 ]]>
]]>
Diversidad y abundancia de moluscos en las praderas de Thalassia testudinum de la Bahía de Mochima, Parque Nacional Mochima, Venezuela
Antulio Prieto 1, Sybil Sant 1, Elizabeth Méndez 1 & César Lodeiros 2
Recibido 13-XI-2001. Corregido 07-V-2002. Aceptado 11-IX-2002
The diversity and abundance of benthic malacological communities associated to Thalassia testudinum beds was studied at four localities of Mochima Bay, Sucre state, Venezuela. At each locality, samples were taken monthly on perpendicular transect at different depths (0-4 m), from January 1991 to December 1991, using a quadrate (0.25 m2 ) for collecting mollusks and sediments. A total of 2 988 organisms of infauna and epifauna belonging to 81 species of the classes Gastropoda (41) and Bivalvia (40) were collected. More abundant species were Anadara notabilis, Codakia orbicularis, Cerithium litteratum, Cerithium eburneum, Batillaria minima, Modiolus squamosus, Modulus modulus, Chione cancellata, Turritella variegata, Arca zebra, y Laevicardium laevigatum. There were significant differences in number of organisms between depth and month at La Gabarra which presented the highest value of total (4.51 bits/ind) and monthly diversity (2.713.90 bits/ind). Biomass and abundance were low in the Mochima Bay while Varadero station presented the highest value. The bivalve A. notabilis and gastropod M. modulus were species common to the four stations.
Key words: benthic community, molluscs, seagrass beds, Mochima, Venezuela, Thalassia.
]]>En Venezuela, la mayoría de los estudios sobre la fauna asociada se han orientados sobre aspectos taxonómicos (Flores 1968, Almeida 1974, Prinz 1978) y ecológicos (Hambrook et al. 1979, Bitter 1993, 1988, 1999, Rebolledo et al. 1993, Prieto et al. 2003).
La Bahía de Mochima es una zona con gran actividad turístico-pesquero en donde las praderas de T. testudinum están ampliamente distribuidas, albergando comunidades particulares donde los moluscos son los organismos mas relevantes. Sobre la zona destaca solamente los trabajos realizados por Jiménez (1994) y Jiménez et al. (2000), quienes analizaron los moluscos asociados a la fanerógama en la Ensenada de Reyes, indicando la presencia de unas 53 especies. El propósito del presente estudio es describir la estructura de la comunidad de moluscos en cuatro localidades de la Bahía de Mochima, analizando específicamente aspectos como diversidad, riqueza específica, abundancia, biomasa, constancia y dominancia.
Materiales y métodos
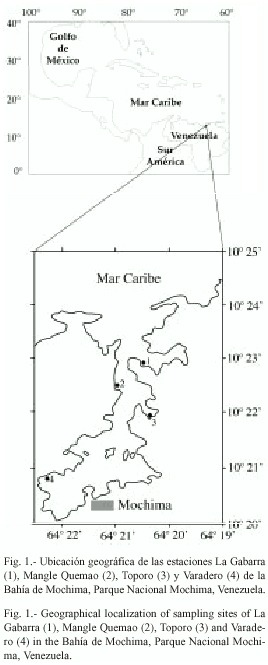
Área de estudio: La Bahía de Mochima está ubicada en la costa nororiental de Venezuela, a 30 km al oeste de la ciudad de Cumaná, entre los 10º2430" y 10º20 N y los 64º1930" y 64º2230" W. (Fig. 1). Constituye un cuerpo de agua largo y semiestrecho, con su eje mayor orientado en sentido NE-SW (7 150 m de longitud), con una anchura mínima de 300 m y máxima de 3 400 m. La longitud máxima es de 5 km en dirección norte-sur y de 3 km en sentido este-oeste, ocupando un área de 10.5 x 106 m2 y un volumen de 196 x 106 m3 (Okuda et al. 1968).
Las características físico-químicas del agua han sido estudiadas por Kato (1961), Okuda et al. (1968) y García (1978). Según estos autores, los parámetros estudiados muestran cierta estacionalidad, los cuales, están relacionados con los periodos de sequía y lluvia, coincidente con la mayor actividad de surgencia y la estratificación del agua, respectivamente.
]]>
Características de las localidades: La Gabarra (1) se encuentra cerca de la boca de la bahía, por lo que el oleaje es constante y fuerte. Presenta una pendiente bastante abrupta y a los 40-50 m de distancia de la zona interma-real, alcanza unos 6 m de profundidad. La profundidad promedio es de 4.5 m, el sustrato en mayor proporción está compuesto de arena media, la pradera de T. testudinum está distribuida en forma de parches y se alterna con formaciones rocosas, coralinas y arenosas.
Mangle Quemao (2) está ubicada en la zona central de la Bahía, el oleaje es moderado y constante. La línea de costa es una playa arenosa de 44 m de longitud con presencia de formaciones rocosas, bastante extensas hacia la parte Este. Según Caraballo (1968) el sustrato es arenoso (fracciones media y fina), y está cubierta por una pradera de T. testudinum bien distribuida con una profundidad promedio de 2 m.
Toporo (3), está bordeada por completo del mangle rojo Rhizophora mangle, y se encuentra ubicada en la zona central de la bahía, el oleaje es moderado ya que la ensenada se encuentra resguardada de vientos fuertes. El sustrato está compuesto por sedimentos limoarenosos y está cubierto por T. testudinum muy densa y frondosa aunque se alterna con formaciones del coral Millepora sp. La profundidad promedio es de 2.27 m y el ancho de la zona es de 40 m.
Varadero (4) está ubicada en la parte más interna de la bahía, bordeada por manglares dispersos. El oleaje es constante y moderado. La profundidad promedio es de unos 4 m y el sustrato está compuesto por sedimentos limo-arcillosos (Caraballo 1968), lo que le confiere al fondo un color grisáceo y una textura blanda, características del sustrato fangoso. La pradera de T. testudinum cubre parcialmente el sustrato, conjuntamente con restos de corales muertos.
Métodos de campo: Desde enero de 1991 hasta diciembre del mismo año se realizaron muestreos mensuales en las cuatro localidades de la Bahía de Mochima (Fig. 1). Las colectas se realizaron sobre transeptos de 50 m de longitud por 2 m de ancho desde 0 hasta 4 m. Cada transepto se dividió en diez cuadrantes de 5 m separados por listones de madera y sujetos con pesos en cada extremo. Además, se usaron dos cuadratas metálicas de 0.25 m2 y tamices con 1 mm de abertura de orificio. Todos los transeptos se colocaron perpendicularmente a la costa, realizando observaciones sobre las características del substrato, las cuadratas fueron colocadas al azar en cada cuadrante, tomándose muestras de 0.25 m2 , removiendo el sedimento hasta unos 10 cm con la ayuda de palas metálicas utilizando un equipo de buceo autónomo. La profundidad se determinó con un profundímetro manual. Todas las muestras recolectadas fueron guardadas en bolsas plásticas, para su posterior análisis y fueron depositadas en la colección del Grupo de Biología de Moluscos del Consejo de Investigación de la Universidad de Oriente.
En cada localidad se determinó la salinidad con un refractómetro (±1), temperatura y oxígeno disuelto con un oxigenómetro, pH con un pH-metro de campo y la transparencia del agua con un disco de Secchi.
Análisis de los datos: Las muestras se almacenaron en un refrigerador y se identificaron con la ayuda de una lupa estereoscópica y microscopio de luz siguiendo las recomendaciones de Abbott (1974), Humphrey (1975) y Lodeiros et al. (1999). El peso y tamaño de los individuos fueron determinados con una balanza y vernier digital de 0.01 g y 0.01 mm de apreciación, respectivamente.
Para interpretar la estructura ecológica se determinaron la diversidad, utilizando la relación de Shannon-Wiener, el Índice de Simpson, el Número de Hill (N2) y la equitabilidad, así como la similaridad por el índice de Morisita, siguiendo las recomendaciones en Krebs (1989). Por otra parte, se determinó la constancia según la relación C = P * 100/p, donde P es el número de muestreos donde aparece la especie y p el número total de muestreos realizados. De igual manera, se determinó la biomasa total de los individuos y la densidad por área de muestreo.
Para establecer diferencias significativas en cada una de las localidades se realizó un análisis de varianza, considerando como factores la profundidad y el tiempo de muestreo. En aquellos factores donde se establecieron diferencias significativas, se aplicó la prueba de Scheffé, siguiendo las recomendaciones de Sokal y Rohlf (1979). Para todos los casos, el nivel de significancia fue de 0.05.
]]> ResultadosNúmeros y composición de especies: Se colectó un total de 2 988 individuos pertenecientes a 81 especies de las clases Gastropoda y Bivalvia. Dentro de la Clase Bivalvia se obtuvo un total de 41 especies pertenecientes a 29 géneros y 19 familias. Dentro de la clase Gastropoda se colectaron un total de 40 especies pertenecientes a 30 géneros y 25 familias. Las familias mejor representadas para la primera clase fueron: Arcidae, Lucinidae y Veneridae, y para la segunda Turbinidae, Muricidae y Olividae (Cuadro 1).

En la estación 4, se colectaron 1504 individuos repartidos en 50 especies, las más abundantes fueron Cerithium litteratum (703), Anadara notabilis (403), Cerithium eburneum (140) y Chione cancellata (68). En la estación 2, se colectaron 421 individuos repartidos en 46 especies, las más abundantes fueron A. notabilis (125) y Modiolus americanus (121). En la estación 3, se obtuvieron 602 individuos repartidos en 41 especies, las más abundantes fueron C. litteratum (169) y C. eburneum (107). En la estación 1, se colectaron 478 individuos repartidos en 54 especies, las especies más abundantes fueron Turritella variegata (65), C. litteratum (49), A. notabilis (47) y C. eburneum (45).
Las especies presentes en todas las estaciones fueron las siguientes: A. notabilis, Arca zebra, Arca imbricata, Batillaria minima, C. eburneum, C. litteratum, C. cancellata, Codakia orbicularis, Codakia orbiculata, Corbula contracta, Laevicardium laevigatum, M. americanus, Modiolus squamosus, Modulus modulus, Trachycardium isocardia, Turbo castanea y Turritella variegata.
Abundancia en relación a la profundidad:
El análisis de la abundancia durante el muestreo arrojó diferencias significativas (p<0.001) de las abundancias en las profundidades y entre los meses para la estación 4. En contraste, las estaciones intermedias (2 y 3) no presentaron diferencias significativas (p>0.05) de la abundancia con respecto a la profundidad y el tiempo de muestreo (meses). En el caso de la estación 1, se observaron diferencias significativas (p<0.001) de la abundancia en el tiempo, pero no con la profundidad. La comparación múltiple establecida en el factor tiempo indicó que la abundancia de los meses enero, febrero, marzo, abril y mayo, forman un primer grupo de mayor abundancia, y las que pertenecen a los meses mayo y junio forman un segundo grupo.
]]> En las estaciones (1 y 4) las abundancias en los meses de mayo y abril fueron intermedias a los grupos formados, pero la abundancia de junio no guardó relación con la de los primeros meses. En la estación 4, se obtuvo diferencias altamente significativas de las abundancias en 2.5 m y 3.5 m con respecto a las demás.Biomasa: Los promedios de biomasa mensual para cada localidad no mostraron diferencias significativas, pero si las biomasas totales (p<0.01). En general, las biomasas mensuales fueron muy bajas. En la estación 1 fluctuaron entre 0.92 g/m2 en enero y 6.81 g/m2 en junio. En la estación 2, el valor mínimo osciló entre 1.86 g/m2 obtenido en marzo y 8.49 g/m2 en agosto. En la estación 3, osciló entre 0.82 g/m2 en marzo y 8.49 g/m2 en septiembre; y en la estación 4 entre 2.03 g/m2 en noviembre y 9.67 g/m2 en septiembre (Fig.2a). Las biomasas totales presentaron un valor mínimo en la estación 3 (25.19 g/m2) y el máximo en la estación 4 (64.61 g/m2) (Cuadro 2).
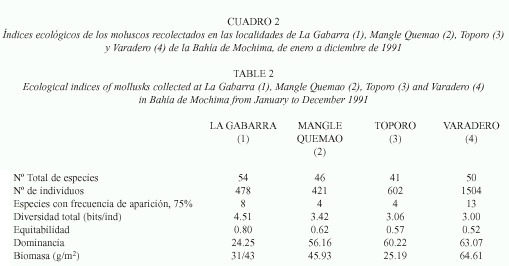
Parámetros comunitarios: La diversidad total (Shannon-Wiener) presentó su valor máximo en la estación 1 (4.51 bits/ind.) y el mínimo en la estación 4 (3.00 bits/ind.) (Cuadro 2). Los valores mensuales de diversidad malacológica presentaron muchas variaciones durante el periodo de estudio, aunque éstas fueron más notables en la estación 3, donde se observó la diversidad más baja en marzo (0.70 bits/ind.). El mayor valor se encontró en la estación 1 (3.81 bits/ind.) (Fig. 2b).
La equitabilidad total osciló entre 0.80 (estación 1) y 0.52 (estación 4) (Cuadro 2). La variación mensual de este parámetro presentó la misma tendencia que la diversidad y el máximo valor (0.86) se obtuvo en Julio para la estación 1, y el menor (0.42) en la estación 3 para mayo (Fig. 2c).
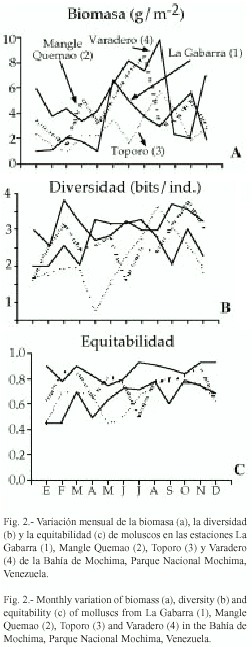
En la estación 4 se observó el mayor número de especies constantes con porcentajes de aparición superiores de 50% (13), seguido de la estación 1 (8) y las estaciones 2 y 3 con cuatro especies respectivamente (Cuadro 3).
El bivalvo A. notabilis y el gasterópodo M. modulus fueron los únicos moluscos presentes en las cuatro localidades. Con relación a la dominancia numérica, C. litteratum fue la especie dominante en las localidades de Varadero (4) y Toporo (3), mientras que A. notabilis y C. cancellata dominaron en Mangle Quemao (2). En la Gabarra (1) las especies dominantes fueron T. variegata, C. litteratum y C. eburneum.
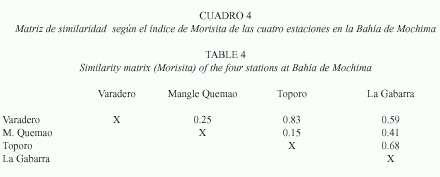
La mayor similaridad entre estaciones según el Índice de Morisita se observó entre las estaciones 3 y 4 (0.83) y el valor mínimo (0.15) entre las estaciones 2 y 3 (Cuadro 4).
Discusión
El número de especies de gasterópodos y de bivalvos asociados a las praderas de T. testudinum en la Bahía de Mochima es elevado con respecto a los trabajos reportados para otras comunidades, tanto en Venezuela, como para otras zonas del Caribe. En la costa nororiental de Venezuela, Vera (1978) informó 56 especies de moluscos. En dos localidades del Golfo de Cariaco, Graterol (1986), señaló un total de 75 especies, en las cuales, 36 fueron bivalvos y 35 gasterópodos. Bitter (1988) en un análisis de la comunidad asociada a la pradera de T. testudinum en el Parque Nacional Morrocoy, reportó 24 especies de macroinvertebrados, de los cuales, 9 especies fueron moluscos. En la misma zona Rebolledo et al. (1993), en un estudio sobre la relación Thalassia- epifauna móvil, encontraron 30 especies de gasterópodos y 8 de bivalvos. En otras áreas del Mar Caribe como en Jamaica, Jackson (1972) registró 74 especies y Howard (1987) en Indian River, Florida, reportó 10 especies de gasterópodos. Bello (1989) en Islas Vírgenes encontró 29 especies de gasterópodos asociados a una pradera de Thalassia. Para la Bahía de Mochima, la mayor cantidad de especies encontradas en este trabajo en relación al de Jiménez et al. (2000) se debe al mayor número de estaciones muestreadas.
Almeida (1974) comparando varios tipos de habitat litorales: manglar, arenoso, rocoso, estuarino, parches coralinas en las praderas de Thalassia, de la región centro-occidental Venezolana, reportó 19 especies de moluscos en el manglar, 13 en el litoral arenoso, 48 especies en litoral rocoso, 23 en el estuarino, 97 en las islas coralinas y 24 en las praderas de Thalassia, citando 12 especies en común con el presente trabajo. Los altos valores encontrados con respecto a datos de áreas del Caribe y otras regiones de Venezuela, pueden deberse a las características de la bahía, la cual presenta una gran variedad de ambientes que proporcionan la existencia de una diversidad de nichos (arenoso, areno-fangoso, formaciones coralinas y rocosas), además, por ser una bahía constituye una zona de refugio y protección para el crecimiento de juveniles de muchas especies. Una hipótesis alternativa es que parte de las diferencias encontradas sean debidas a diferencias en el tipo y tiempo de muestreo utilizado en los trabajo realizados; no obstante, la marcada diferencia en los resultados obtenidos no sustentan dicha argumentación.
Las especies más abundantes y en consecuencia mejor adaptadas a los habitats suministrados por la T. testudinum en la Bahía de Mochima fueron A. notabilis, C. orbicularis, C. litteratum, C. eburneum, B. minima, M. squamosus, M. modulus, C. cancellata, T. variegata, A. zebra, y L. laevigatum. Algunas de estas especies también han resultado numerosas en otras comunidades de Thalassia. Graterol (1986) en Guaracayal (Golfo de Cariaco) reporta a C. orbicularis, C. cancellata, A. notabilis, T. variegata como las predominantes. No obstante, M. modulus y C. cancellata fueron los más abundantes en praderas de Thalassia del Parque Nacional Morrocoy en el occidente de Venezuela (Bitter 1988, Rebolledo et al., 1993). A su vez, Jackson (1972) en Jamaica encontró como especies abundantes a C. litteratum, A. notabilis, C. orbicularis, A. zebra y M. americanus, los cuales, también obtuvieron el mayor número de individuos en este estudio. De igual manera, A. notabilis, C. eburneum, C. litteratum, C. cancellata, M. modulus y L. laevigatum han sido reportadas como las especies más abundantes en la praderas de Thalassia de Cuba (Alcolado 1990); no obstante, M. modulus es la especie predominante en Florida (Howards 1987) y C. litteratum y C. eburneum en las islas Vírgenes Americanas (Bello 1989).
En la estación 4, la abundancia de las especies se relacionó con la profundidad y con el tiempo. Esta estación está caracterizada por tener una pendiente fuerte, y debido a su ubicación los vientos pueden producir un oleaje constante durante casi todo el año, además el fondo contiene una gran cantidad de restos de corales en las zonas más someras. Todo lo anteriormente expuesto condiciona la elevada variación del número de individuos en las profundidades. Durante los meses de mayo y junio se apreciaron diferencias en las abundancias con respecto a los demás meses, lo que se podría explicar por la existencia, durante esos meses, de una mayor cantidad de materia orgánica de origen terrígeno, por efecto de las lluvias.
Las diferencias significativas de las abundancias en los meses en la estación 1 puede deberse a su cercanía a la boca de la bahía, lo que implica una mayor exposición a los vientos más fuertes durante el periodo mayojunio, lo que ocasionaría una remoción del sedimento, además del continuo intercambio con las aguas superficiales de esta zona de la bahía. Las estaciones 2 y 3 de Mangle Quemao y Toporo, presentaron ambientes menos fluctuantes durante el año, quizás debido a su ubicación dentro de la bahía, lo cual produce que el oleaje sea menos intenso; aparte de ello, la profundidad es poco variable y el fondo es más homogéneo, por lo que los valores de abundancia no registraron variaciones significativas con relación a la profundidad y a los meses.
En la estación 4, los resultados indican que la abundancia de especies es mayor en el rango de profundidad entre 2.5 m y 3.5 m, donde la fanerógama alcanza sus máximas coberturas. En general, se puede inferir que este es el mejor habitat dentro de la profundidad a la cual se extiende la pradera de T. testudinum en la zona, lo cual coincide con los datos reportados por Rebolledo et al. (1993) para otras comunidades de Thalassia en el occidente de Venezuela, señalando que la distribución de los organismos asociados depende de la distribución de la fanerógama en la franja costera, indicando que los individuos se ubican en una profundidad preferencial donde están menos expuestos a las condiciones ambientales extremas y donde Thalassia alcanza un mayor desarrollo, proporcionando mayor cantidad de rizomas y de follaje para la fijación, protección y alimentación de los moluscos.
Las altas biomasas obtenidas en junio, julio, agosto y septiembre (Fig. 2a) podrían estar relacionadas con el mayor aporte de sedimentos de origen terrígeno producido en el periodo lluvioso en la Bahía (Okuda et al. 1968). La mayor biomasa total por estación observada en Varadero es baja cuando se compara con las estimada por Jiménez (1994), en la localidad de Bahía de Reyes, ambas situada hacia la parte interna de la bahía. Probablemente estos resultados están condicionados por las características de esta localidad. Varadero es la única estación en este estudio que presenta un aporte importante de materia orgánica de origen terrígeno, además de la acumulación de sedimentos por efecto del transporte de los vientos en dirección este-oeste dentro de la bahía.
]]> Los valores bajos de biomasa encontrados en Toporo, estarían condicionados por su ubicación, resguardada de los vientos y sin aportes terrígenos considerables, además de la presencia de organismos predadores y competidores observados en el transcurso de los muestreos (observación personal); como peces de las familias Lutjanidae, Labridae, Pomasidae y Diodontidae; en efecto, Diodon holocanthus es un predador voraz de moluscos (Rebolledo et al. 1993), y se ha reportado para las estaciones de Mangle Quemao y Toporo (Méndez et al. 1988). También están presentes caracoles pertenecientes a las familias Cymathidae y Muricidae; y corales competidores por el espacio, disminuyendo así la probabilidad de encontrar una elevada biomasa. Los resultados de las pruebas estadísticas, sugiere que las biomasas de Toporo, La Gabarra y Mangle Quemao son similares, mientras que la de Varadero se diferencia de todas, siendo mucho mayor, quizás debido a la mayor disponibilidad de recursos en esta estación.La diversidad tiende a aumentar desde agosto hasta diciembre. Estos resultados podrían explicarse por las condiciones favorables de la Bahía y los procesos reproductivos de los organismos. Durante este período las fuentes de alimento aumentan por la disponibilidad de la materia orgánica de origen terrígeno, estableciéndose una mayor estabilidad del medio, lo cual, propicia un mayor éxito en el desarrollo de las especies, debido a que, en los períodos precedentes (marzo-julio), podrían acontecer la mayor actividad reproductiva en esta área tropical, tal como ha sido indicado por Lodeiros y Himmelman (1994) para el Golfo de Cariaco, área adyacente a la zona de estudio.
Es importante señalar que los resultados de la diversidad podrían estar influenciados por el tipo de muestreo, el cual, a su vez puede variar dependiendo de los hábitos alimenticios y migratorios (diarios e interdiarios) de los moluscos (Howard 1987). Sin embargo, estas fluctuaciones notables de diversidad, y abundancia también han sido observadas en otras comunidades de organismos bentónicos (Bitter 1999). La mayor diversidad total para todos los índices observada en la estación 1 (4.58 bit-s/ ind), es superior al informado por Jackson (1972), con una diversidad máxima de 3.81 bits/ind; por Graterol (1986) en el Golfo de Cariaco (3.53 bits/ind) y por Marvál (1986) en litoral rocoso en la Isla de Margarita cuya diversidad máxima fue de 2.47 bits/ind. La diversidad está afectada por factores ambientales, cuyo gradiente aumenta desde la parte interna hacia la boca de la Bahía. Esta misma característica ha sido señalada para el Golfo de Batabano, Cuba (Alcolado, 1988), y para comunidades de peces en la misma Bahía de Mochima (Méndez et al. 1988). Se infiere que las condiciones ideales para una mayor diversidad de especies están en la estación 1, la cual, presenta factores físico-químicos más estables.
La menor diversidad correspondió a Varadero (3.00 bits/ind), siendo aún considerablemente alto. Margalef (1980) señala que la diversidad de especies es baja en comunidades transitorias, explotadas o bajo condiciones fluctuantes. Este último aspecto es característico de la estación de Varadero donde se presume que hay alta descarga de materia orgánica y acumulación de sedimento, por ser la estación más próxima a la zona de descarga de material terrígeno del estudio. Este mismo fenómeno se ha reportado en otras comunidades de T. testudinum en Venezuela; Jiménez (1994) lo señala para comunidades bentónicas de la Bahía de Mochima; Graterol (1986) en praderas de Thalassia de Tocuchare (Golfo de Cariaco) y Bitter (1988) en el Parque de Morrocoy.
Apesar del desarrollo de la Thalassia en la estación 3, su diversidad fue la segunda más baja. Según varios autores las especies asociadas a la Thalassia se incrementan con el aumento de la biomasa (Heck y Welstone, 1977, Heck y Orth, 1980), aunque persiste cierta duda con respecto a la biomasa como predictora de la abundancia de la epifauna (Virstern y Howard, 1987) la diversidad en Toporo parece ser un ejemplo de esta segunda situación.
El comportamiento de la equitatividad en todas las estaciones fue muy similar al de la diversidad, siendo probablemente, determinada por las mismas razones que regulan la diversidad. Los valores altos de equidad en la estación 1, indican que el número de especies encontradas en esta área es mayor y están mejor distribuidas. Esto también se ha informado en otras comunidades de moluscos asociadas a T. testudinum en la zona nororiental de Venezuela (Graterol 1986). Al igual que en el caso de comunidades de Thalassia del Parque Nacional Morrocoy (Bitter, 1988), la equidad varió dependiendo de la localidad. Todo lo anterior supone la dominancia de una o un número reducido de especies. Margalef (1980) señala que las comunidades con dominancia de una pocas especies son siempre comunidades con poca diversidad, explicándose los valores relativamente bajos de la diversidad en la estación 4 y 3, donde dominan dos especies (C. litteratum y A. notabilis, C. litteratum y C. eburneum, respectivamente). En la estación 2, la diversidad aumenta un poco debido a que la dominancia se distribuye entre tres especies (A. notabilis, M. americanus y C. cancellata). En el caso la estación 1, la diversidad es la mayor y la dominancia está distribuida entre siete especies, siendo A. notabilis, C. litteratum y T. variegata las más abundantes. La dominancia influye sobre la diversidad y en la distribución no equitativa de las especies en el medio natural. Jackson (1972), señala una relación directa que existe entre la depredación y la diversidad de moluscos en las comunidades de Thalassia en Jamaica, demostrando que el aumento de la depredación tiende a disminuir la dominancia y aumenta la diversidad y la equidad. Graterol (1986) relacionó los valores elevados de diversidad de gasterópodos con aumento en la diversidad de bivalvos, debido a la capacidad de muchos gasterópodos de ser depredadores auténticos de criaderos de bivalvos, lo cual disminuye la dominancia de estas especies. En este trabajo se encontraron gasterópodos predadores (murícidos) mayormente en las estaciones 1 y 3, lo cual, pudo contribuir a sus valores altos de diversidad. Los porcentajes de aparición corroboran la tendencia de la dominancia a ser mayor cuando la diversidad y la equidad son menores. En la estación 4, el porcentaje de aparición de A. notabilis y C. litteratum fue de 100%, lo contrario sucede en la estación 1, donde solo tres especies presentan un porcentaje máximo de aparición de 75%.
El análisis del porcentaje de aparición permitió ubicar las 81 especies de las cuatro estaciones dentro de tres categorías: 16 especies entre 50% y 75%, 24 especies entre 25% y 50%, y las 41 especie restantes con menos del 25%.
Los moluscos son considerados conjuntamente con los equinodermos y poliquetos como los macroinvertebrados más abundantes asociados a T. testudinum (Jackson 1972, Bitter 1999), donde la salinidad y la temperatura pueden ser factores importantes en la distribución de los bivalvos de las zonas someras del Caribe. La presencia en las estaciones de especies de las familias Veneridae y Semelidae son debidas a la escasa cantidad de arcilla que caracterizan los sedimentos de la bahía (Caraballo 1968). La composición de especies de moluscos asociados a T. testudinum en la Bahía de Mochima coincide con la informada para otras áreas del Golfo de Cariaco (Graterol 1985), pero defiere de las reportadas para otras localidades del Parque Nacional Morrocoy (Bitter 1999, Rodríguez y Villamizar 2000).
Resumen
Se estudio la diversidad y abundancia de la comunidad béntica malacológica asociada a Thalassia testudinum en cuatro localidades de la Bahía de Mochima, Estado Sucre, Venezuela. Las muestras fueron recolectadas mensualmente en cada localidad sobre transeptos perpendiculares a diferentes profundidades (0-4m) desde enero 1991 hasta diciembre 1991, utilizando una cuadrata de 0.25 m 2 para recolectar los organismos con el sedimento. Un total de 2 988 organismos de la infauna y epifauna, pertenecientes a las clases Gastrópoda (40) y Bivalvia (41) fueron identificados. Las especies más abundantes y en consecuencia mejor adaptadas a los habitats suministrados por la T. testudinum en la Bahía de Mochima fueron Anadara notabilis, Codakia orbicularis, Cerithium litteratum, Cerithium eburneum, Batillaria minima, Modiolus squamosus, Modulus modulus, Chione cancellata, Turritella variegata, Arca zebra, y Laevicardium laevigatum. Se determinaron diferencias significativas en el número de organismos en relación a la profundidad y el tiempo en la localidad de la Gabarra, la cual presentó los valores mas altos de diversidad total (4.51 bits/ind) y mensual (2.75-3.90 bits/ind). La biomasa y la abundancia de moluscos fueron bajas y la estación mas interna (Varadero) presentó los valores mas elevados. El bivalvo A. notabilis y el gastropodo Modulus modulus fueron las especies constantes en las cuatro estaciones.
]]> ReferenciasAbbott, T. 1974. American Seashells. Van Nostrand. New York. 663 p. [ Links ]
Alcolado, P. 1990. El bentos de la macrofauna del Golfo de Batabanó, Cuba. Editorial Academia del Instituto de Oceanología 161 p. [ Links ]
Almeida, P. 1974. Distribución de moluscos de las costas centro-occidentales (Patanemos-Punta Tucacas) de Venezuela. Comparación de hábitas litorales. Mem. Soc. Cienc. Nat. La Salle 34: 24-32. [ Links ]
Bello, G. 1989. Comunita di gasteropodi di una partería di Thalassia testudinum di St. Croix, Caraibi. Men. Biol. Mar. Oceanogr. 17: 15-26. [ Links ]
Bitter, R. 1988. Análisis multivariado de la comunidad asociada a Thalassia testudinum en el Parque Nacional Morrocoy, Venezuela. Tesis Doctoral, Facultad de Ciencias, Postgrado de Biología, Universidad Central de Venezuela. 144 p. [ Links ]
Bitter, R. 1993. Estructura y función del campo de Thalassia testudinum como ecosistema. Ecotrópicos 6: 30-42. [ Links ]
Bitter, R. 1999. Benthic communities associated to Thalassia testudinum at three localities of Morrocoy National Park, Venezuela. Rev. Biol. Trop. 47: 443-452. [ Links ]
Caraballo, L. 1968. Sedimentos recientes de la Bahía de Mochima. Bol. Inst. Oceanogr. Univ. Oriente 7: 45-64. [ Links ]
Flores, C. 1968. Algunos gasterópodos de la Isla Las Aves, Venezuela y su distribución. Bol. Inst. Oceanogr. Univ. Oriente 42-109. [ Links ]
García, A. 1978. Observación de algunos parámetros ambientales de la Bahía de Mochima durante el período de enero-junio de 1977. Tesis de Maestreia. Postgrado en Ciencias Marinas, Instituto Oceanográfico de Venezuela, Universidad de Oriente, Cumaná, Venezuela. 111 p. [ Links ]
Graterol, A. 1986. Diversidad de moluscos en dos localidades del Golfo de Cariaco (Edo. Sucre), Venezuela. Tesis de Licenciatura. Departamento de Biología, Universidad de Oriente, Cumaná, Venezuela. 132 p. [ Links ]
Hambrook, J., M. Layrise & R. Colmenares. 1979. Contribución al conocimiento de la comunidad de Thalassia testudinum en Punta Morón. In P.G. Penchazzadeth (ed). Ecología del ambiente marino-costero. Ed. Universidad Simón Bolivar. [ Links ]
Heck, K.L. & G.S. Wetstone. 1977. Habitat complexity and invertebrate species Richness and abundance in tropical seagrass meadows. J. Biogeogr. 4: 135-142 [ Links ]
Heck, K. L. & R. J. Orth. 1980. Seagrass habitats: The roles of habitat complexity competition and predation in structuring associated fish and motile macroinvertebrates assemblages, p. 449-464. In V.S. Kennedy (ed). Estuarine Perspective, New York. [ Links ]
Howards, R. 1987. Dial variation in the abundance of epifauna associated with seagrasses of the Indian River, Florida, USA. Exp. Mar. Biol. 69: 137-142. [ Links ]
]]>Humphrey, M. 1975. Seashells of the West Indies. Ed. Collins. Londres 351 p. [ Links ]
Jackson, J. 1972. The ecology of the mollusc of Thalassia communities, Jamaica, West Indies. II Moluscan population variability along an environmental stress gradient. Mar. Biol. 14: 304 337. [ Links ]
Jiménez, M. 1994. Comunidad de moluscos asociadas a Thalassia testudinum en la Ensenada de Reyes, bahía de Mochima, Edo. Sucre, Venezuela. Bol. Inst. Oceanog. Univ. Oriente. 33: 67-76. [ Links ]
Jiménez, M., I. Liñero, J. P. Blanco & J. Fermín. 2000. Macrofauna béntica asociada con Thalasssia testudinum en Bahía de Mochima, Sucre, Venezuela. Rev.Biol. Trop. 48 (Supl.1): 233-242. [ Links ]
Kato, K. 1961. Some aspects on biochemical characteristics of sea water and sediments in Mochima Bay, Venezuela.
Bol. Inst. Oceanogr. Univ. Oriente 1: 246-265. ]]>
Krebs, C. 1989. Ecological Methodology: The Experimental Analysis of Distribution and Abundance. Harper
& Row, Nueva York. 654 p.
1 Dpto. de Biología, Escuela de Ciencias
2 Dpto. Biología Pesquera, Instituto Oceanográfico de Venezuela, Universidad de Oriente, apdo. Postal 245, Cumaná 6101, Edo. Sucre, Venezuela; clodeiro@sucre.udo.edu.ve

 ]]>
]]>