 ]]>
]]>
Biología floral, sistema reproductivo y éxito reproductivo de Macroptilium fraternum (Fabaceae)
Patricia S. Hoc 1, Susana I. Drewes1 & María T. Amela García1
Abstract
Observations about the floral biology and the reproductive system of Macroptilium fraternum were made in two populations in Argentina, with different edaphic conditions, situated at the southern area of distribution of the species. The coexistence of two kind of flowers in the same plant was detected in both populations and in herbarium material from other localities: a) cleistogamous preanthesis flowers and b) pseudocleistogamous flowers. The cleistogamous preanthesis flowers possesed wings longer than 5 mm and were disposed in pubescent, erect, racemes exposed above the level of the foliage. The anthesis lasted approximately 5 hours on sunny days or 9 hours on rainy days, the right wing covered the left wing, the corolla acquired a bilabiate aspect, offering the left wing as landing platform; they produced a low quantity of nectar (0.18 ± 0.13 µl); they did not receive pollinator visits; approximately four hours after the beginning of the anthesis on sunny days the ovary started to grow; in the bud, the receptive stigma was covered with germinating pollen grains from the same floral unit. The pseudocleistogamous flowers possesed wings shorter than 5 mm that were disposed in brief, hirsute and prostrate racemes, non subterranean as in other Macroptilium species; the banner started to spread exposing partially the wings, the left wing limb surrounded the keel and never spread; the right wing started to spread and after 2 seconds refolded and began to wither, the ovary began to grow immediately; the flower did not offer a surface to allow visitors to settle; in the buds the stigma was receptive and found with pollen grains from the same floral unit emiting their tubes. The relative reproductive success was low (natural pollination = 8%; spontaneous self-pollination = 3%), probably due to the low pollen viability, the high percentage of seed abortion and the lack of pollinators visits. The production of both flower types is not conditioned by edaphic factors, as they were found in plants belonging to different environments. Study of the progeny of both floral types in both populations has demonstrated that each one produces a similar amount of seeds contributing to the next generation with plants of similar characteristics (slow development, low resistance to infections and high mortality). The absence of pollinators in these flowers (in part because they preferred to visit flowers of other species with more reward and in part because of the fragmentation of the habitats), would be producing inbreeding depression in the studied populations; if the germplasm variability is not preserved, these populations situated at the Southern end of the distribution range of the species could become extinguished.
Key words: Macroptilium, Fabaceae, floral biology, cleistogamy, Southern distribution.
]]> A pesar de los aportes realizados al conocimiento sobre la biología floral del género Macroptilium (Benth.) Urban 1928 (Brizuela et al. 1993, Etcheverry et al. 1994, 1998), hasta el presente se desconocen tanto la biología floral como el sistema reproductivo de Macroptilium fraternum (Piper) Juárez et Pérez (1987). Varios autores (Bacigalupo 1987, Drewes 1995, 1997) han detectado en especies de Macroptilium la coexistencia de flores casmógamas y cleistógamas, sin embargo, no se han realizado observaciones de campo con el fin de interpretar la función de cada clase.
Lord (1981), basándose en aportes previos documentados, realizó una clasificación de los distintos tipos de flores cleistógamas y señaló que en una misma planta pueden coexistir distintas clases.
Con el fin de determinar la función de cada tipo de flor y si la producción de flores cleistógamas se halla condicionada por las características del suelo, se estudió la biología floral y la producción de flores en 2 poblaciones de M. fraternum con distintas condiciones edáficas, en el extremo sur de distribución de esta especie. Además se analizaron exsiccata provenientes de herbarios cuyas siglas se indican siguiendo a Holmgren et al. (1990).
El presente estudio constituye la primer contribución al conocimiento de la biología floral, el sistema y el éxito reproductivo de M. fraternum, interpreta la función de los diferentes tipos de flores y señala el impacto de la fragmentación ambiental en las dos áreas estudiadas, así como la necesidad de implementar planes para la conservación de esta especie nativa con cualidades forrajeras (Hoc, obs. pers.).
Materiales y métodos
Material de herbario estudiado: BOLIVIA: "In Bolivia", 1891, Miguel Bang 1011 (G) (holotypus!). ARGENTINA: Prov. Jujuy: Dpto. El Carmen: Ruta 9, 2 km al N Ayo. Las Vaquerias, 21/3/1982, Bravo y Palacios 1011 (BAFC); Ruta 9 y Ayo. Huaico Hondo, 21/3/1982, Bravo y Palacios 1014 (BAFC). Prov. Salta: Dpto. Rosario de Lerma: Quebrada del Toro, a lo largo de la ruta 51 (24 o 54 S 65 o 38 W, 27/2/95, 24/4/95, Hoc 284, 351 (BAFC); Campo Quijano, 20/2/1988, Hoc 71, 72 y 73 (BAFC). Dpto. La Caldera: La Caldera, 3/3/1988, Hoc 79 y 80 (BAFC); 25/3/79, Palacios et al. 781(BAFC). Dpto. La Capital: San Lorenzo, 25/3/1979, Palacios et al. 773, 775 y 25/3/1982, 1038 (BAFC); 20/2/1988, Hoc 58, 59 y 62 (BAFC); Salta, 25/3/1979, Palacios et al. 767 (BAFC). Dpto. Cerrillos: Cerrillos Estación de Extensión Experimental, I.N.T.A., 11/3/1982, Schinini 22261(CTES). Prov. Tucumán. Dpto. Trancas: Gonzalo, 30/3/1982, Bravo y Palacios 1046 (BAFC). Prov. Córdoba. Dpto. Punilla: Cabalango, 31 o 23 S 64 o 34 W, 24/2/1998, Drewes 506 (BAFC).
Áreas de estudio: a) las plantas de M. fraternum crecían formando varios manchones, en un terreno pedregoso, con escasa cobertura vegetal (margen de un arroyo con caudal sólo durante el período estival), en la Quebrada del Toro, a lo largo de la ruta 51 (24 o 54 S 65 o 38 W, Dpto. Rosario de Lerma, Salta, Argentina); esta área corresponde a la provincia fitogeográfica de las Yungas (Cabrera 1971), que en esa latitud se encuentra fragmentada debido a la actividad humana (cría de ganado en las pasturas naturales sin reposición, cultivos e intenso tránsito a lo largo de la ruta hacia Chile). Uno de los manchones fue el centro de las observaciones y estaba compuesto por tres plantas de M. fraternum y tres plantas de Phaseolus vulgaris var. aborigineus Burkart (1952), la parcela medía aproximadamente 5 x 10 m y se encontraba al costado de la ruta. Las observaciones se llevaron a cabo entre el 27 de febrero y el 24 de abril de 1995, comprendiendo todo el período de floración de la especie en el área. b) Las plantas de M. fraternum crecían en Cabalango (31 o 23 S 64 o 34 W, Dpto. Punilla, Córdoba, Argentina) en los faldeos serranos, que se encuentran fragmentados debido a la actividad humana (cría de ganado en las pasturas naturales y actividad turística), en un pastizal donde predominaban gramíneas; el suelo estaba cubierto por una densa capa de humus. Se identificaron dos manchones de aproximadamente 2 x 1.5 m y de 1 x 1.5 m en los cuales M. fraternum era la especie dominante. Las observaciones se llevaron a cabo durante el mes de febrero de 1998.
Condiciones meteorológicas: se registraron la temperatura y humedad relativa con un termohigrómetro digital así como la duración e intensidad de las precipitaciones.
Fases florales: su secuencia se siguió desde el comienzo de la antesis utilizando lupas de mano y un estéreomicroscopio; tanto en los capullos próximos a la antesis como en cada fase, se registró la existencia de receptividad estigmática siguiendo el método de Osborn et al. (1988), tomando como resultado positivo el burbujeo del peróxido de hidrógeno en el estigma. Se registró el momento en que se producía la dehiscencia de las anteras. Se fijaron en FAA (formol-acido acético-alcohol) flores correspondientes a cada estadío de la antesis desde el capullo hasta el crecimiento ovárico (si hubo fecundación). Cada planta se observó continuamente con el objeto de registrar el espectro y el comportamiento de los visitantes florales y extraflorales.
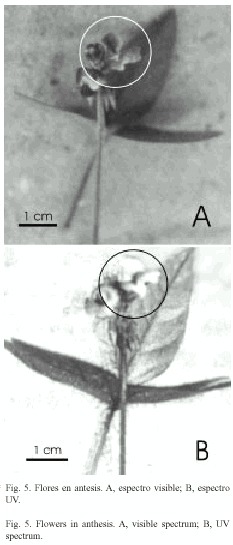
]]> Atractivos florales: para designar el color de las piezas florales se siguió a Kelly (1965); para determinar la existencia de guías de néctar en el espectro UV (en el cual los Himenópteros tienen visión), las flores en distintas fases de la antesis fueron fotografiadas con un filtro Kodak 18 A. La recolección del néctar floral se realizó al final de la antesis en un grupo de flores embolsadas antes de la apertura, con tubos capilares de diámetro conocido; con la medida de la altura alcanzada se calculó el volumen, utilizando la fórmula del cilindro. La concentración de hexosas se midió con tiras Diastix.Sistema reproductivo: en la población de la Quebrada del Toro, se embolsaron capullos para determinar la existencia de autopolinización espontánea, y se marcaron otros como testigos de polinización libre. Los frutos maduros de ambos tratamientos se colectaron el último día de observaciones. Se calculó el índice de autocompatibilidad ISI (Nº de frutos producidos por autopolinización/ Nº de frutos producidos por polinización cruzada) (Ruiz Zapata y Kalin Arroyo 1978) con una modificación equivalente, ya que se emplearon los frutos obtenidos de la autopolinización espontánea y de la polinización libre.
Se calcuó el éxito reproductivo relativo RRS (Nº de frutos/ Nº de flores) X (Nº de semillas / Nº de óvulos) x 100 (Dafni 1992) de cada tratamiento.
La cantidad de óvulos en esta especie es fácil de contar en los frutos maduros por su disposición uniseriada, y porque, de no producirse semillas, persisten sus restos en la placenta.
Observaciones de laboratorio: en ejemplares de distintas procedencias se midieron: inflorescencias (longitud total, longitud del pedúnculo, longitud del raquis, número de nudos, distancia entre nudos, pubescencia); flores (longitud de las alas); legumbres (número por inflorescencia, longitud, ancho, forma y número de semillas); semillas (número y ubicación por legumbre, forma, longitud, ancho, espesor y color).
Germinación del polen: la existencia de polen germinando en el estigma se determinó en capullos y flores (fijados en FAA en distintos estadios de desarrollo), montados sobre un portaobjetos con agua glicerinada (50%), posteriormente coloreados con una solución acuosa de violeta de cresilo (saturada) y observados con un microscopio de epifluorescencia provisto de un filtro V (380-425 nm).
Viabilidad polínica: se analizó siguiendo el método de Greissl (1989).
Aptitud: además del éxito reproductivo relativo se utilizaron los siguientes componentes, recomendados por Schemske y Paulter (1984) en Dafni (1992):
a) Germinación de las semillas: se seleccionaron semillas de ejemplares procedentes de las dos poblaciones y de los distintos tratamientos reproductivos, las cuales fueron reunidas en lotes según su procedencia y tratamientos reproductivos. Las semillas se desinfectaron sumergiéndolas en una solución al 5% de hipoclorito de sodio durante 5 minutos y a continuación se realizaron cinco lavados sucesivos con agua destilada. Todos los lotes, excepto uno (formado por una semilla de cada lote) se escarificaron realizando un corte en el tegumento seminal con una lanceta estéril. El material se hidrató durante 6 horas, luego se colocó cada lote en cámaras de germinación esterilizadas, con base de algodón cubierto por papel de filtro, previamente humedecido. Las cámaras de germinación se colocaron a temperatura ambiente (15-22°C) con intervalos de luz-oscuridad (14-10 horas).
b) Desarrollo de las plántulas: permanecieron en las cámaras de germinación durante 5 días; para su descripción se siguió el criterio de Baudet (1974).
]]> c) Formas de crecimiento de las plantas: las plántulas se transplantaron a macetas de 500 ml, que contenían 1/3 de arena en la base y 2/3 de tierra abonada, y se trasladaron a un invernáculo donde se siguió su desarrollo durante tres meses. Para su descripción se siguió el criterio de Tourn et al. (1999).Resultados
Se reconocieron dos tipos de flores durante el trabajo de campo (posteriormente corroborada analizando otros ejemplares herborizados) presentadas en:
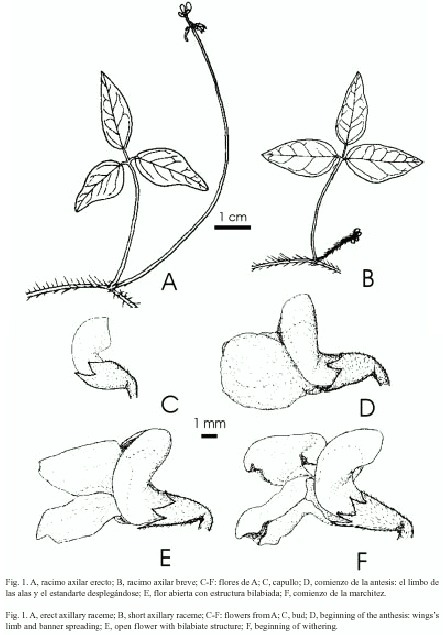
a) Inflorescencias largas (IL): racimos axilares largos (9-16 cm), erectos, portando hasta 11 flores generalmente geminadas, con alas mayores de 5 mm y expuestas por encima de las plantas circundantes (Fig. 1 A; Fig. 2 A).
b) Inflorescencias cortas (IC): racimos axilares breves (1.3-7 cm), paucifloros, de pedúnculo hirsuto y postrado, con flores geminadas y alas menores de 5 mm, parcialmente cubiertos por los cantos rodados del sustrato (en Salta) o por las plantas circundantes (en Córdoba), (Fig. 1 B; Fig. 2 A, recuadro).
Antesis y características de las flores
a) Flores con alas mayores de 5 mm en IL: Se abrían de una a tres por día en cada inflorescencia. Las fases florales desde el comienzo hasta la finalización de la antesis se representan en la Fig. 1 C-F y en la Fig. 2 B y C. Las alas comenzaban a desplegarse entre las 7:15-8:35 hr; el ala derecha cubría al ala izquierda, adquiriendo la corola aspecto bilabiado, ofreciendo el ala izquierda como plataforma de aterrizaje; el estigma, receptivo, estaba cubierto de polen proveniente de la misma flor presentado en los tricomas estilares y germinando (Fig. 3 A-C). Al cabo de 10 minutos en días sin lluvia, las alas estaban completamente desplegadas con el estigma aún receptivo. En el espectro visible las piezas florales exhibían los siguientes colores: alas, naranja rojizo moderado con las venas de color rojo oscuro; limbo del estandarte, marrón rojizo claro a oscuro; base del estandarte, uña y base de la quilla, amarillo verdoso pálido; ápice de la quilla, amarillo verdoso claro. El volumen de néctar producido era 0.18 ± 0.13 µl y contenía más de 2 000 mg de hexosas/dl. Después de cuatro horas del comienzo de la antesis las flores comenzaban a marchitarse y el ovario comenzaba a crecer. Al cabo de, aproximadamente, cinco horas en los días soleados y nueve horas en los días nublados o lluviosos, todas las flores que se habían abierto tenían las alas marchitas y en los estigmas persistía el polen propio germinando (Fig. 4). En las flores aún no marchitas, al ejercer una presión descendente sobre el ala izquierda, se exponía el extremo del estilo presentando el estigma y los tricomas circundantes, cubiertos de polen propio (que era expulsado como "pasta dental"); inmediatamente, las alas y el estandarte comenzaban a marchitarse. No se detectaron guías de néctar en el espectro UV (Fig. 5). Las flores no recibieron ningún visitante durante el período de observaciones.
b) Flores con alas menores de 5 mm en IC: el estandarte comenzaba a desplegarse exponiendo las alas parcialmente, el limbo del ala izquierda rodeaba la quilla y nunca se desplegaba. El limbo del ala derecha comenzaba a desplegarse pero rápidamente se marchitaba (t= 2 s), al cabo de pocos minutos el ovario comenzaba a crecer. En ningún momento las alas ofrecían una superficie para permitir que algún visitante se posara. No fue posible la recolección del néctar debido al tamaño diminuto de las flores y a su breve antesis.
]]> Nectarios extraflorales: En cada nudo de la inflorescencia se observaron glándulas extraflorales que secretaban néctar continuamente, entre las 8:30 y las 11:30 hs, período durante el cual eran visitadas constantemente por hormigas, sólo durante los días soleados.Viabilidad polínica: fue baja (polen viable 6%; polen no viable 8%; polen subviable 86%) (n=133).
Sistema reproductivo: el valor obtenido del ISI (0.64) indica que la especie es parcialmente autocompatible. Los resultados de los distintos tratamientos reproductivos aplicados a flores en IL se muestran en el Cuadro 1. La mayoría de las legumbres de ambos tratamientos no contenía semillas; en las que tenían semillas, generalmente no se habían fecundado los óvulos más próximos al estilo (o si se habían fecundado, las semillas abortaban), en ambos tratamientos.

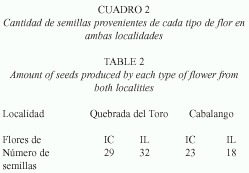
Producción de flores: La cantidad de semillas producidas por los 2 tipos de flores y en ambas localidades fue semejante (Cuadro 2).
]]>
Germinación: 48 h después de la siembra en las cámaras de germinación, se produjo la emergencia de la radícula. A las 96 h emergieron las plántulas completas. El porcentaje de germinación fue 90%, excepto el lote sin escarificar, que demoró más de diez días en germinar, obteniéndose un porcentaje de germinación del 50%.
Descripción de las plántulas: Presentan germinación hipógea con epicótile piloso; las hojas primordiales son opuestas, unifolioladas y de base cordada, con estípulas enteras y pecíolo con tres articulaciones; las primeras hojas son trifolioladas, opuestas y con estípulas bífidas.
Descripción de las formas de crecimiento: En los primeros estadios de desarrollo las plántulas son erectas, las hojas primordiales son caducas y el tallo y los pecíolos son hirsutos.
Al aumentar la longitud del eje principal, adquieren hábito postrado. El ritmo de crecimiento es lento, ya que a los 52 días las plantas miden entre 8-10.5 cm de longitud, poseen entre 7-9 nudos y las hojas miden entre 2.5-3.1 cm de longitud. Después de tres meses de desarrollo, la longitud del eje principal es de 15-18 cm, con 11-15 nudos, que portan hojas de 6.3-8 cm de longitud. No se observaron ramificaciones y las yemas axilares presentes se mantuvieron retenidas durante el seguimiento. La pérdida de las hojas basales fue común. Las plantas adultas poseen un eje plagiótropo con filotaxis dística alterna, las hojas se ubican en un mismo plano por reorientación de los pecíolos y las láminas de los folíolos. En el campo se observó que las estructuras reproductivas aparecen en posición lateral (axila de las hojas) y por lo tanto no afectan el desarrollo del eje principal (monopodio).
Desarrrollo de las plántulas: fue muy lento, tuvieron baja resistencia a infecciones y no prosperaron después del tercer mes de emergidas.
Discusión
Según la clasificación de Lord (1981), las flores de alas mayores de 5 mm presentadas en IL son cleistógamas preantesis, ya que en ellas la autopolinización es previa al inicio de la antesis; las flores de alas menores de 5 mm presentadas en IC, que permanecen cerradas, son pseudocleistógamas, ya que no se observaron diferencias morfológicas entre ambos tipos, excepto la diferencia de tamaño y que carecen de antesis. En ambos tipos, la polinización ocurre en el capullo. Lord (1981) sugirió la coexistencia de flores chasmógamas con flores cleistógamas; este estudio confirma la coexistencia de dos tipos de flores presentadas en inflorescencias con distintas características en el mismo individuo.
]]> Darwin (1877) y Maheshwari (1962) señalaron que la producción de las flores cleistógamas estaría condicionada por factores edáficos y ambientales. En M. fraternum estaría determinada genéticamente, sin ser influenciada por factores extrínsecos, ya que los dos tipos florales fueron registrados no sólo en las dos poblaciones donde se desarrolló el trabajo de campo, sino también en la totalidad del material de herbario examinado, el cual pertenece a una amplia gama de ambientes.Considerando la antesis diurna, la superficie donde podrían posarse insectos, la necesidad de abrir la flor para acceder a la recompensa, el néctar escaso y oculto no muy profundamente y los órganos reproductivos no expuestos, el síndrome de las flores que se abren (cleistógamas preantesis) correspondería al de melitofilia descripto por Faegri y Van der Pijl (1979); aunque potencialmente pueden ser visitadas, no fueron visitadas durante el período de observación; no obstante, produjeron frutos. La antesis se alargaba con las lluvias o la nubosidad y la temperatura más baja. Probablemente este mecanismo constituya una estrategia para obtener fecundación cruzada prolongando la duración de la antesis cuando las condiciones climáticas no son favorables.
Los granos de polen presentados en los tricomas estilares y el estigma antes del comienzo de la antesis pueden germinar, por lo tanto, es posible la autogamia, concordando con los resultados obtenidos de los tratamientos reproductivos realizados a campo. En las flores embolsadas se registró un mayor porcentaje de crecimiento ovárico una vez finalizada la antesis; es posible que al haber estado protegidas hayan tenido mayor probabilidad de desarrollo inicial con respecto de las no embolsadas, sin embargo el valor del RRS fue menor que el de las flores expuestas a la polinización natural; probablemente, el mayor contenido de humedad dentro de las bolsas, debido a las lloviznas y lluvia persistente, afectó el desarrollo de los frutos.
La fecundación de los óvulos basales y el aborto de semillas apicales en ambos tratamientos indicaría que el desarrollo de ellos es favorecido por su cercanía a los recursos de la planta madre.
El bajo RRS es consecuencia de la baja viabilidad polínica y la escasa producción de semillas.
El aporte a la progenie por ambos tipos de flores parece equitativo.
La presencia de nectarios extraflorales ya había sido señalada por Drewes (1991, 1995) para M. fraternum; en el presente aporte se detectó la producción de néctar, el período de actividad y la presencia de hormigas en los mismos, comprobándose que, durante los días en que se producían precipitaciones, aunque el néctar extrafloral era secretado, las hormigas no visitaban las inflorescencias.
El desarrollo de las plantas corresponde al modelo arquitectural de Troll (Tourn et al.1999).
Si bien las flores fructificaban a pesar de la ausencia de polinizadores, la cantidad de frutos y semillas en ambas poblaciones fue escasa, no se observaron plántulas en el campo y, al menos las emergidas en el laboratorio, presentaban baja viabilidad; estos factores podrían estar reflejando una depresión por autogamia en las poblaciones estudiadas, causada por la falta de polinizadores, debido a la fragmentación y/o antropización del ambiente y a la competencia por polinizadores, dado que otras especies en flor eran frecuentemente visitadas (Hoc y Amela 1997, 1999) por insectos que podrían polinizar a M. fraternum. Teniendo en cuenta lo precedente, las poblaciones estudiadas de esta especie podrían desaparecer.
<>Es probable que en un ambiente no tan fragmentado, las flores cleistógamas preantesis reciban polinizadores y se produzca xenogamia, aumentando la variabilidad génica de la especie. Se espera poder corroborar esta inferencia en un área expuesta a menor antropización en un futuro próximo.Considerando las cualidades forrajeras de M. fraternum es importante la preservación de su germoplasma para cultivarla para la cría de ganado, para planes de repoblación de pasturas naturales (en general sobrepastoreadas).
]]> ResumenSe realizaron observaciones de la biología floral y el sistema reproductivo de Macroptilium fraternum en dos poblaciones de la Argentina, con diferentes condiciones edáficas, localizadas en el extremo Sur del área de distribución de esta especie. En ambas poblaciones y en material de herbario de distintas procedencias se determinó la coexistencia en una misma planta de dos tipos florales: a) flores cleistógamas preantesis y b) flores pseudocleistógamas. Las flores cleistógamas preantesis con alas mayores de 5 mm, dispuestas en racimos pubescentes, erectos, expuestos sobre el nivel del follaje. La antesis duraba aproximadamente 5 horas en los días soleados y 9 horas en los días lluviosos, el ala derecha cubría al ala izquierda, adquiriendo la corola aspecto bilabiado, ofreciendo el ala izquierda como plataforma de aterrizaje; producían escasa cantidad de néctar (0.18 ± 0.13 µl) y no recibieron visitas de polinizadores; aproximadamente cuatro horas después del inicio de la antesis en días soleados el ovario comenzaba a crecer; en el capullo, el estigma receptivo se encontraba cubierto con granos de polen de la misma unidad floral germinando. Las flores pseudocleistógamas con alas menores de 5 mm, dispuestas en racimos breves, hirsutos y postrados, no subterráneos como en otras especies de Macroptilium. El estandarte comenzaba a desplegarse exponiendo parcialmente las alas, el limbo del ala izquierda rodeaba la quilla y nunca se desplegaba; el ala derecha comenzaba a desplegarse y a los 2 segundos se replegaba y marchitaba, inmediatamente el ovario comenzaba a crecer; la flor no ofrecía ninguna superficie donde algún visitante pudiera posarse; en los capullos el estigma estaba receptivo y con los granos de polen de la misma unidad floral emitiendo sus tubos polínicos. El éxito reproductivo relativo fue bajo (polinización natural = 8%, autopolinización espontánea = 3%), debido probablemente a la baja viabilidad polínica, el alto porcentaje de aborto de semillas y la falta de visitas de polinizadores. La producción de ambos tipos de flores no está condicionada por factores edáficos, ya que se encontraron en plantas provenientes de distintos ambientes. El estudio de la progenie de ambos tipos florales provenientes de ambas poblaciones, ha demostrado que cada uno produce una cantidad semejante de semillas aportando a la próxima generación plantas con similares características (desarrollo lento, baja resistencia a enfermedades y alta mortalidad). La ausencia de polinizadores en estas flores (en parte porque preferían visitar flores de otras especies, con mayor recompensa y en parte por la fragmentación de los habitats), estaría produciendo una depresión por autogamia en las poblaciones estudiadas; si no es preservada la variabilidad del germoplasma, estas poblaciones situadas en el extremo Sur de distribución de la especie podrían extinguirse.
Agradecimientos
Agradecemos a A. Martínez por su ayuda durante las observaciones de campo, a R. Palacios por la lectura del manuscrito, a Janice Saunders por la revisión del "abstract", a la Universidad de Buenos Aires y al CONICET por los subsidios EX 306 y 0204/97, respectivamente, con parte de los cuales fue financiada la presente investigación.
Referencias
Bacigalupo, N.M. 1987. Macroptilium (Benth. ) Urban, pp. 732-738. In A. Burkart, N.S. Troncoso de Burkart & N.M. Bacigalupo (eds.) Flora Ilustrada de Entre Ríos III. Colección Científica del I.N.T.A., Buenos Aires. [ Links ]
Baudet, J.C. 1974. Signification taxonomique des caracteres blastogéniques dans la tribu de Papilionaceae Phaseoleae. Bull. Jard. Bot. Nat. Belg. 44: 259-293. [ Links ]
Brizuela, M.M., P.S. Hoc, V.S. Distilio, M.A. Agulló & R.A. Palacios. 1993. Biología floral de Macropilium bracteatum (Leguminosae, Phaseoleae). Darwiniana 32 (1-4): 41-57. [ Links ]
Burkart, A. 1952. Las leguminosas argentinas silvestres y cultivadas. Ed. Acme, Buenos Aires. 569 p. [ Links ]
Cabrera, A. 1971. Las regiones fitogeográficas de la Argentina. Ed. Acme, Buenos Aires. [ Links ]
Dafni, A. 1992. Pollination Ecology. A practical approach. Oxford University, Oxford. 250 p. [ Links ]
Darwin, C. 1877. The different forms of flowers on plants of the same species. Appleton, New York. [ Links ]
Drewes, S.I. 1991. Typology of the synflorescence, development of extra floral nectaries and growth forms in Macroptilium (Benth.) Urban (Leguminosae- Phaseoleae). Beitr. Biol. Pflanzen 66: 407-420. [ Links ]
Drewes, S.I. 1995. Revisión de las especies argentinas del género Macroptilium (Leguminosae-Phaseolineae). Tesis doctoral, Universidad de Buenos Aires, Buenos Aires, Argentina. [ Links ]
Drewes, S.I. 1997. El género Macroptilium (Fabaceae) en la flora argentina. Bol. Soc. Argent. Bot. 32 (3-4): 195-216. [ Links ]
Etcheverry, A., S.M. Pérez de Bianchi & D. Martín de López. 1994. Fenología floral de M. erythroloma (Benth.) Urban (Fabaceae) y características reproductivas. ST N 16. Resúmenes: 822. VI Congreso la-tinoamericano de botánica. Mar del Plata. Argentina 2-8/X/7/94. [ Links ]
Etcheverry, A., S.M. Pérez de Bianchi & D. Martín de López. 1998. Fenología floral de Macroptilium lathyroides (L.) Urban (Fabaceae) y características reproductivas. Aportes Bot. Salta, Ser. Misc. 1 (7): 14 p. [ Links ]
Faegri, K. & L. Van der Pijl. 1979. The Principles of Pollination Ecology. Pergamon, Oxford. 244 p. [ Links ]
Greissl, R. 1989. Vitality analysis of monadic and polyadic pollen grains using optical contrast fluorescence microscopy. Zeiss, Sci. and Techn. Inform. 15: 180-184. [ Links ]
Hoc, P.S. & M.T. Amela García. 1997. Floral biology and reproductive system of Phaseolus augusti (Fabaceae). Beitr. Biol. Pflanzen 70: 121-140. [ Links ]
Hoc, P.S. & M.T. Amela García. 1999. Biología floral y sistema reproductivo de Phaseolus vulgaris var. aborigineus (Fabaceae). Rev. Biol. Trop. 47: 59-67. [ Links ]
Holmgren, P.K., N.H. Holmgren & C. Batnett. 1990. Index Herbariorum. Part I: The Herbaria of the World. Ed. Reg. Veg. 120: 1-693. New York Botanical Garden. [ Links ]
Juárez, F. & S. Pérez. 1987. El género Macroptilium (Fa-baceae) en la provincia de Salta, Argentina. Anales INTA Salta 1: 31-42. [ Links ]
Kelly, K.L. 1965. ISCC-NBS Color-name Charts Illustrated with Centroid Colors. Standard Sample N![]() 2106. Suppl. to Nat. Bur. Standards Cir. 53. U.S. Government Printing Office, Washington, D.C.
2106. Suppl. to Nat. Bur. Standards Cir. 53. U.S. Government Printing Office, Washington, D.C.
Lord, E.M. 1981. Cleistogamy: a tool for the study of floral morphogenesis, funtion and evolution. Bot. Rev. 47: 421-449. [ Links ]
Maheshwari, J.K. 1962. Cleistogamy in angiosperms, pp. 145-155 In P. Maheshwari, B.M. Johri, I.K. Vasil (eds.). Proceedings of the summwer school of botany. June 2-15, 1960. Darjeeling. Ministry of Scientific Research and Cultural Affairs, New Delhi. [ Links ]
Osborn, M.M., P.G. Kevan & M.A. Lane. 1988. Pollinaton biology of Opuntia polyacantha and Opuntia phaeacantha (Cactaceae) in Southern Colorado. Pl. Syst. Evol. 159: 85-94. [ Links ]
Ruiz Zapata, T. & M. Kalin Arroyo. 1978. Plant reproductive ecology of a secondary deciduous tropical forest in Venezuela. Biotropica 10: 221-230. [ Links ]
]]>Tourn, G.M., D. Brathelemy & J. Grosfeld. 1999. Una aproximación a la arquitectura vegetal: conceptos, objetivos y metodología. Bol. Soc. Argent. Bot. 34 (1-2): 85-100. [ Links ]
Urban, I. 1928. Plantae cubenses novae vel rariores. L. Elkman lectae. IV. Symbol. Antill. 9: 433-543. [ Links ]
1 Lab. Sist. Plantas Vasculares, Pab. 2, 4º piso, Ciudad Universitaria. 1428EHA Buenos Aires. Argentina. Tel./Fax: 54-11-45763384; hoc@bg.fcen.uba.ar , drewes@bg.fcen.uba.ar