Abstract:Live coral predation by fish in Tayrona Nature National Park,Colombian Caribbean. Live coral predation by fish was evaluated in two bays of the Tayrona National Natural Park (Colombia),as a possible biological agent causing coral mortality.Visual censuses were used to identify the most important predator. Predation incidence was determined by examining all colonies present in permanent belt transects (20 x 2 m)in two reef environments (one dominated by Colpophyllia natans and the other one by Montastraea faveolata ),for two climatic seasons (rainy and dry seasons).The parrotfish Sparisoma viride was the most important predator due to its biting frequency and bite size.S .viride adults of the initial and terminal phases,removed live tissue and part of the calcareous matrix of M.faveolata ,M.annularis ,Porites astreoides and C.natans ,of which,the last one lost a major amount of tissue per area (3.51 cm2 )and volume (3.22 cm3) per bite.A negative exponential tendency (r2 = 0.94), between coral density and volume removed was found,indicating that the coral density determines the bite s damage.There is no clear relationship between predation incidence and climatic seasons at the sites studied.At Chengue and Gayraca bays,live coral predation is one of the factors contributing to coral tissue loss and could have important consequences on the dynamic of these reefs.Rev.Biol.Trop.52(4):883- 895.Epub 2005 Jun 24.
Key words:Predation of corals,reef fishes,Sparisoma viride ,Colombian Caribbean.
Los arrecifes coralinos son sistemas dinámicos sometidos a procesos erosivos o de destrucción, mediados por factores físicos (como tormentas,altas temperaturas,oleaje, corrientes,etc.)y por agentes biológicos y antropogénicos (Hubbard 1997,García et al. 2003,Zubillaga et al.2003).La bioerosión consiste en la remoción y debilitamiento del esqueleto calcáreo de las especies que construyen el arrecife por parte de un organismo a través de sus diversas actividades (Glynn 1997).Entre los principales agentes biológicos de destrucción coralina están los erosionadores epilíticos o raspadores,compuestos por peces,moluscos y erizos (Hutchings 1986,Bruggemann et al.1996,Glynn 1997, Toro-Farmer et al.2004).Los peces pueden ser bioerosionadores activos del coral cuando fragmentan piezas vivas de las colonias mientras se alimentan de ellas,extraen pólipos individuales del esqueleto coralino,extraen otros invertebrados de las colonias de coral o pastorean las algas que crecen sobre el sustrato coralino muerto (Randall 1974,Hixon 1997). Los consumidores directos o peces coralívoros están representados por algunas pocas familias como Chaetodontidae (mariposas),Balistidae (ballestas)y Tetraodontidae (globos)(Glynn et al.1972,Glynn 1990,Guzmán y López 1991,Hixon 1997),y la mayoría consumen sólo partes suaves sin necesidad de alterar el esqueleto coralino.Los peces loro (Scaridae) aunque no son comúnmente mencionados como coralívoros,pueden remover tejido vivo mediante excavación directa del sustrato coralino (Hutchings 1986,Glynn 1990).
El efecto que tienen los peces loro en los arrecifes coralinos ha sido un tema de muchas controversias y fue revisado por Randall (1974).Desde 1831-1836,durante el viaje de C.Darwin en el Beagle a las Islas Cocos-Keeling,se han mencionado las actividades erosivas de los peces loro sobre el coral.Estos peces han sido ampliamente estudiados con el propósito de medir la intensidad y los efectos de su pastoreo en arrecifes coralinos tropicales. Dentro de los estudios realizados en el Caribe se han encontrado diferencias muy marcadas en sus resultados y conclusiones.Algunos sugieren que bajo altas presiones de pastoreo los corales podrían obtener un beneficio porque los peces loro reducen la competencia por espacio entre los corales y las algas (Wanders 1977, Lewis 1986,Birkeland 1997).A diferencia de éstos,otros autores indican que los peces loro frecuentemente arrancan tejido coralino vivo al alimentarse de las algas que crecen adyacentes a estos sustratos (Frydl 1979,Bellwood y Choat 1990).Más aún,aunque los peces loro han sido reconocidos como un componente mayor de las asociaciones de herbívoros en el Caribe (Wanders 1977,Hay 1984,1985),se ha comprobado que ciertas especies de escáridos en ocasiones muerden directamente el coral vivo (Frydl y Stearn 1978,Frydl 1979,Bellwood y Choat 1990,Bruggemann et al.1994a).Otros estudios (Frydl 1979,Bruggemann et al.1994a, Bruckner y Bruckner 1998a)han registrado eventos de depredación de coral más frecuentes, pero restringidos a los adultos de Sparisoma viride (Bonaterre,1788),como principal depredador y Scarus vetula Bloch &Schneider 1801. Entre estas dos especies mencionadas como consumidoras se han observado diferencias en el modo de pastorear sobre el coral vivo.Debido a que S.viride forrajea mediante excavación, logra erosionar mayores proporciones de carbonato de calcio fresco (Bruggemann et al.1996) y por lo tanto deja impreso sobre el coral visibles huellas de sus mordiscos (Frydl y Stearn 1978, Bruckner y Bruckner 1998a).
Con el propósito de ampliar el conocimiento sobre la depredación de coral por peces, este estudio pretendió reconocer las especies de peces depredadoras de coral y cual de ellas tiene un mayor efecto sobre el arrecife. Además,determinar las especies de corales más comúnmente depredadas,la cantidad de coral vivo que puede ser removido por mordisco y la incidencia de la depredación en dos épocas con marcadas diferencias climáticas.
]]>
Materiales y métodos
Las observaciones de campo fueron realizadas durante agosto de 1999 a febrero de 2000 en Chengue (11 °20 N;74 °08 W)y Gayraca (11 °20 N;74 °07 W),dos bahías contiguas localizadas dentro del Parque Nacional Natural Tayrona,al noreste de la ciudad de Santa Marta,en la costa Caribe colombiana.
Reconocimiento del principal depredador de coral vivo:Mediante buceo con equipo SCUBA,se realizaron 33 censos de reconocimiento rápido (30 minutos por censo),durante los cuales el observador se desplazaba lenta y erráticamente a través de la profundidad previamente seleccionada.Cada vez que un pez realizaba una acción de mordisqueo sobre la superficie viva del coral se registró la especie de coral atacada,la especie de pez y la fase sexual para el caso de los escaridos (juvenil, adultos inicial y terminal).Además se midió el tamaño del mordisco (con una reglilla plástica) sobre su parte más larga y se categorizó dentro de la siguiente escala: imperceptible, pequeño ( 5 mm),mediano (6-14 mm)y grande ( 15 mm);esto para estimar de una manera simple y rápida el grado de afección por mordisco.Se evaluaron dos estaciones en Chengue y tres en Gayraca en los niveles de profundidad (somero=2-5 m,medio=9-12 m y profundo=16-19 m)presentes en cada estación y se efectuaron al menos tres censos por nivel de profundidad.
Con base en los datos mencionados anteriormente,se determinó la principal especie de pez depredadora de coral vivo y el nivel de profundidad donde su acción de mordisqueo es mayor,a partir de la frecuencia relativa de mordisqueo y del tamaño de los mordiscos o cicatrices individuales.
Características de los mordiscos individuales hechos por la principal especie depredadora: Con un calibrador plástico se midió el largo (L),ancho (W)y la profundidad (P)del mayor número posible de mordiscos individuales ya presentes en el arrecife sobre diferentes especies de corales.Con estos datos se estimó el área (A=L x W)y volumen (V=A x P)por mordisco.Los datos de áreas y volúmenes de mordiscos fueron caracterizados en máximos, mínimos y promediados por especie de coral para cuantificar la magnitud del daño en términos de mordiscos individuales.De otro lado, mediante análisis de regresión se estableció la relación ente la densidad coralina (g/cm3)y el tamaño promedio de los mordiscos (volumen en cm3).Los valores de las densidades esqueléticas de los corales fueron tomadas de varias fuentes:(Highsmith 1981 fide Hughes 1987, Hughes 1987, R. Dodge, com.pers.).
Estacionalidad en la incidencia de depredación:En el área de Santa Marta las comunidades de algas presentan una fuerte estacionalidad en su abundancia,con valores altos de cobertura de macroalgas durante los meses de aguas frías (enero-marzo)y valores bajos durante los meses de altas temperaturas (septiembre-noviembre)(Díaz-Pulido y Garzón-Ferreira 2002).Debido a que dicha estacionalidad podría incidir en la intensidad de la actividad de depredación de tejido coralino por parte de los peces (ante la escasez o abundancia de las algas para los herbívoros), durante los meses de octubre (época de lluvia) y febrero (época seca)se efectuaron muestreos para determinar si existía estacionalidad en la incidencia de depredación sobre los corales. Para ello se evaluaron dos ambientes arrecifales con signos de depredación y caracterizados por su homogeneidad en la composición estructural coralina,uno dominado por Colpophyllia natans (Houttuyn,1772)(CNAT)y el otro por Montastraea faveolata (Ellis &Solander, 1786)(MFAV).Se muestrearon dos bandas (20 x 2 m) por ambiente en cada bahía,para un total de ocho bandas (320 m2 de arrecife examinado). Las bandas se instalaron permanentemente con varillas de hierro galvanizado a una profundidad media (6-12 m)y se delimitaron con una cinta métrica.En cada banda se inspeccionaron todas las colonias coralinas presentes y se registró visualmente la presencia y magnitud (cobertura relativa a la colonia)de las lesiones de depredación (DE)hechas por peces en el tejido vivo.Partiendo de que los peces loro pastorean preferentemente sobre tapetes de algas filamentosas (Bruggemann et al.1994a, 1994b,1996),la cobertura de dichas algas se midió simultáneamente en las mismas bandas, de forma similar al estudio de Díaz-Pulido y Garzón-Ferreira (2002),mediante estimaciones visuales utilizando cinco cuadrantes de 25x25 cm colocados aleatoriamente sobre áreas del arrecife cubiertas por algas.La incidencia de las lesiones se expresó en términos de frecuencia de ocurrencia de dicha condición en cada época climática y se evaluó mediante estadística descriptiva (promedio y error estándar=EE)por ambiente arrecifal y por especie de coral afectada en cada ambiente.La cobertura promedio de tejido coralino afectado fue calculada para las especies de corales depredadas en cada ambiente arrecifal y en las dos épocas climáticas.Para conocer si había diferencias en las incidencias de depredación entre las épocas para cada ambiente arrecifal y para las especies de corales más comúnmente depredadas,se realizaron pruebas t-Student.Se estimó la cobertura promedio de algas filamentosas (en términos de porcentaje)en cada ambiente arrecifal y época climática y se relacionó con la incidencia por época y ambiente,mediante correlaciones de Spearman.Además se realizaron pruebas t-Student con las coberturas de las filamentosas por ambiente para conocer si existían diferencias entre épocas.
Resultados
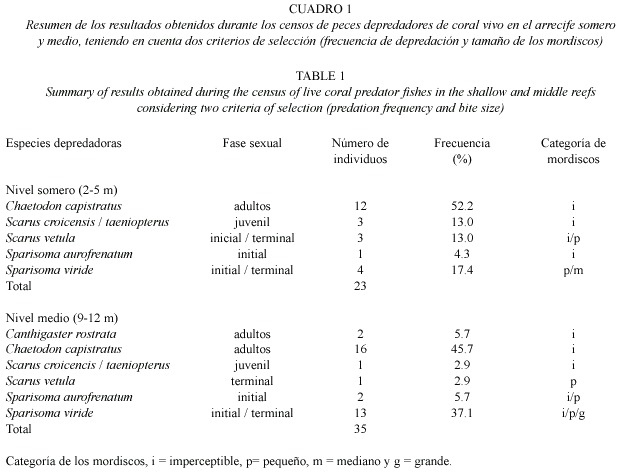
Especies de peces depredadoras de coral:En el nivel somero se observaron 23 individuos de seis especies de peces mordiendo el coral vivo,siendo los adultos de Chaetodon capistratus Linnaeus 1758,los más frecuentes mordedores (52.2%)de tejido coralino, seguidos por adultos de las fases terminal e inicial de S.viride (17.4%)(1).En el nivel medio se hallaron 35 individuos de seis especies de peces (Cuadro 1)mordiendo el coral,incluyendo a Canthigaster rostrata (Bloch,1782) -especie no registrada para el nivel somero-. Nuevamente los adultos C.capistratus presentaron la frecuencia más alta de mordisqueo con 45.7%,seguidos por adultos de S.viride de la fase inicial principalmente (37.1%). Para el nivel profundo se registraron eventos de mordisqueo ocasionados por siete individuos adultos de C.capistratus (frecuencia del 100%). En cuanto al tamaño de los mordiscos,solo tres especies dejaron huellas en los tres niveles de profundidad;S.vetula y S.aurofrenatum (Valenciennes, 1840)presentaron mordiscos desde imperceptibles hasta pequeños ( 5 mm), mientras que S.viride desde imperceptibles hasta grandes ( 15 mm)(Cuadro 1).Debido a sus importantes frecuencias de mordisqueo y al tamaño de los mordiscos,se considera que S.viride (adultos de la fase inicial y terminal) es la principal especie depredadora de tejido coralino en el nivel medio de profundidad en los sitios estudiados.Los corales depredados por estos peces no solo presentaron remoción del tejido,sino también de la capa superficial del esqueleto calcáreo.Aunque C.capistratus se destacó por la frecuencia de mordisqueo,sus mordiscos fueron siempre imperceptibles.
]]>
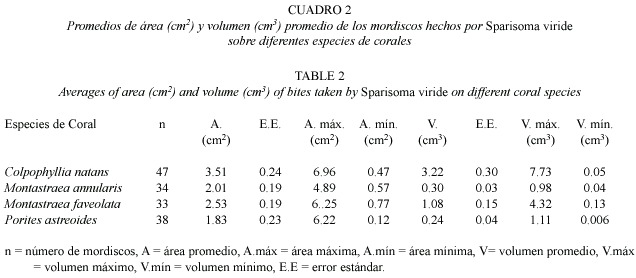
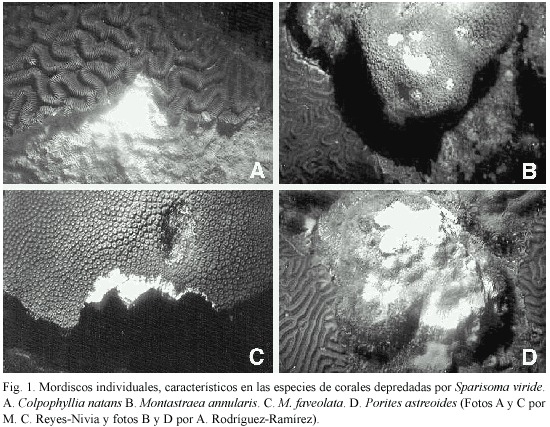
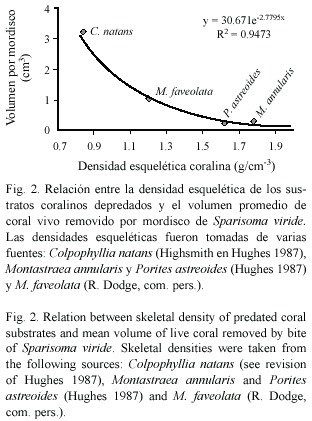
Características de los mordiscos de S. viride :En cuanto al tamaño de los mordiscos individuales,el coral C.natans presentó el mayor promedio tanto de área como de volumen removido por mordisco, con 3.51 cm2 y 3.22 cm 3 respectivamente (Cuadro 2).El tamaño máximo de un mordisco se registró igualmente en esta especie de coral,con un área total de 6.96 cm2 y un volumen de 7.73 cm3, mientras que los mordiscos más pequeños fueron medidos sobre Porites astreoides Lamarck 1816 con 0.12 cm2 de área y 0.006 cm3 de volumen.El Cuadro 2 agrupa los resultados obtenidos para cada especie de coral depredada y la Figura 1 muestra las características particulares de las cicatrices individuales de las mismas.Se encontró además que el volumen promedio removido por mordisco está relacionado exponencial y negativamente (r2 = 0.94)con la densidad esquelética de los corales depredados por S.viride (Fig.2).La tendencia encontrada sugiere que a menor densidad,S. viride es capaz de remover más volumen por mordisco y que a pesar de una alta densidad podría causar alguna remoción.
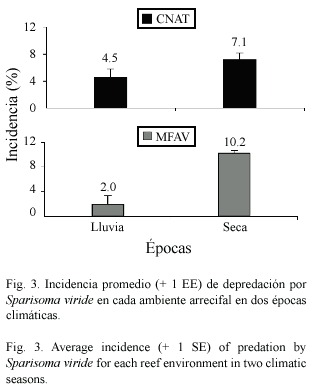
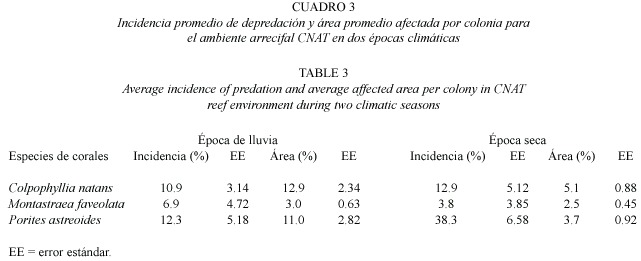
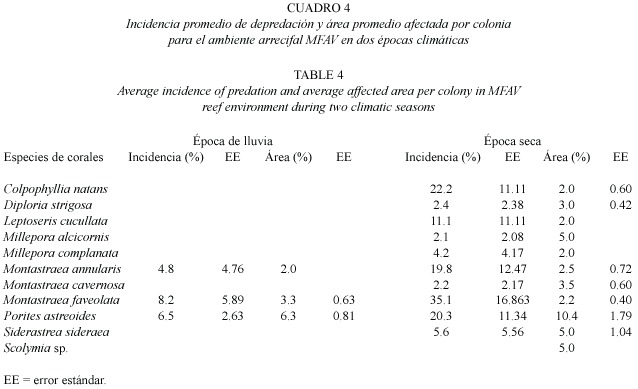
Incidencia de depredación en dos épocas climáticas:Se examinaron en total 722 colonias de 26 especies de corales duros,de las cuales 11 presentaron signos de depredación por S.viride -Colpophyllia natans,Diploria strigosa (Dana, 1846), Helioseris cucullata (Ellis &Solander, 1786), Millepora alcicornis Linnaeus,1758, M.complanata Lamarck,1816 ,Montastraea annularis Ellis &Solander,1786 ,M.cavernosa Linnaeus,1767 ,M.faveolata (Ellis &Solander, 1786),Porites astreoides,Siderastrea siderea (Ellis &Solander,1786)y Scolymia sp.-.Los ambientes de CNAT y de MFAV presentaron la más alta incidencia promedio de depredación durante la época seca,con 7.1 ±1.00 %y 10.2 ±0.48%respectivamente (Fig.3).Sin embargo, al comparar los promedios entre las épocas,se encontró que solo en el ambiente MFAV hay diferencias significativas (p=0.001; t-Student) en la incidencia de depredación.Por especies de corales,se observó que en las bandas de CNAT (tanto en la época lluviosa como en la seca)las especies depredadas fueron C.natans , M.faveolata y P.astreoides ;ésta última presentó las mayores incidencias de depredación: 38.3 ±6.58%durante la época seca y 12.3 ±5.18%en la de lluvias (Cuadro 3).Además, P.astreoides registró diferencias significativas en la incidencia entre ambas épocas (p=0.02; t-Student).En términos de superficie afectada, en el ambiente CNAT se observó una mayor área promedio de tejido depredado durante la época de lluvia para todas las especies (Cuadro 3),de las cuales C.natans registró el valor más alto con 12.9 ±2.34%por colonia.En el ambiente MFAV se observaron 11 especies de corales depredadas durante la época seca,de las cuales sólo Montastraea annularis,M.faveolata y P.astreoides presentaron así mismo signos de depredación en la época de lluvia (Cuadro 4). Precisamente estas tres especies,junto con C.natans presentaron las mayores incidencias de depredación durante la época seca en dicho ambiente,siendo M.faveolata la más frecuentemente depredada 35.1 ±16.83%. Durante la época de lluvia la incidencia de las especies depredadas en MFAV fue muy similar y no sobrepasó el 8.2%.Las pruebas t-Student revelaron que no hay diferencias significativas (p> 0.05) entre la incidencia promedio de depredación para las especies comunes a ambas épocas en MFAV.En cuanto al área promedio de coral vivo afectada por colonia en este ambiente,P. astreoides presentó los valores más altos con 6.3 ±0.81%y 10.4 ±1.79%en las épocas de lluvia y seca respectivamente; para las demás especies la superficie afectada de las colonias no excedió el 5%(Cuadro 4).
]]>
Relación entre la incidencia de depredación y la cobertura de algas filamentosas: Pese a la disminución de la cobertura de los tapetes de filamentosas de la época de lluvias a la seca en ambos ambientes,las pruebas t- Student no revelaron diferencias significativas (p>0.05);en CNAT el porcentaje pasó de 87% (lluvia)a 71.8%(seca)y en el ambiente MFAV varió de 84.9%(lluvia)al 72.4%(seca).En consecuencia,no se encontró relación entre incidencia de depredación y la cobertura de algas filamentosas,la cual fue confrontada por las correlaciones de Spearman que no mostraron coeficientes significativos (todas las probabilidades>0.05)para los ambientes arrecifales en ambas épocas climáticas.
Discusión
Peces depredadores de corales en las Bahías de Chengue y Gayraca:Con base en su comportamiento alimentario,ciertas especies de la familia Chaetodontidae han sido reconocidas como predominantemente coralívoras (Reese 1977,1981).Durante un estudio llevado a cabo en algunas bahías del Parque Nacional Natural Tayrona,Rivera (1991)reconoció que C .capistratus presentaba un régimen alimentario dirigido básicamente hacia los pólipos de corales hermatípicos, octocorales y pequeños invertebrados asociados a las superficies duras de los corales.Birkeland y Neudecker (1981) describieron a C.capistratus como un "generalista ramoneador "de los pólipos coralinos.
Así mismo,durante este trabajo,se encontró que en las bahías de Chengue y Gayraca, C.capistratus presentó un comportamiento alimentario similar,derivado de su frecuente mordisqueo sobre diferentes especies de corales en los tres niveles de profundidad examinados.Los eventos de depredación registrados para esta especie,nunca lograron exponer el esqueleto calcáreo de los corales depredados, por lo tanto,visualmente no se registró ningún tipo de alteración de la superficie coralina.Se ha encontrado que estos de coralívoros sólo consumen partes suaves del coral,sin llegar a destruir o desnudar el esqueleto calcáreo debido a que extraen delicadamente los pólipos individuales (Glynn 1990,Hixon 1997).
]]>
A pesar de que los peces loro son principalmente consumidores de algas bentónicas (Choat 1991),la mayoría de especies observadas en este estudio también depredaron directamente el coral vivo.En un estudio de contenido estomacal de peces arrecifales,Randall (1967) informó que S.aurofrenatum fue el único escárido que presentó fragmentos de coral vivo.Más adelante,Frydl (1979)observó que S.aurofrenatum y S.taeniopterus tomaban ocasionales mordiscos en los ápices vivos de Madracis mirabilis (Duchassaing &Michelotti, 1860).En otro estudio,S.aurofrenatum fue descrito como uno de los mayores depredadores de coral,atacando específicamente fragmentos de Porites divaricata Lesueur,1820 (Miller y Hay 1998).En contraste,la dieta alimentaria de S.croicensis está constituida principalmente por sedimentos que toma del fondo y ocasionalmente algas filamentosas e incrustantes (Ogden y Buckman 1973).Las observaciones de campo en el Parque Tayrona,sugieren que tanto S .aurofrenatum como S.croicensis pastorean constantemente sobre tapetes de algas filamentosas,mientras que la depredación de coral vivo puede presentarse de manera incidental.Es decir,debido a que los sustratos de pastoreo (coral muerto)están muy cerca de la superficies coralinas vivas o son parte de las mismas colonias,el tejido vivo puede ser fácilmente removido de manera accidental.
Las especies S.viride y S.vetula ,que dejaron claras cicatrices de remoción del tejido coralino,han sido los escáridos más comúnmente mencionados por sus actividades de depredación sobre coral vivo (Frydl y Stearn 1978, Frydl 1979,Bruggemann et al.1994a,1996). Las actividades de depredación frecuentes y las conspicuas cicatrices de mordisqueo por parte de S.viride ,lo señalan como el depredar más significativo de coral vivo en el Parque Natural Tayrona y está acorde con lo encontrado en otros estudios.Frydl (1979)concluyó que la depredación importante de coral vivo estaba limitada a los adultos de las dos especies antes mencionadas,pero que S.viride podía alimentarse más extensivamente del tejido vivo.Frydl y Stearn (1978) señalaron así mismo que S.viride es la única especie de loro que deja cicatrices bien definidas al depredar el coral vivo. Bruggemann et al.(1994b)encontraron que aunque S.vetula y S.viride en general evitan el coral vivo como sustrato de forrajeo, este último toma mordiscos más a menudo que S.vetula .Más aún,Miller y Hay (1998)observaron que colonias coralinas completas eran consumidas por S.viride ,y lo consideran el principal depredador de coral.
En cuanto a la depredación por niveles de profundidad,se logró establecer que S.viride presentó la mayor frecuencia en el nivel medio (9-12 m),donde precisamente está el mejor desarrollo coralino de los arrecifes examinados. Similarmente,algunos estudios demuestran que la depredación de coral vivo no abarca la completa distribución vertical de los corales (Bruggemann et al.1994a,Bruckner y Bruckner 1998a)y que esencialmente la mayor frecuencia de mordisqueo se concentra en las zonas arrecifales de alta diversidad y cobertura de coral vivo (Bruggemann et al. 1994a,1994b).
Características de los mordiscos:La depredación de S.viride produjo una cantidad importante de conspicuos mordiscos individuales en el área arrecifal examinada.Tales cicatrices presentaron características particulares en cuanto a la cantidad de tejido removido, dependiendo de la especie de coral afectada. Bruckner y Bruckner (2000)denominaron a este tipo de depredación como "mordisqueo puntual ",ya que son mordiscos tomados individualmente y de manera azarosa por grupos errantes de S.viride y otros peces loro al raspar la superficie de los corales.
Debido a la escasa profundidad encontrada en las cicatrices de P.astreoides ,esta especie presentó el menor volumen promedio (0.24 cm3)removido por mordisco similar a lo informado por Frydl (1979). Por otro lado,M.annularis fue intensamente depredada acorde con la mayoría de informes consultados (Frydl 1979, Bythell et al.1993,Bruggemann et al.1994a). Cabe mencionar que dicha especie presenta en el área de estudio coberturas relativas bajas (2.1%)y se podría considerar que los mordiscos y las lesiones (mordiscos agregados en una misma área)dejadas por la depredación de S. viride tendrían una alta significancia para este coral.No obstante,observaciones de campo (no cuantificadas)evidenciaron una recuperación parcial de los mordiscos individuales en M.annularis .De igual forma,algunos estudios han comprobado que lesiones pequeñas en esta especie de coral pueden regenerarse en gran proporción,pero no logran cerrarse completamente (Bak et al.1977, Meesters y Bak 1994, Meesters et al.1994).Por lo tanto,algunos factores como el tamaño de las lesiones,la capacidad de regeneración de los corales -la cual es variable entre especies-y la colonización de organismos determinan en gran medida el proceso de regeneración coralina (Bak et al. 1977,Meesters et al.1994).
Recientemente M.faveolata ha sido reconocida como una de las principales especies depredadas (Bruckner y Bruckner 1998a, 2000),probablemente debido a que en estudios anteriores no se discriminaban las especies del complejo Montastraea annularis (Weil y Knowlton 1994).En el arrecife examinado,M.faveolata es una especie dominante y presentó una considerable remoción por mordisco, principalmente en términos de área (2.53 cm2).C.natans ha sido señalada como una especie frecuentemente depredada por S. viride (Bruckner y Bruckner 1998a,1998b, 2000)y en este estudio se destacó además por la profundidad de los mordiscos dejados por S.viride ;en promedio,C.natans presentó el mayor volumen removido por mordisco (3.22 cm3)e igualmente en área promedio registró la más alta remoción (3.51 cm2).
Relación entre la densidad del esqueleto coralino y el volumen removido por mordisco:La relación exponencial resultante entre estas dos variables sugiere que a pesar de una alta densidad del esqueleto coralino,S.viride es capaz de remover el tejido vivo.El esqueleto de los corales escleractinios tiene un complejo de poros que determina su densidad,la cual varía interespecíficamente y está determinada en parte por las diversas formas de crecimiento de los corales (Hughes 1987). Por ejemplo los corales masivos son en general el grupo morfológico menos denso,del cual C.natans presenta una densidad menor a 1g cm-3,a diferencia la forma masiva de P.astreoides tiene uno de los esqueletos más densos de los corales con este crecimiento (Hughes 1987). Weil y Knowlton (1994)reconocieron que entre las especies del complejo Montastraea annularis ,M faveolata es siempre menos densa.Teniendo en cuenta tales diferencias en las especies de corales depredadas,se logró establecer que la densidad coralina determina en gran medida la cantidad de tejido vivo y esqueleto calcáreo que puede ser removido por S.viride y por consiguiente C. natans es la especie principalmente afectada.
Estacionalidad en la incidencia de depredación:Hasta ahora no existen informes acerca de si la depredación por S.viride está concentrada durante una época del año y relacionada con particulares características del ambiente.Si bien se encontró una tendencia en ambos ambientes a presentar mayor incidencia de depredación durante la época seca,hay ciertos aspectos que no permiten ser concluyentes con respecto a la estacionalidad en la escala de los ambientes estudiados.Únicamente en el ambiente MFAV la incidencia de depredación fue significativamente mayor durante la época seca como resultado de un aumento generalizado en la frecuencia de depredación de las especies comunes a ambas épocas,así como la aparición de nuevas especies mordidas.Las observaciones de campo permiten aducir que el mayor número de especies mordidas pudo ser accidental y no corresponder a una interacción de depredación.Esto porque la mayoría de las lesiones fueron pequeños mordiscos encontrados entre los límites del tejido vivo de los corales y los tapetes de algas filamentosas y porque fueron registradas sobre corales que raramente son depredados.De otro lado,el área promedio afectada solo presentó variaciones en P.astreoides ,siendo mayor en la época seca.Si bien las variaciones en la abundancia de las poblaciones de S.viride podrían estar relacionadas con la aparente estacionalidad en la incidencia de depredación entres las dos épocas evaluadas,estudios importantes realizados sobre las densidades de escáridos en el Caribe no han encontrado una variación temporal de sus poblaciones aún durante períodos que implican cambios climáticos (Lewis y Wainwright 1985,Bruggemann et al.1994c).Así mismo nuestras observaciones de campo por más de diez años no solo en las estaciones de la bahía de Chengue sino a lo largo del PNN Tayrona sugieren que las poblaciones de peces loro no varían temporalmente.En contraste,el ambiente de CNAT no presentó diferencias significativas en la incidencia y su tendencia al aumento en la incidencia durante la época seca,estuvo únicamente relacionado con la contribución de P.asteroides ,que además presentó mayor área promedio afectada pero en la época lluviosa.Por consiguiente,no se puede hacer una generalización con relación a la estacionalidad de depredación para los ambientes estudiados. Al nivel de las especies,los casos de P.astreoides y de la depredación incidental,merecen particular atención y por lo tanto mayor detalle de investigación,porque las explicaciones a las tendencias observadas no se pudieron extraer de la información tomada -cobertura de algas filamentosas,ver adelante-y podrían aportar información para comprender mejor el comportamiento de depredación en una escala mayor y determinar si efectivamente existe un patrón de estacionalidad.
Las algas filamentosas presentaron un patrón de cobertura similar durante las dos épocas y al correlacionar la incidencia de depredación de S.viride con dichas coberturas -por ser su alimento preferencial (Bruggemann et al. 1994a, 1994b, 1996)-para los dos ambientes arrecifales en ambas épocas,no se encontró dependencia entre ambas variables.Esto indicaría que la variación hallada en la incidencia de depredación entre las épocas no estaría relacionada con los tapetes de algas filamentosas,cuyas coberturas se mantienen relativamente estables tanto en el ambiente CNAT como MFAV (época de lluvia=87%y 84.9% respectivamente;época seca=71.8%y 72.4% respectivamente).
Es importante resaltar que la depredación de coral vivo es un factor clave que controla en parte la organización de las comunidades arrecifales (Glynn et al.1972,Glynn 1988, 1990)y ha sido reconocida como una forma incesante de perturbación biológica (Bythell et al.1993).No obstante,al comparar con los impactos catastróficos esporádicos (tormentas, huracanes,grandes eventos de blanqueamiento y la consecuente muerte asociada),es muy poco lo que se conoce acerca de los efectos de la mortalidad cotidiana del tejido coralino en los arrecifes.Los resultados del presente estudio demuestran que en general la depredación de los corales por parte de los peces y en especial de
]]>
S.viride ,es un suceso habitual en los arrecifes estudiados.Por lo tanto las pérdidas del tejido coralino podrían tener importantes consecuencias para la estructura de éstos arrecifes (composición y abundancia de corales)y su salud, teniendo en cuenta que actualmente son afectados también por otros agentes de mortalidad de origen natural y antrópico.
Agradecimientos
Este trabajo se realizó gracias al apoyo financiero del Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología Francisco José de Caldas (Proyecto Colciencias -BID 2105-09-327-97)y al apoyo financiero y logístico del Instituto de Investigaciones Marinas y Costeras (INVEMAR),en el marco del proyecto "Sistema de monitoreo para los ambientes arrecifales en Colombia y evaluación de agentes de mortalidad coralina ".También un agradecimiento especial a Nadiezdha Santodomigo y a Eduardo Vilarete por su valiosa colaboración en el campo.Contribución Nº 724 del INVEMAR.
Resumen
Para estudiar uno de los agentes biológicos implicados en la mortalidad coralina en dos bahías del Parque Nacional Natural Tayrona (Caribe colombiano),se evaluó la depredación de coral vivo por peces mediante censos visuales y se determinó la incidencia de depredación en dos ambientes arrecifales (uno dominado por Colpophyllia natans y otro por Montastraea faveolata ) para dos épocas climáticas (lluvia y seca),examinando todas las colonias presentes en bandas permanentes (20 x 2 m).Sparisoma viride fue el depredador más importante por su frecuencia de mordisqueo y por el tamaño de los mordiscos.Los adultos de las fases terminal e inicial de S.viride lograron remover tejido vivo y parte de la matriz calcárea de M. faveolata, M.annularis ,Porites astreoides y C.natans , de las cuales esta última perdió mayor cantidad de tejido en área (3.51 cm2)y volumen (3.22 cm3)por mordisco. Al relacionar la densidad coralina (gr/cm3)de las especies depredadas con el volumen promedio removido por mordisco,se encontró una tendencia exponencial negativa (r2= 0.94),indicando que el alcance de los mordiscos está determinado por la densidad de los corales. Además,se estableció que no hay una clara relación entre en la incidencia de depredación y las estaciones climáticas en los ambientes estudiados.En las bahías de Chengue y Gayraca,la depredación de coral vivo es uno de los factores que está contribuyendo con la pérdida de tejido coralino y podría tener importantes consecuencias en la dinámica de estos arrecifes.
Bak,R.P.M.,J.J.W.M.Brouns &F.M.L.Heys.1977. Regeneration and aspects of spatial competition in the scleractinian corals Agaricia agaricites and Montastraea annularis .Proc.3rd Int.Coral Reef Symp.,Miami 1:143-148. [ Links ]
Bellwood,D.R &J.H.Choat.1990.A functional analysis of grazing in parrotfishes (Family Scaridae):The Ecological Implications.Env.Biol.Fish.28:189-214. [ Links ]
Birkeland,C.1997.Life and death of coral reefs.Chapman &Hall,Nueva York.536 p. [ Links ]
Birkeland,C.&S.Neudecker.1981.Foraging behavior in two Caribbean Chaetodontids:Chaetodon capistratus and C.aculeatus .Copeia 1:169-178. [ Links ]
Bruckner,A.W.&R.J.Bruckner.1998a.Destruction of coral by Sparisoma viride .Coral Reefs 17:350. [ Links ]
Bruckner,A.W.&R.J.Bruckner.1998b.Rapid wasting syndrome or coral predation by Stoplight Parrotfish?. Reef Encounter 23:18-22. [ Links ]
Bruckner,A.W.&R.J.Bruckner.2000.Parrotfish predation on live coral:"spot biting "and "focused biting ". Coral Reefs 19:50. [ Links ]
Bruggemann,J.H.,M.J.H.Van Oppen &A.M.Breeman. 1994a.Foraging by the Stoplight Parrotfish Sparisoma viride .I.Food selection in different,socially determined habitats,p.29-52.In J.H.Bruggemann (ed.). Parrotfish grazing on coral reefs:A trophic novelty. Ponsen &Looijen,Holanda. [ Links ]
Bruggemann,J.H.,M.Kuiper &A.M.Breeman.1994b. Comparative analysis of foraging and habitat use by the sympatric Caribbean Parrotfish Scarus vetula and Sparisoma viride (Scaridae),p.105-130.In J.H. Bruggemann (ed.).Parrotfish grazing on coral reefs: A trophic novelty.Ponsen &Looijen,Holanda. [ Links ]
Bruggemann,J.H.,A.M.Van Kessel,J.M.Van Roiji &A.M.Breeman.1994c.A parrotfish bioerosion model: implications of fish size,feeding mode and habitat use for the destruction of reef substrates, p.131-152.In J.H.Bruggemann (ed.).Parrotfish grazing on coral reefs:A trophic novelty.Ponsen & Looijen,Holanda. [ Links ]
Bruggemann,J.H.,A.M.Van Kessel,J.M.Van Roiji &A.M.Breeman.1996.Bioerosion and sediment ingestion by the Caribbean Parrotfish Scarus vetula and Sparisoma viride :Implications of fish size, feeding mode and habitat use.Mar.Ecol.Progr.Ser. 134:59-71. [ Links ]
Bythell,J.E.,E.H.Gladfelter &M.Bythell.1993.Chronic and catastrophic natural mortality of the three common Caribbean reef corals.Coral Reefs 12:143-152. [ Links ]
Choat,J.H.1991.The Biology of herbivorous fishes on coral reefs,p.120-155.In P.F.Sale (ed.).The ecology of fishes on coral reefs.Academic.San Diego. [ Links ]
Díaz-Pulido,G.&J.Garzón-Ferreira.2002.Seasonality in algal assemblages on upwelling-influenced coral reefs in the Colombian Caribbean.Bot.Mar.45: 284-292. [ Links ]
Frydl,P.1979.The effect of parrotfishes (Scaridae)on coral in Barbados,W.I.Int.Rev.Ges.Hydrob.64: 737-748. [ Links ]
Frydl,P &C.W.Stearn.1978.Rate of bioerosion by parrotfish in Barbados reef environments. J. Sed. Petrol 48:1149-1158. [ Links ]
García,A.,A.Croquer &S.M.Pauls.2003.Estado actual de las enfermedades y otros signos de deterioro coralino en siete arrecifes del Parque Nacional Archipielago de Los Roques,Venezuela.Rev.Biol. Trop.51(Sup.4):173-180. [ Links ]
Glynn,P.W.1988.Predation on coral reefs:some key process,concepts and research directions.Proc.6th Int. Coral Reef Symp.Townsville 1:51-62 [ Links ]
Glynn,P.W.1990.Feeding ecology of selected coral-reef macroconsumers:patterns and effects on coral community structure,p.365-400.In Z.Dubinsky (ed.). Ecosystems of the world 25:Coral Reef. Elsevier, Amsterdam. [ Links ]
Glynn,P.W.1997.Bioerosion and coral-reef growth:a dynamic balance,p.68-95.In C.Birkeland (ed.).Life and Death of Coral Reefs.Chapman,Nueva York. [ Links ]
Glynn,P.W.,R.H.Stewart &J.E.Mccosker.1972.Pacific coral reefs of Panamá:structure,distribution and predators. Geol. Rundschau 61:483-519. [ Links ]
Guzmán,H.M.&J.D.López.1991.Diet of the corallivorous pufferfish Arothron meleagris (Pisces: Tetraodontidae)at Gorgona Island,Colombia.Rev. Biol.Trop.39:203-206. [ Links ]
Hay,M.E.1984.Patterns of fish and urchin grazing on Caribbean coral reefs:are previous results Typical? Ecology 65:446-454. [ Links ]
Hay,M.E.1985.Competition between herbivorous fishes and urchins on Caribbean reefs.Oecologia 65: 591-598. [ Links ]
Hixon,M.A.1997.Effects of reef fishes on corals and algae,p.230-248.In C.Birkeland (ed.).Life and Death of Coral Reefs.Chapman.Nueva York. [ Links ]
Hubbard,D.K.1997.Reefs as dynamic systems,p.43-67. In C.Birkeland (ed.).Life and Death of Coral Reefs. Chapman. Nueva York. [ Links ]
Hughes,T.P.1987.Skeletal density and growth form of corals.Mar.Ecol.Prog.Ser 35:259-266. [ Links ]
Lewis,S.M.1986.The role of herbivorous fishes in the organization of a Caribbean reef community.Ecol. Monogr.56:183-200. [ Links ]
Lewis,S.M.&P.C.Wainwrigth.1985.Herbivore abundance and grazing intensity on a Caribbean Coral Reef.J. Exp. Mar. Biol. Ecol.87:215-228. [ Links ]
Meesters,E.H.&R.P.M.Back.1994.Effects of coral bleaching on tissue Regeneration potential and colony survival,p.27-41.In E.H.Meesters (ed.).The function of damage and regeneration in the Ecology of reef-building corals (Scleractinia). Proefschrift Universiteit van Amsterdam,Holanda. [ Links ]
Meesters,E.H.,M.Noordeloos &R.P.M.Back.1994. Damage and regeneration:links to growth in the reef building coral Montastraea annularis .p.43- 72.In E.H.Meesters (ed.).The function of damage and regeneration in the ecology of reef-building corals (Scleractinia).Proefschrift Universiteit van Amsterdam,Holanda. [ Links ]
Miller.W.M.&M.E.Hay.1998.Effects of fish and seaweed competition on the survival and growth of coral.Oecologia 113:231-238. [ Links ]
Ogden,J.C.&N.S.Buckman.1973.Movements foraging groups,and diurnal migrations of the striped parrotfish Scarus croicensis Bloch (Scaridae).Ecology 54:589-596. [ Links ]
Randall,J.E.1967.Food habits of reef fishes of the West Indies.Stud.Trop.Oceanogr.5:665-847. [ Links ]
Randall,J.E.1974.The effect of fishes on coral reefs.Proc. 2nd Int.Coral Reef Symp.1:159-166. [ Links ]
Rivera,G.1991.Ecología de los peces de las familias Chaetodontidae y Pomacanthidae en la región de Santa Marta Caribe colombiano.Tesis de pregrado, Universidad de Antioquia.Medellín,Colombia. [ Links ]
Reese,E.S.1977.Coevolution of Corals and Coral Feeding Fishes of the Family Chaetodontidae.Proc.3rd Int. Coral Reef Symp.Miami 1:267-274. [ Links ]
Reese,E.1981.Predation on corals by fishes of the family Chaetodontidae:Implications for conservation and management of coral reef ecosystems.Bull.Mar.Sci. 31:594-604. [ Links ]
Toro-Farmer,G.,J.R.Cantera K.,E.Londoño-Cruz,C. Orozco &R.Neira O.2004.Patrones de distribución y tasas de bioerosión del erizo Centrostephanus coronatus (Diadematoidea:Diadematidae),en el arrecife de Playa Blanca,Pacífico colombiano.Rev.Bio. Trop.52:67-76. [ Links ]
Wanders,J.B.M.1977.The Role of benthic algae in the shallow reef of Curaçao (Netherlands Antilles)III: the significance of grazing.Aquat.Bot.3:357-390. [ Links ]
Weil,E.&N.Knowlton.1994.A Multi-character analysis of the Caribbean coral Montastraea annularis (Ellis and Solander, 1786)and its two sibling species,M. faveolata y M.franksi (Gregory,1895).Bull.Mar. Sci.55:151-175. [ Links ]
Zubillaga,A.L.,S.M.Pauls &A.Croquer.2003.Evaluación de las actividades del buceo recreativo sobre la estructura comunitaria de algunos arrecifes del Parque Nacional Archipielago de Los Roques, Venezuela. Rev. Biol. Trop. 51 (Sup.4):189-195. [ Links ] ]]>19771143-148199028189-214199753619811169-1781998a173501998b2318-22200019501994a29-521994b105-1301994c131-152199613459-71199312143-1521991120-155200245284-292197964737-7481978481149-1158200351^sSup.4^sSup.4Sup.4173-1801988151-621990365-400199768-95197261483-519199139203-206198465446-454198565591-5981997230-248199743-67198735259-26619864239-252198656183-200198587215-228199427-41199443- 721998113231-238197354589-59619675665-84719741159-166199119771267-274198131594-60420045267-7619773357-390199455151-175200351^sSup.4^sSup.4Sup.4189-195
 5 mm),mediano (6-14 mm)y grande (
5 mm),mediano (6-14 mm)y grande ( 15 mm);esto para estimar de una manera simple y rápida el grado de afección por mordisco.Se evaluaron dos estaciones en Chengue y tres en Gayraca en los niveles de profundidad (somero=2-5 m,medio=9-12 m y profundo=16-19 m)presentes en cada estación y se efectuaron al menos tres censos por nivel de profundidad.
15 mm);esto para estimar de una manera simple y rápida el grado de afección por mordisco.Se evaluaron dos estaciones en Chengue y tres en Gayraca en los niveles de profundidad (somero=2-5 m,medio=9-12 m y profundo=16-19 m)presentes en cada estación y se efectuaron al menos tres censos por nivel de profundidad.