reproductivo, planicie costera, forma de vida, zona árida, Venezuela.
Referencias
]]>Anderson, G.J., G. Bernardello, T.F. Stuessy & D.J. Crawford. 2001. Breeding system and pollination of selected plants endemic to Juan Fernández Islands. Amer. J. Bot. 88: 220-233. [ Links ]
Arroyo, M.T.K. & E. Cabrera. 1977. Preliminary self-incompatibility test for some tropical cloud forest species in venezuela. Incomp. Newsletter 8: 72-77. [ Links ]
Arroyo, M.T.K., F.L.S. & P. Uslar. 1993. Breeding systems in a temperate mediterranean-type climate montane sclerophylluous forest in central Chile. Bot. J. Linn. Soc. 111: 83-102. [ Links ]
Ashton, P.S. 1969. Speciation among tropical forest trees: some deductions in light of recent evidence. Biol. J. Linn. Soc. 1: 156-196. [ Links ]
Audemard, F. 1996. Late quaternary marine deposits of the Venezuelan basin, State Falcón, Northwestern Venezuela. Climates of the past 1: 14-15. [ Links ]
Baker, H.G. 1955. Self-compatibility and establishment after "long-distance" dispersal. Evolution. 9: 347-349. [ Links ]
Baker, H.H. & P.A. Cox. 1984. Further thoughts on dioecism and island. Ann. Miss. Bot. Gard. 71: 244-253. [ Links ]
Bawa, K.S. 1974. Breeding systems of tree species of a lowland tropical community. Evolution 28: 85-92. [ Links ]
Bawa, K.S. & P.A. Opler, 1975. Dioecism in tropical forest trees. Evolution 29: 167-179. [ Links ]
Bawa, K.S. 1980. Evolution of dioecy in flowering plants. Ann. Rev. Ecol. Syst. 11: 15-39. [ Links ]
Bawa, K.S. & J.H. Beach. 1981. Evolution of sexual systems in flowering plants. Ann. Missouri Bot. Gard. 68: 254-274. [ Links ]
Bawa, K.S. 1982. Outcrossing and incidence of dioecism in island floras. Amer. Nat. 119: 866-871. [ Links ]
Bawa, K.S., D.R. Perry & J.H. Beach. 1985. Reproductive biology of tropical lowland rain forest trees. I. Sexual systems and incompatibility mechanism. Amer. J. Bot. 72: 331-345. [ Links ]
Bullock, S.H. 1985. Breeding systems in the flora of tropical deciduous forest in México. Biotropica 17: 287-301. [ Links ]
Chan, H.T. 1981. Reproductive biology of some Malaysian dipterocarp. III. Breeding systems. Malaysian For. 44: 28-36. [ Links ]
Croat, T.B. 1979. The sexuality of Barro Colorado Island flora (Panama). Phytologia 42: 319-348. [ Links ]
Faegry, K. & L. van der Pijl. 1979. Principles of Pollination Ecology. Pergamon. New York. 244 p. [ Links ]
Flores, S. & D.W. Schemske. 1984. Doiecy and monoecy in the flora of Puerto. Rico and the Virgin Island: Ecological correlates. Biotropica 16: 132-139. [ Links ]
Fox, J.F. 1985. Incidence of dioecy in relation to life form, pollination and dispersal. Oecologia 67: 244-249. [ Links ]
Freeman, D.C., K.T. Harper & W.K. Ostler. 1980. Ecology of plant dioecy in the intermountain region of western North America. Oecologia 44: 410-417. [ Links ]
Givnish, T.J. 1982 Outcrossing versus ecological constrain in the evolution of dioecy. Amer. Nat. 119: 849-865. [ Links ]
Grant, B.R. & P.R. Grant. 1981. Explotation of Opuntia cactus by birds on the Galapagos. Oecología 49: 179-187. [ Links ]
Grases, C. & N. Ramírez. 1998. Biología reproductiva de cinco especies ornitófilas en un fragmento de bosque caducifolio secundario en Venezuela. Rev. Biol. Trop. 46: 1095-1108. [ Links ]
Huber, O. & C. Alarcon. 1988. Mapa de Vegetación de Venezuela, Ministerio del Ambiente y de los Recursos Naturales y The Nature Conservancy. Oscar Editores, C.A. Caracas. [ Links ]
Ibarra-Manriquez, G. & K. Oyama. 1992. Ecological correlates of reproductive traits of mexican rain forest trees. Amer. J. Bot. 79: 383-394. [ Links ]
Jaimes, I. & N. Ramirez. 1999. Breeding systems in a secundary deciduous forest in Venezuela: The importence of life form, habitat and pollination specificity. Pl. Syst. Evol: 215: 23-36. [ Links ]
Janzen, D.H. 1970. Herbivores and the number of tree species in tropical forest. Amer. Natur. 104: 501-528. [ Links ]
Kress, W.J. & J.H. Beach, 1994. Flowering plant reproductive systems, p. 161-182. In L.A. McDade, K.S. Bawa, H.A. Hespenheide & G.S. Hartshorn (eds.). La Selva: Ecology and natural history of a neotropical rain forest. Univ. Chicago. [ Links ]
Lemus-Jimenez, L. & N. Ramírez. 2002. Fenología reproductiva en tres tipos de vegetación de la planicie costera de Paraguaná, Venezuela. Acta Cient. Venezolana. 53: 266-278. [ Links ]
Lemus-Jimenez, L. & N. Ramírez. 2003. Polinización y polinizadores en la vegetación de la planicie costera de Paraguaná, Estado Falcón, Venezuela. Acta Cient. Venezolana. 54: 97-114. [ Links ]
Levin, D. 1975. Pest pressure and recombination systems in plants. Amer. Nat. 190: 437-451. [ Links ]
Lloyd, D.G. 1980. Demographic factors and mating patterns in angiosperms. p. 67-88. In O.T. Solbrig (ed.). Demography and evolution in plants populations. Blackwell, Oxford. [ Links ]
Lloyd, D.G. 1982. Selection combined versus separate sexes in seed plants. Amer. Nat. 120: 571-585. [ Links ]
Lloyd, D.G. & C.J. Webb. 1986. The avoidance of interference between the presentation of pollen and stigmas in angiosperms. I. Dichogamy. New Zelands J. Bot. 24: 135-162. [ Links ]
Maynard-Smith, J. 1979. The evolution of sex. Cambridge University Press, New York. 222 p. [ Links ]
McMullen, C.K. 1987. Breeding Systems of Selected Galapagos Islands Angiosperms. Amer. J. Bot. 74: 1694-1705. [ Links ]
McMullen, C.K. 1993. Flower-visiting insects of the Galapagos islands. Pan-Pacific Entom. 69: 95-106. [ Links ]
Moldenke, A.R. 1975. Niche specialization and species diversity along California transect. Oecologia. 21: 219-242. [ Links ]
Muenchow, G.E. 1987. Is dioecy associated with fleshy fruit? Amer. J. Bot. 74: 287-293. [ Links ]
Oliveira, P.E. & P. E. Gibbs. 2000. Reproductive biology of woody plants in a cerrado community of Central Brazil. Flora. 195:311-329. [ Links ]
Opler, P.A. & K.S. Bawa. 1978. Sex ratio in some tropical forest trees. Evolution 32: 812-821. [ Links ]
Percival, M. 1974. Floral Ecology of Coastal Scrub in Southeast Jamaica. Biotropica 6: 104-129. [ Links ]
Proctor, M., P. Yeo & Lack. 1996. The natural history of pollination. Harper Collins, London. 479 p. [ Links ]
Ramírez, N. & Y. Brito. 1990. Reproductive biology of a tropical palm swamp community in the Venezuelan llanos. Amer. J. Bot. 77: 1260-1271. [ Links ]
Ramírez, N. 1993. Reproductive biology in a tropical schrubland of Venezuelan Guayana. J. Veg. Sci. 4: 5-12. [ Links ]
Ramírez, N. & A. Seres. 1994. Plant reproductive biology of herbaceous mon ocots in a venezuelan tropical cloud forest. Pl. Syst. Evol. 190: 129-142. [ Links ]
Ruiz Zapata, T. & M.T.K. Arroyo. 1978. Plant reproductive ecology of a secondary deciduous tropical forest in Venezuela. Biotropica 10: 221-230. [ Links ]
Schemske, D.W., M.F. Willson, M.N. Melampy, L.J. Miller, L. Verner, K.M. Schemske & L.B. Best. 1978. Flowering ecology of some spring woodland herbs. Ecology 59: 351-366. [ Links ]
Scariot, A.O., E. Lieras & J.D. Hay. 1991. Reproductive Biology of the palma Acrocomía aculeata in Central Brazil. Biotropica 23: 12-22. [ Links ]
Sobrevila. C. & M.T.K. Arroyo. 1982. Breeding systems in a montane tropical cloud forest in Venezuela. Pl. Syst. Evol. 140: 19-37. [ Links ]
StatSoft, Inc. 2001. STATISTICA for windowds Computer program manual) Tulsa, OK. [ Links ]
Steiner, K. E. 1988. Dioecism and its correlated in the Cape flora of South Africa. Amer. J. Bot. 75: 1742-1754. [ Links ]
Takebayashi, N. & P. L. Morrell. 2001. Is self-fertilization an evolutionary dead end revisiting an old hypothesis with genetic theories and a macroevolutionary approach. Amer. J. Bot. 88:1143-1150. [ Links ]
Tanner, E. V. J. 1982. Species diversity and reproductive mechanism in Jamaican trees. Biol. J. Linn. Soc. 18: 263-278. [ Links ]
Thomas, S.C. & J. V. La Frankie. 1993. Sex, sixe and interyear variation in flowering among dioecious trees of the Malayan rain forest. Ecology 74: 1529-1537. [ Links ]
Tomlinson, P. B. 1974. Breeding mechanisms in trees native to tropical florida - A morphological assessment. J. Arnold Arbor. 55: 269-290. [ Links ]
Van Dulmen, A. 2001. Pollination and phenology of flowers in rthe canopy of two contrasting rain forest types in Amazonia, Colombia. Pl. Ecol. 153: 73-85. [ Links ]
Willson, M. F. 1983. Plant reproductive ecology. Willey, New York. 282 p. [ Links ]
Xena, N., N. Ramírez & C. Sobrevila. 1988. Notas sobre la dinámica reproductiva de una comunidad vegetal de vega da río. Actes del Simposi Internacional de Botánica Pius Font y Quer. 2: 69-82. [ Links ]
]]>