
Abstract
The development of the prothallia in nine species of Callipteris Bory (Woodsiaceae), four species of Diplazium, the genus from which Callipteris has been segregated, and one species of Dryoathyrium are studied in this paper. The data obtained are compared with the development of the prothallia of several other genera of Woodsiaceae. The main difference between Callipteris and the other genera of Woodsiaceae was that the antheridia dehiscence of Callipteris, occurs by an opercular cell and whereas some of Diplazium and other Woodsiaceae antheridia dehiscence occurs by a lateral or apical pore. The presence of antheridiogens is suspected on the basis of small ameristic male gametophytes in the cultures. Spore germination in Callipteris is Vittaria-type, prothallial development corresponds to Adiantum -type.
Key words: Callipteris, Diplazium, ferns, gametophyte, germination, prothallus, Woodsiaceae.
Callipteris es un género pantropical y fue segregado de Diplazium por Bory (1804) tomando como base las venas anastomosadas. Recientemente Pacheco y Moran (1999) redefinieron el género con base en las escamas del rizoma con márgenes castaño oscuros a negros y dientes marginales bífidos, caracteres que se encuentran en la especie típica, C. prolifera (Lam.) Bory.
Se revisó la literatura para conocer la morfología de los gametofitos de Callipteris y otros géneros de la familia Woodsiaceae. La literatura más importante encontrada fue: Momose (1938), Mehra (1949), Blasdell (1963), Atkinson (1967), Profumo (1969), Bir (1969, 1970), Nayar y Bajpai (1970) Nayar y Kaur (1971), Kaur y Chandra (1973), Mehra y Gupta (1986), Kato y Kramer (1990), Lin et al. (1992), Gastony y Ungerer (1997), Bao et al. (1998), Takamiya et al. (1999). ]]>
El propósito de este trabajo fue estudiar el desarrollo y morfología de los gametofitos de nueve especies de Callipteri s: Callipteris aberrans (Maxon y C. V. Morton) Pacheco y R.C. Moran, C. ceratolepis (H. Christ) Pacheco y R.C. Moran, C. chimborazensis (Baker) Pacheco y R.C. Moran, C. matamensis (A. Rojas) Pacheco y R.C. Moran, C. pactilis (Lellinger) Pacheco y R.C. Moran, C. pinnatifida (Kunze) Fée, C. prolifera (Lam.) Bory, C. rivalis (Baker) Pacheco y R.C. Moran, C. sanderi (C. Chr.) Pacheco y R.C. Moran, así como compararlos con otros géneros de la familia Woodsiaceae tales como: Diplazium hians Kunze ex Klotzsch, D. striatum (L.) C. Presl, D. ternatum Liebm., D. werckleanum H. Christ, Dryoathyrium sp., y la información obtenida de la literatura publicada.Materiales y métodos
Las esporas usadas fueron recolectadas en México, Costa Rica, Ecuador y el Jardín Botánico de Nueva York. Los ejemplares de respaldo están depositados en el Herbario Metropolitano "Dr. Ramón Riba y Nava Esparza" (UA-MIZ), (véase Cuadro 1). Las esporas se obtuvieron de hojas fértiles, desecadas dentro de sobres de papel. Para eliminar residuos de esporangios el contenido de éstos se tamizó a través de una malla de 0.074 mm de diámetro.
Sin embargo algunos esporangios lograron pasar a través de la malla. Las esporas de cada especie, sin recibir ningún tratamiento de esterilización, se sembraron en cuatro cajas de Petri de 5 cm de diámetro con cuatro repeticiones en agar, con medio de cultivo de Thompson (Klekowski 1969), una de las cuales se cubrió con papel estaño para mantenerla en la oscuridad y determinar fotoblastismo. La siembra se hizo con un pincel delgado. La densidad promedio de las esporas sembradas es de 230 esporas por cm 2 . Los cultivos se incubaron en condiciones de laboratorio con luz artificial (Osram 75 watts, luz de día), con un fotoperíodo de 12 hr luz-oscuridad y temperatura de 18-30 ºC.
Los cultivos se revisaron semanalmente para detectar el inicio de la germinación, humedeciéndose el medio con agua destilada estéril en cada revisión. La caja mantenida en la oscuridad se abrió hasta los 100 días aproximadamente, después de la siembra. En las cajas con cultivos donde se sospechaba la presencia de anteridiógenos se resembraron esporas nuevamente, con cuatro repeticiones.
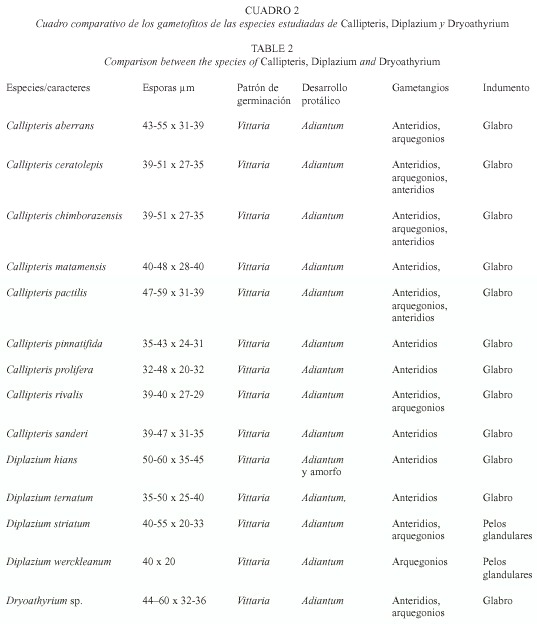
Esporas. Son monoletes, elipsoidales, la abertura con frecuencia está cubierta por amplias alas del perisporio, en las especies estudiadas estas alas son ligeramente erosas en el margen, en C. rivalis el perisporio entre las alas es irregularmente reticulado (Pacheco y Moran 1999). Las esporas más grandes de las especies estudiadas corresponden a D. hians con 50-60 µm y las más pequeñas a C. prolifera 32-48 µm. El perisporio entre las alas en Diplazium hians es equinado-perforado, por el contrario en D. ternatum presenta un patrón reticulado entre las alas.
Germinación. La germinación se inicia en un lapso de 4-24 días después de la siembra (Fig. 1), siendo las más tempranas en Dryoathyrium y las más tardías en C. ceratolepis y C. sanderi. Las esporas de C. chimborazensis (Pacheco 3320) y C. pinnatifida no germinaron en el primer caso, en la segunda especie el porcentaje de germinación fue mínimo ya que la mayoría de las esporas fueron abortadas. Las esporas mantenidas en la oscuridad no mostraron señales de germinación después de 100 días de sembradas, lo que indica que son fotoblásticas positivas. ]]>
En las especies de Callipteris estudiadas, el rizoide es la primera célula en emerger de la cubierta de la espora (Fig. 2), es hialino, pero bajo condiciones favorables de luz puede desarrollar pequeños cloroplastos, se encuentra en un plano perpendicular al filamento de las células protálicas. En D. hians y D. ternatum hay un crecimiento de la célula protálica y una rizoidal; en ambos casos las células protálicas se caracterizan por presentar gotas de grasa grandes de color amarillo brillante y numerosos cloroplastos.El patrón de germinación es tipo Vittaria ya que la célula protálica inicial se divide en ángulo recto a la primera pared y una de las dos células se divide para formar el filamento (Nayar y Kaur 1969). En un cultivo de D. ternatum sin tamizar, se observó germinación intraesporangial, ya que las esporas que se encontraban dentro de las cápsulas desarrollaron gametofitos similares a los desarrollados por esporas liberadas.
Fase filamentosa. En general se inicia entre los 8-25 días y se compone de 2-5 células (Fig. 3). La fase bidimensional se inicia en todas las especies de Callipteris entre los 16-26 días por divisiones anticlinales de la célula terminal del filamento. En Dryoathyrium se observaron gametofitos bidimensionales a los 13 días. D. werckleanum a los 20 días, en D. hians y D. ternatum se presentan gametofitos bidimensionales a los 30 días; observamos que a los 27 días de la siembra algunas de las esporas de D. hians presentan el tipo de patrón de desarrollo amorfo, que se caracteriza por no presentar una polaridad definida.
En todos los cultivos de Callipteris hay una asincronía más marcada en su germinación. En C. pactilis, por ejemplo, a partir de los 22 días se pueden observar distintas fases: dos células protálicas y una rizoidal (20%); fases filamentosas de 3-5 células protálicas con 2 células rizoidales (50%), filamentos de 6 células protálicas y 2 rizoidales, quedando una célula meristemática inicial cuneiforme, la cual desde esta edad va formando la lámina (20%). El 10% restante presenta fases bidimensionales de 15-26 células. A los 26 días se diferencia la célula meristemática en forma de una cuña central, los gametofitos desarrollan alas más o menos isodiamétricas, carecen de tricomas y conservan la cubierta de la espora a los 36 días de la siembra (Fig. 4).
Fase laminar. En general a los 57 días el gametofito se caracteriza por ser espatulado y por presentar amplias alas isodiamétricas, con o sin pliegues y la zona meristemática ampliamente desarrollada por lo que corresponde al desarrollo protálico tipo Adiantum (Nayar y Kaur 1969), (Figs. 5 y 6). Estos rasgos morfológicos son compartidos por las especies de Callipteris y Dryoathyrium difiriendo solo en el número de días en aparecer, los más tempranos se presentan en Dryoathyrium a los 20 días. En Diplazium ternatum y D. hians esta fase se presenta alrededor de los 80 días, mientras que en D. striatum se desarrollan entre los 18-20 días.
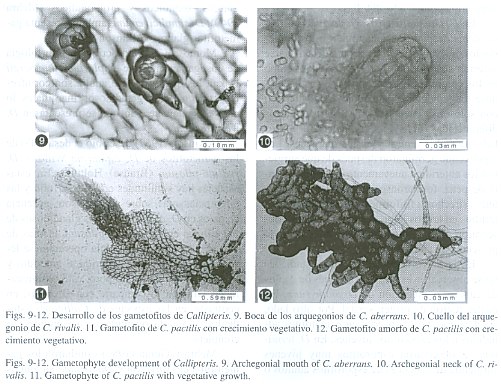
En C. chimborazensis, a los 29 días se presentan gametofitos bidimensionales con la célula meristemática apical más o menos central en forma de cuña, y con crecimientos vegetativos adicionales de 3-4 células. En C. aberrans a los 37 días se presenta un desarrollo similar (Fig. 7). A los 57 días los crecimientos vegetativos marginales alcanzan 10 células y se diferencia en ellos una célula meristemática en forma de cuña. Los crecimientos vegetativos adquieren forma espatulada similar al gametofito que les dio origen (Fig. 11). Algunos de los gametofitos de Callipteris en esta fase y debido a los crecimientos vegetativos tienen un crecimiento amorfo y conservan en la base la cubierta de la espora (Fig. 12). Este proceso se presenta en todas las especies.
Fase adulta. Los gametofitos adultos se presentan entre los 66 a 150 días en promedio, en Callipteris y Dryoathyrium son cordiformes con márgenes ondulados y glabros. En D. striatum y D. werckleanum los gametofitos son cordiformes con pelos glandulares en todo el gametofito. En D. striatum algunos gametofitos son espatulados con pelos glandulares, algunos gametofitos presentan crecimientos vegetativos apicales, los cuales también son espatulados. En D. ternatum los gametofitos adultos son glabros de forma espatulada con meristemo apical, las alas isodiamétricas y el talo continúa alargándose, con un cojinete denso y va adquiriendo la forma de banda.
Los anteridios tienen forma de barril, con tres células, la célula basal, la anular y una opercular (Fig. 8). En C. aberrans surgen los anteridios a los 66 días sobre el cojinete y márgenes del gametofito, en C. pactilis a los 76 días empiezan a surgir los anteridios sobre el cojinete y cercanos a la zona meristemática, en C. rivalis a los 67-70 días, y en C. ceratolepis a los 83 días, en C. matamensis a los 119 días y en C. chimborazensis a los 77 días. D. striatum tiene anteridios a los 132 días de la siembra. Los anteridios en D. ternatum y D. hians surgen a los 90-92 días de la siembra, la dehiscencia anteridial en esta última especie es por la formación de un poro que se abre en la célula opercular. No se observaron arquegonios ni esporofitos a los 230 días de siembra en estas dos especies.
El cuello de los arquegonios tiene cuatro hileras de células y cuatro células en la boca. En C. chimborazensis y Dryoathyrium presentan arquegonios a los 84 días, en C. pactilis y C. matamensis se encuentran a los 88 días, en C. ceratolepis a los 92 días, C. aberrans (Fig. 9) y C. rivalis a los 131 días y 155 días respectivamente (Fig. 10). En C. pinnatifida y C. sanderi no se observaron arquegonios después de 2 años de cultivo. En D. werckleanum presentaron arquegonios a los 62 días; D. striatum tiene arquegonios a los 84 días.
En C. chimborazensis, C. pactilis y C. ceratolepis después de que surgieron los arquegonios, nuevamente empezaron a desarrollarse anteridios tanto en los gametofitos viejos como en los crecimientos vegetativos, éstos haciéndose largamente espatulados, amerísticos o con sólo unas cuantas células meristemáticas diferenciadas (Fig. 11) y con anteridios en los lados dorsal y ventral y en los márgenes. Las esporas que germinaron después de que surgieron los anteridios nuevamente en la misma caja de petri, formaron gametofitos espatulados muy estrechos a filiformes, y desarrollaron anteridios en ambos lados y márgenes. No se observaron arquegonios. ]]>
Todo esto documenta la presencia de anteridiógenos. En la mayoría de las especies estudiadas no se observaron esporofitos después de 2 años de cultivo, a excepción de Dryoathyrium, los cuales fueron de origen sexual. Por otro lado, en el campo, no se observaron gametofitos adheridos a los esporofitos jóvenes. En D. ternatum se observaron esporofitos muy jóvenes surgiendo de las raíces de esporofitos adultos o bien por medio de las yemas, poblaciones con un gran número de individuos.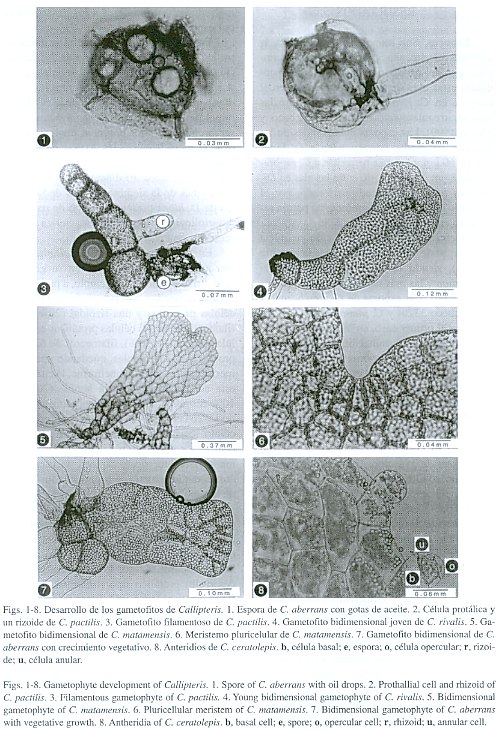
Takamiya et al. (1999) mencionan apomixis con gametofitos apogámicos para varias especies de Diplazium tales como D. hachi-joense Nakai, D. virescens Kunze var. virescen s; D. virescens Kunze var. conterminum (H. Christ) Sa Kurata, D. dilatatum Blume, D. taiwanense Tagawa; esta condición generalmente asociada con una triploidía.
Existen semejanzas en el desarrollo protálico de las especies de Callipteris estudiadas con D. latifolium T. Moore, D. esculentum (Retz.) Sw. y D. simplicivenium (Mett.) Holttum, en D. latifolium la fase adulta es cordiforme con márgenes ondulados y glabros (Momose, 1938), en D. esculentum lo más relevante es la presencia de anteridios en fases filamentosas, cuando adultos son gametofitos glabros, monoicos y proterándicos (Mehra 1949), D. simplicivenium también presenta gametofitos glabros (Atkinson 1967).
Momose (1938) describió la morfología de los gametofitos de Diplazium thunbergii Nakai, los que en la fase adulta son cordiformes y con pelos glandulosos marginales lo cual es semejante a lo que se presenta en D. werckleanum.
Atkinson (1967) describió el desarrollo de los gametofitos de D. expansum Willd. y D. stellatopilosum (Brause) Holttum. En estas especies hay similitudes en su desarrollo y las diferencias más sobresalientes son: ausencia de pelos en D. expansum, pelos glandulosos de 1-2 células en D. stellatopilosum además de que en esta especie la célula opercular se levanta pero permanece adherida al anteridio y la dehiscencia es por un poro, este último carácter también lo comparte con D. hians; en contraste, en las especies de Callipteris estudiadas, la célula opercular se desprende por completo. ]]>
Mehra y Gupta (1986) estudiaron los gametofitos de Athyrium y Diplazium, en ambos casos los gametofitos son glabros pero en Athyrium se presentan anteridios con dos células anulares o dos células operculares, lo cual no se encontró en las especies estudiadas de Diplazium y Callipteris. Entre las especies de Diplazium que estudiaron: D. esculentum, D. polypodioides (Mett.) Blume, D. stoliczkae Bedd., y D. asperum Blume presentaron el desarrollo de los gametofitos como los encontrados en las especies de Callipteris estudiadas; pero la dehiscencia anteridial en Diplazium y Athyrium puede ser por un poro lateral o apical.Para otros géneros de la familia Woodsiaceae, Blasdell (1963) y Profumo (1969) mencionan que el gametofito de Cystopteris tiene pelos papilados. Cornopteris tenuisecta (Blume) Tardieu, tiene gametofitos con pelos unicelulares, papilados, clorofílicos y no glandulares, marginales y en la superficie del prótalo (Bir 1970). Atkinson (1967) y Lin et al. (1992) describieron pelos glandulares clavados unicelulares en el margen y en ambas superficies del talo en los gametofitos de Deparia japonica (Thunb.) M. Kato (=Diplazium japonicum (Thunb.) Bedd.) Kaur y Chandra (1973), Bao et al. (1998) citan para Woodsia y Protowoodsia un desarrollo protálico de tipo Aspidium, las alas con pelos unicelulares, papilados y con una capa cerosa. Bir (1969) menciona que los gametofitos de Diplaziopsis javanica (Blume) C. Chr., tienen pelos marginales, no glandulares. Kato y Kramer (1990) citan que para Gymnocarpium los gametofitos son pubescentes en las alas. El desarrollo protálico de Hypodematium es de tipo Aspidium y presenta pelos papilados, glandulares, o unicelulares en todo el talo (Nayar y Bajpai 1970, Lin et al. 1992).
Debido a que en Diplazium existen alrededor de 400 especies de las cuales sólo se han estudiado un número reducido, las inferencias taxonómicas que se mencionan son tentativas.
Los gametofitos de Diplazium y Athyrium comparten algunos caracteres tales como la germinación de tipo Vittaria, se presentan dos patrones de desarrollo, de tipo Adiantum y amorfo. La mayoría de los géneros de la familia presentan gametofitos pubescentes, (véase Cuadro 2). Pryer et al. (1995) menciona que los pelos en los gametofitos se presentan en familias derivadas, por lo cual se podría inferir que el carácter primitivo lo presenta Callipteris, algunas especies de Diplazium y Athyrium. La pérdida o ausencia de los tricomas en los gametofitos de Callipteris es un carácter que con seguridad lo une a Diplazium, pero la pérdida de la célula opercular durante la liberación de los anterozoides, las escamas del rizoma en el esporofito y la venación reticulada, apoyan la separación de Callipteris del resto de los géneros de la familia. A nivel específico los caracteres aportados por el gametofito son muy similares entre sí, variando solamente en el tiempo de aparición de cada una de las fases protálicas.
El trabajo de campo fue apoyado por el Consejo Nacional de Ciencia y Tecnología, México, el Jardín Botánico de Misurí y el Instituto Nacional de Biodiversidad, Costa Rica. Enrique Forero, David Neill, Nelson Zamora y Luis Diego Gómez proporcionaron ayuda económica y logística. Blanca Pérez-García facilitó el uso del laboratorio para el cultivo de los gametofitos así como la revisión y comentarios hechos al presente estudio. Robbin C. Moran aportó valiosos comentarios literatura. Irma Reyes Jaramillo revisó el trabajo, aportando comentarios y sugerencias. Jorge Lodigiani por la digitalización de las imágenes. María Burbano, Daniel Rubio, Angel Alvarado, William A. Haber, Rafael Chacón, Cristina Formoso, Reynaldo Aguilar Fernández y Erick Bello acompañaron al primer autor durante sus viajes de recolección a Ecuador y Costa Rica.
Resumen
En este estudio se presenta el desarrollo de los gametofitos de nueve especies de Callipteris Bory (Woodsia-ceae), cuatro especies de Diplazium, el género del cual fue segregado Callipteris y una especie de Dryoathyrium. Los resultados obtenidos se comparan, con los de otros géneros de Woodsiaceae. La principal diferencia encontrada entre Callipteris y los otros géneros de Woodsiaceae fue que la dehiscencia anteridial de Callipteris se presenta por una célula opercular, mientras que la dehiscencia anteridial de Diplazium y otras Woodsiaceae es por un poro lateral o apical. Se sospecha la presencia de anteridiógenos, por la presencia de pequeños gametofitos amerísticos y an-teridiados en los cultivos. En Callipteris la germinación es del tipo Vittaria, y el desarrollo protálico es del tipo Adiantum, para otros géneros de la familia, el desarrollo protálico puede ser también de tipo amorfo o Aspidium. ]]>
ReferenciasAtkinson, L. R. 1967. The gametophyte of Diplazium. Phytomorphology 17: 99-109. [ Links ]
Bao, W-M, X.H. Lin, Q-X. Wang & Z-W. Aur. 1998. Study on the development of gametophytes of ferns from north-eastern China X. Woodsiaceae. Bull. Bot. Research. 18: 407-413.
Bir, S.S. 1969. Observations on the morphology and anatomy of Diplaziopsis javanica (Bl.) C. Chr. Pl. Sci. 1:109-118. [ Links ]
Bir, S.S. 1970. A note on the gametophytes of Cystopteris tenuisecta. Amer. Fern J. 60:98-101. [ Links ]
Blasdell, B.F. 1963. A monographic study of the fern genus Cystopteris. Mem. Torrey Bot. Club 21: 1-102. [ Links ]
Bory de Saint-Vincent, J.B.G.M. 1804. Voyage dans les quatre principales iles des mers dAfrique. Vol. 1 F. Buisson, Paris. [ Links ]
Gastony, G.J. & M.C. Ungerer. 1997. Molecular systematics and a revised taxonomy of the onocleoid ferns (Dryopteridaceae: Onocleeae). Amer. J. Bot. 84:840-849.
Kato, M. & K.U. Kramer. 1990. Dryopteridaceae subfamily Athyrioideae. In K. Kubitzki (ed.). The Familiies and Genera of Vascular Plants. Pteridophytes and Gymnosperms. Springer-Verlag. Nueva York.
Kaur, S. & S. Chandra. 1973. Morphology of the prothallus of Woodsia obtusa. Phytomorphology 23: 171-175. [ Links ]
Klekowski, E.J., Jr. 1969. Reproductive biology of the Pteridophyta. III. A study of the Blechnaceae. J. Linn. Soc. 62: 347-359. [ Links ]
Lin, S-J., K. Hir & K. Iwatsuki. 1992. Prothallia of the ferns from Yunnan, China I. J. Faculty of Science, University of Tokyo. Secc. III, 15: 173-198. [ Links ]
Mehra, P.N. 1949. Some observations on the prothallus of Diplazium esculentum (Retz) Sw. J. Ind. Bot. Soc. 28: 28-35. [ Links ]
Mehra, P.N. & A. Gupta. 1986. Athyriaceae. pp. 146-152 In Gametophytes of Himalayan ferns. Pramodh P. Kapur at Raj Bandhu Industrial Co., New Delhi. Momose, S. 1938. Studies on the gametophytes of ferns (IV): On the prothallium of Diplazium thunbergi Nakai and D. latifolia Moore. J. Jap. Bot. 14: 43-49.
Nayar, B.K. & S. Kaur. 1969. Types of prothallial development in homosporous ferns. Phytomorphology 19:179-188. [ Links ]
Nayar, B.K. & N. Bajpai. 1970. A reinvestigation of the morphology of Hypodematium crenatum. Amer. Fern J. 60: 107-118. [ Links ]
Pacheco, L. & R.C. Moran. 1999. Monograph of the neotropical species of Callipteris with anastomosing veins (Woodsiaceae). Brittonia 51: 343-388. [ Links ]
Profumo, P. 1969. The gametophytes of Cystopteris fragilis and Cystopteris dickieana. Webbia 23: 317-328. [ Links ]
Pryer, K.M., A.R. Smith & J.E. Skog. 1995. Phylogenetic relationship of extant ferns based on evidence from morphology and rbcL sequences. Am. Fern J. 85: 205-282. [ Links ]
Takamiya, M., C. Takaoka & N. Ohta. 1999. Citological and reproductive studies on Japanese Diplazium (Woodsiaceae; Pteridophyta): apomictic reproduction in Diplazium with evergreen bi-to tripinnate leaves. J. Plant Res. 112: 419-436. [ Links ]
1 Universidad Autónoma Metropolitana-Iztapalapa, División de Ciencias Biológicas y de la Salud, Depto. de Biología-Botánica Estructural y Sistemática Vegetal, Apartado Postal 55-535, 09340 México, D. F. Fax: (52-55) 5804-4688. pacheco@xanum.uam.mx ]]>