Specific leaf mass, fresh: dry weight ratio, sugar and protein contents in species of Lamiaceae from different light environments
M.Castrillo*, D.Vizcaíno, E.Moreno & Z.Latorraca
Depto. Biología de Organismos. Universidad Simón Bolívar. Apdo. Postal 89000, Caracas 1080. Venezuela. fax: ++58 2 9063064; mcastr@usb.ve
Received 28-VII-2004. Corrected 14-IX-2004. Accepted 17-X-2004.
Abstract
Samples from eleven species of Lamiaceae were collected from different light environments in Venezuela for laboratory analysis.The studied species were: Plectranthus scutellarioides (Ps), Scutellaria purpurascens (Sp), Hyptis pectinata (Hp), H. sinuata (Hs), Leonorus japonicus (Lj), Plecthranthus amboinicus (Pa) Ocimum basilicum (Ocb), O.campechianum (Occ) Origanum majorana (Orm), Rosmarinus officinali ,(Ro) and Salvia officinalis (So). Protein and soluble sugar contents per unit of area were measured, Specific Leaf Mass (SLM)and fresh: dry weight (FW/DW) ratios were calculated. The higher values for soluble sugars contents were present in sun species: Lj, Pa, Ocb, Occ, Or. m, Ro and So; the lower values were obtained in low light species: Ps, Sp, Hp, Hs. The values of protein content do not show any clear trend or difference between sun and shade environments. The lowest values for the fresh weight: dry weight ratio are observed in sun species with the exception of Lj and Pa, while the highest value is observed in Pa, a succulent plant. The higher values of specific leaf mass (SLM)(Kg DMm-2) are observed in sun plants. The two way ANOVA revealed that there were significant differences among species and between sun and low light environments for sugar content and FW: DW ratio, while SLM was significant for environments but no significant for species, and not significant for protein for both species and environments. The soluble sugar content, FW: DW ratio and SLM values obtained in this work, show a clear separation between sun and shade plants. The sugar content and FW:DW ratio are distinctive within the species,and the light environment affected sugar content, FW:DW ratio and SLM. These species may be shade-tolerant and able to survive in sunny environments. Perhaps these species originated in shaded environments and have been adapting to sunny habitats.Rev.Biol.Trop.53(1-2):23-28.Epub 2005 Jun 24.
Key words:Tropical Lamiaceae, light, shadow, adaptation.
The Lamiaceae (Labiatae) family is an relevant taxonomic group because of its economic importance. This family has approximately 4000 species around the world (Hedge 1992); it has been extensively and interdisciplinarily studied (Harley and Reynolds 1992). In Venezuela it is represented by 21 genera and 80 species, distributed around the country between 0 and 3600 m of elevation (Velazquez 1997). These species have been taxonomically and etnobotanically studied in Venezuela (Velazquez 1997, Velazquez et al. 1997, Velazquez and Orsini 1997). They are mainly herbaceous and the majority of the wild species grow in the forest edge and in savannah.
The species used for commercial purposes are cultivated. Some species grow under direct sun and others grow under tree shade (Velazquez 1997, Avalos and Mulkey 2004). Variation in the light environment can lead to plasticity in physiology (Valladares et al. 2000). The sun and shade plants have been extensively studied. It has been reported that the efficiency of photosynthesis during sunflecks also varies between species, being typically higher in shade tolerant species (Ogren and Sundin 1996) and also that photosynthesis in sunflecks is an important component of carbon gain in understory plants (Leakey et al. 2003). Due to the diversity of light environments in which the Lamiaceae species are found, this study aimed to compare the protein and soluble sugar contents, the Specific Leaf Mass (SLW)and the fresh/dry matter ratio among eleven species of Lamiaceae family growing under natural, perturbed and greenhouse environments and receiving different levels of irradiance.
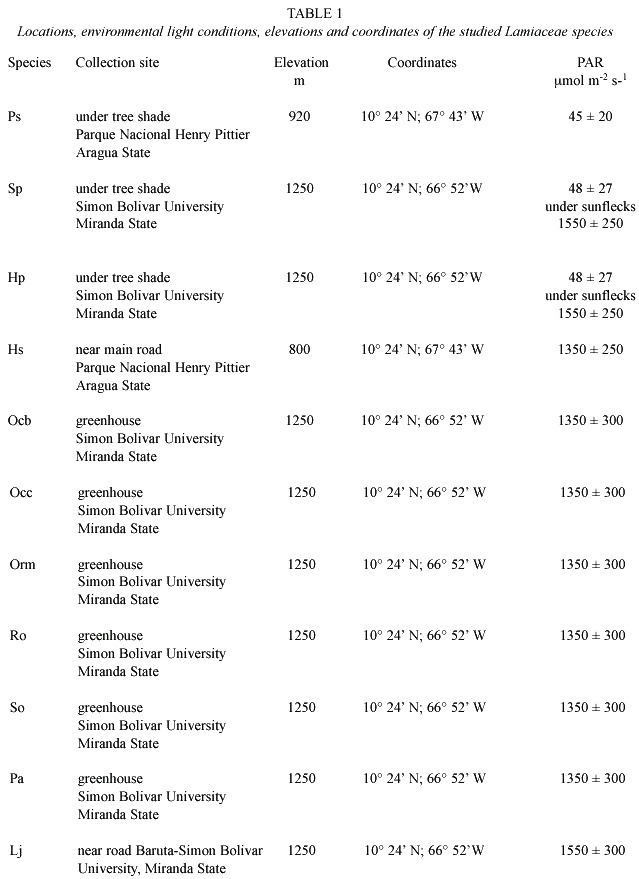
]]> Materials and methodsThe studied species were collected in Venezuela. Table 1 shows specific locations, environmental light conditions,elevations and Young, fully expanded leaves of vegetative plants were collected in plastic bags,kept out of the light and transported immediately to the laboratory inside a cooler at an approximate temperature of 4 °C. Leaves from ten different plants (ten replicates) were used. The leaves were collected between 10:00 and 15:00 hours. The leave collection and PAR measurements were made 5 times for the period of seven months (May to November). The PAR was measured using a LI-189 Quantum Radiometer Photometer (LI-COR, LI-COR, inc. Lincoln, NE, USA).The fresh and dry weight were determined using a Sartorius balance model 2432 and leaf area was measured using a planimeter (Lasiko-Est 1929,Los Angeles Scientific Instrument Co. Inc., Los Angeles, California,USA).
One gram of fresh leaves was grounded with 5 ml of: 0.15 M Tricine-NaOH pH 7.5 (in a ratio of 1:5,w/v) and filtered through four museline layers. From this leaf preparation aliquots were taken for chlorophyll,protein and soluble sugars analyses. The soluble protein content was estimated according to Bradford (1976) using bovine serum albumin for the calibration curve. The soluble sugar content was determined following the anthrone method:l ml of a mixture of 0.2 % anthrone in 100 °/o H2S04 was added to 1 ml of sample and incubated in a 40 ° C water bath for 20 min. After cooling, absorbance was measured at 625 nm in an UV-160A Shimadzu spectrophotometer. Sucrose was used for the calibration curve. Two way analysis of variance was performed among species and between shade and sun species
Results
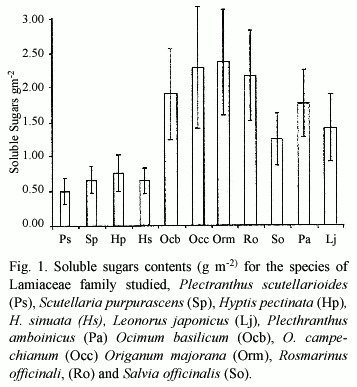
The higher values for soluble sugars contents (g m-2) were present in species grown under full sun (Fig.1): Lj, Pa, Ocb, Occ, Or. m., Ro and So. The lower values were obtained in plants under deep shade conditions, and most frequently shade interrupted by sunflecks: Ps, Sp, Hp and Hs. The two way ANOVA revealed that there were significant difference among species and between light environments (p= 3.85 10-8)
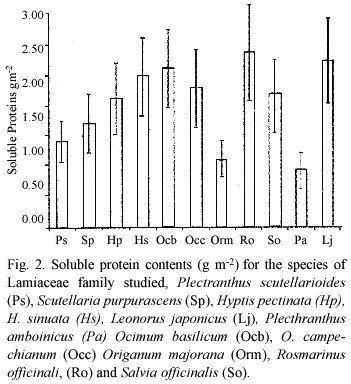
The values of protein content (g m-2 )are shown in figure 2; no definite trend is shown, or differences between sun and shade environments. The lowest values were observed in two sun species: Pa and Orm. The two way ANOVA revealed that there were no significant differences among species and between light environments (p=0.5833)
]]>
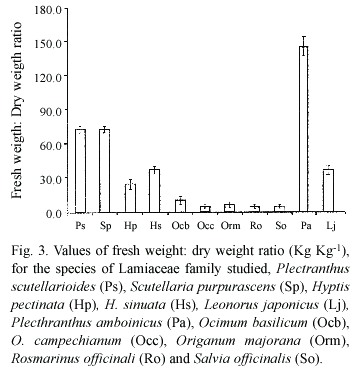
The lowest values for the fresh weight: dry weight ratio (Kg.Kg-1 ) (Fig.3)are observed in species growing under full sun, with exception of Lj and Pa.The highest value is observed in Pa, which is a succulent plant. The two way ANOVA revealed that there were significant difference among species and between light environments (p<2.2 10-16)
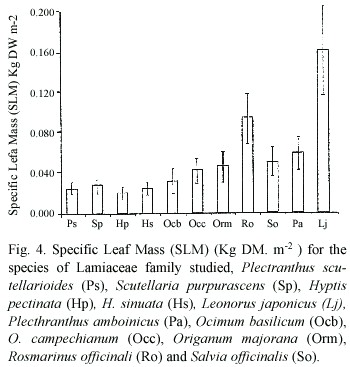
The lower values for Specific Leaf Mass (SLM)(Kg DWm-2 )are observed in shade plants (Fig.4);the higher values are observed in sun plants, Lj being the highest, followed by Ro. Both plant species show a combination of high leaf dry mass and low leaf area. In this case, the two way ANOVA revealed that there were no significant difference among species and significant differences between light environments (p=1.8 10-4 ).
Discussion
The analyzed parameters showed that sugar content and fresh weight: dry weight ratio presented significant differences among species and between light environments; SLM was significant for environment and no significant among species and protein contents were no significant for both species and light environments. High soluble sugar levels are related to high irradiance conditions, and are also correlated to high SLM. We found that both parameters are higher in species growing under full sun. SLM is correlated to light environment; differences in environmental conditions, including PAR, prior to and /or during sampling, have shown to affect SLM (Lugg and Sinclair 1979). The fresh weight: dry weight ratio(FW:DW) is related to both, species and environment.
The two way ANOVA shows that sugar content and FW:DW ratio are significantly different among species and non so SLM and protein content; on the other hand, sugar content, FW:DW ratio and SLM are significantly different between sun and low light environments.
]]> Fjeld (1992) reported an increase in NSC (non-structural carbohydrates) at high values of irradiance. Niinimets (1995) reported that most of the increase in SLW can be attributed to an increase in soluble carbohydrates. Working with canopy leaves of Nothofagus fusca trees, Griffin et al.(2001) found that leaf protein content, non-structural carbohydrates and leaf mass per unit area (LMA), all decreased with depth in the canopy. We did not find any definite difference in protein content between plants either from sun or shade environments.Valladares and Pearcy (1998) working with sun and shade plants of Heteromeles arbutifolia, a Californian chaparral shrub, reported that sun leaves have higher leaf mass per unit area than shade leaves.
In Claytonia virginica, shading reduced SLW and total biomass (Anderson and Eickmeier 1998). Working with coffee, Fahl et (1994)reported that specific leaf weight was 15% higher in plants grown in full sunlight than in shaded areas.
The highest SLM value obtained for L.japonicus could be associated with the direct sun growth environment, as well to the related weed condition of this plant species.
Nagy and Proctor (2000) mentioned that small leaf size and high specific leaf weight are xeromorphic features and that have proposed as adaptive character.
In nine of the eleven species studied here, belonging to the subfamily Nepetoidea, the chlorophyll a: b ratio (Chl a/b)was low, suggesting a prevailing shade aspect regardless of cultivated or wild origin (Castrillo et al.2001).
Morales et al.(1991) mentioned that plants exhibit several kinds of adaptation to the prevailing irradiance, phenotypic adaptation (modulative and modificative)and genotypic adaptation (heliophyte and sciophyte plants) and that these adaptations are not mutually exclusive, bit superimposed so that they permit fine adjustments that guarantee the greatest possible efficiency of radiant energy utilization (Larcher 1983).
The soluble sugar content, FW:DW ratio and SLM in this work, show a clear separation between sun and shade plants.
In the species studied here, the sugar content and FW:DW ratio are distinctive within themselves and the light environment affected the sugar content, FW:DW ratio and SLM. It appears that the Lamiaceae studied species could be shade tolerant and are able to live in sunny environments; moreover, according to these results and those on chlorophyll content reported before (Castrillo et al.2001) it could be interpreted that this species have shade environment origins and have been occupying, and adapting to sunny habitats based on the induced plasticity in physiology caused by variation in the light environment (Valladares et al. 2000).
Acknowledgments
]]> We thank CONICIT (National Council for Research and Technology) for financial support (Project RP-VII-240054).Resumen
En once especies de la familia Lamiaceae: Plecthranthus scutellarioides R. Br. (Ps); S cutellaria purpurascens Sw.(Sp); Hyptis pectinata (L.) Poit.(Hp); Hyptis sinuata Kunth (Hs); Ocimum basilicum L.(Ocb); Ocimum campechianum Miller.(Occ); Origanum majorana L.(Orm); Rosmarinus officinalis L.(Ro); Salvia officinalis L.(So) and Plecthranthus amboinicus (Lour) Spreng.(Pa) and Leonorus japonicus Hout., (Lj), provenientes de diferentes ambientes de exposición a la luz solar, fueron medidos los contenidos de proteínas y azúcares, expresados por unidad de área y fueron calculados la masa específica foliar (MEF)y la relación peso fresco: peso seco (PF:PS). Los mayores valores en el contenido de azúcar fueron observados en plantas que crecieron a exposición directa de luz solar: Lj, Pa, Ocb, Occ, Or. m, Ro and So; mientras que los menores valores se presentaron en aquellas plantas provenientes de ambientes más frecuentemente sombreados e interrumpidos por rayos de sol: Ps, Sp, Hp, Hs. Los valores en el contenido de proteínas no muestran diferencias. Los menores valores de PF:DW fueron observados en las plantas expuestas a radiación solar directa, con la excepción de L. japonicus y P. amboinicus ,esta última es suculenta. Los mayores valores de MEF se presentaron en plantas de sol. El análisis de varianza de dos vías reveló que había diferencias significativas entre especies y ambientes para el contenido de azúcares y la relación PF: PS, mientras que para MEF, la diferencia fue significativa solo entre ambientes; en el caso del contenido de proteínas las diferencias no fueron significativas para especies y ambientes. El contenido de azúcares, la relación PF: PS y la MEF muestran una clara separación entre las especies de sol y las de sombra. Parece que en las especies de Lamiaceae estudiadas, el contenido de azúcares, y la relación PF: PS son distintivos entre las especies mismas y la radiación solar afecta el contenido de azúcares, la relación PF: PS y la MEF. Podría suponerse que estas especies sean tolerantes a la sombra y capaces a su vez, de vivir en ambientes soleados, más aún, de acuerdo a estos resultados y a otros relativos a estas mismas especies, pudiera interpretarse que tienen su origen en ambientes sombreados y que han ido ocupando y adaptándose a ambientes soleados, basándose en la plasticidad fisiológica inducida por las variaciones ambientales de luz.
Palabras clave: Lamiaceae, tropical, luz, sombra, adaptación.
References
Anderson, W. B. & W. G. Eickmeir. 1998. Physiological and morphological responses to shade and nutrient additions of Claytonia virginica (Portulacaceae): implications from the vernal dam, hypothesis. Can: J. Bot. 76:1340-1349. [ Links ]
Avalos, G. & S. S. Mulkey. 2004. Photochemical efficiency of adult and young leaves in the neotropical under-story shrub Psychotria limonensis (Rubiaceae) in response to changes in the light environment. Rev. Biol.Trop. 52(4):839-844. [ Links ]
Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem,71:248-54. [ Links ]
Castrillo, M., D. Vizcaíno, E. Moreno & Z. Latorraca. 2001. Chlorophyll content in some cultivated and wild species of the family Lamiaceae. Biol. Plant. 44: 423-425. [ Links ]
Fahl J.I., M. L. C. Carelli, J. Vega & A. C. Magalhaes. 1994. Nitrogenand irradiance level affecting net photosynthesis and growth of young coffee plants (Coffea arabica L). J. Hort. Sci. 69:161-169. [ Links ]
Fjeld, T. 1992. Effects of temperature and irradiance level on carbohydrate content and keeping quality of Christmas begonia (Begonia x cheimantha Everett). Sci. Hort. 50:219-228. [ Links ]
Griffin, K. L., D. T. Tissue, M. H. Turnbull, W. Schuster & D. Whitehead. 2001. Leaf dark respiration as a function of canopy position in Nothofagus fusca trees grown at ambient and elevated CO2 partial pressures for 5 years. Funct. Ecol. 15:597-605. [ Links ]
Harley R. M. & T. Reynolds (eds.). 1992. Advances in Labiatae Science.The Royal Botany Gardens Kew, Richmond. [ Links ]
Hedge I. C. 1992 . A global survey of the biogeography of the Labiatae .In R.M.Harley &T.Reynolds (eds.). Advances in Labiatae Science.The Royal Botany Gardens Kew, Richmond. pp 7-17. [ Links ]
Larcher,W.1983.Physiological Plant Ecology.Springer, Berlin.pp 303. [ Links ]
Leakey, A. D. B., M. C. Press, J. D. Scholes. 2003. Patterns of dynamic irradiance affect the photosynthetic capacity and growth of dipterocarp tree seedlings. Oecologia 135:184-193. [ Links ]
Lugg, D. G. & T. G. Sinclair. 1979. A survey of soybean cultivars for variability in specific leaf weight. Crop Sci. 19:887-892. [ Links ]
Morales D., M. S. Jimenez & M. Caballero. 1991 Morphological and gas exchange response of Canarina canariensis (L.) Vatke to sun and shade. Photosynthetica 25:481-487. [ Links ]
Nagy, L. & J. Proctor. 2000. Leaf delta 13 C signatures in heath and lowland evergreen rain forest species in Borneo. J. Trop. Ecol. 5:757-761. [ Links ]
Niinimets, U. 1995. Distribution of foliar carbon and nitrogen across the canopy of Fagus sylvatica: adaptation to a vertical light gradient. Acta Oecol. Pl, 16:525-541. [ Links ]
Ogren, E. & U. Sundin. 1996. Photosynthetic responses to variable light:a comparison of species from contrasting habitats. Oecologia 106:18-27. [ Links ]
Valladares, F. & R. W. Pearcy. 1998. The functional ecology of shoot architecture in sun and shade plants of Heteromeles arbustifolia M Roem., a Californian chaparral shrub. Oecologia 114:1-10. [ Links ]
Valladares, F., S. J. Wright, E. Lasso, K. Kitajima & R. W. Pearcy. 2000. Plastic phenotypic response to light of 16 congeneries shrubs from a Panamanian rainforest. Ecology 74:1925-1936. [ Links ]
Velázquez, D. 1997. Clave para los géneros de Lamiaceae en Venezuela. Acta Bot. Venez. 20:1-42 [ Links ]
Velázquez D., E. Arrioja & S. Tillet. 1997. Usos populares de Lamiaceae en Venezuela. Acta Bot. Venez. 18: 5-20. [ Links ]
Velázquez D., & G. Orsini. 1997. Aportes al conocimiento de la familia Lamiaceae en Venezuela I. Ajuga, Teucrium y Scutellaria. Acta Bot. Venez 20:93-115. [ Links ]
]]>