The seed of Stylosanthes hamata is characterized by it´s a high level of dormancy, which is imposed by seed-coat impermeability to water entry (harseededness or hard-seed condition). The level of hardseededness between upper and lower articulations of seven diploid and five tetraploid accessions was studied under tropical conditions. Germination tests on naked seeds (relative humidity: 55-65%, constant temperature: 27-30º C, photoperiod: 12/12 hr) showed that hard-seed percentage was, in most cases, over 85% independent of ploidy level and seed position within the fruit. Histochemical analysis indicated that hardseededness in this species was associated with a completely sealed microphile and hilum. This feature was also associated to: a) an epidermis consisting of a palisade-like layer of macrosclereids, b) a like-line at the outermost region of these cells and c) a high concentration of hydrophobic compounds located all across the seed-coat.
]]>
Dentro de Stylosanthes hamata se reconocen dos niveles de ploidía: las líneas diploides localizadas en un amplio ambito de altitud (100 - 1 600 msnm), con regímenes de precipitación entre 500 - 1 700 mm anuales; y las líneas tetraploides que se distribuyen en regiones más bajas (menos de 1 000 msnm) pero con un amplio rango de variación en cuanto a precipitación anual se refiere (500 2 300 mm). Sin embargo, al norte de Suramérica ambos niveles de ploidia se localizan fundamentalmente en zonas áridas y semiáridas comprendidas entre los ocho y doce grados de latitud norte (Edye 1988, Guenni 1992).
La semilla de S. hamata así como la de otras leguminosas se caracteriza por presentar en general un alto grado de latencia impuesta por la cubierta seminal. Este tipo de latencia obedece a una restricción en la absorción de agua asociada a la composición química y a las características morfo-anatómicas de la cubierta. Tal y como lo informa Werker (1980/81) en el caso particular de las leguminosas, varias partes de la epidermis han sido sugeridas como responsables de la impermeabilidad, entre ellas una cutícula cerosa, la línea lúcida, paredes celulares cutinizadas o suberizadas y la presencia de material semejante al corcho en el tapón estrofiolar.
No obstante, dentro de S. hamata se ha observado que el grado de dureza puede variar de acuerdo al nivel de ploidía y al origen climático del genotipo (Guenni 1992). Desde el punto de vista ecológico tal característica reviste mucha importancia puesto que permite la incorporación de la semilla al banco de semillas del suelo, desempeñando así un papel fundamental en cuanto a la perpetuidad de la especie en un hábitat determinado y al mantenimiento de los bancos naturales de germoplasma (Baskin & Baskin 1989).
Dada la importancia que la latencia representa dentro de la especie, la presente investigación tuvo como fin el determinar la relación existente entre la latencia asociada a la restricción de la entrada de humedad y las características morfo-anatómicas e histoquímicas que presenta la cubierta seminal en la semilla de S. hamata desarrollada bajo condiciones tropicales.
Materiales y métodos ]]>
Las plantas fueron cultivadas en condiciones de invernadero en Maracay, Venezuela (10o15 N, 67o39 W), utilizando un suelo franco-arenoso de mediana fertilidad. Los potes fueron regados cada 48 horas hasta drenar, mientras se aplicó fertilizante cada 15 días con una solución nutritiva comercial (Bayfolan, Bayer MR). A los 120 días de crecimiento, la semilla fue cosechada y almacenada bajo condiciones de laboratorio (+ 25 ºC), durante 60 días, hasta superar el período de latencia embrionaria propio de la especie (Gardener 1975). La semilla almacenada se separó de acuerdo a su posición en fruto y posteriormente se procedió a la realización de las siguientes pruebas:
a) Porcentaje de dureza: Para tal fin 50 semillas distales y basales de c/u de las 12 líneas desprovistas de la vaina se colocaron en placas de Petri que contenían 40 ml de agar al 1%. Dichas placas (4 por línea y posición dentro del fruto) fueron llevadas a una cámara de germinación durante diez días. Las semillas fueron expuestas diariamente a una humedad relativa entre 55 y 65%, y a una temperatura que osciló entre 27 y 30 ºC. La luz fue suministrada mediante dos bombillos de luz blanca fluorescente (110 W) con un fotoperíodo de doce horas. Se contaron diariamente las semillas germinadas y al final de la prueba las que no germinaron fueron seccionadas en su porción apical (Mott 1979) para facilitar la entrada de humedad. Al cabo de 48 horas se contaron las semillas duras germinadas, determinando así el total de semillas duras en función al total de semillas viables.
b) Determinación del sitio de entrada de humedad: Un total de 40 semillas distales y basales sin vaina de c/u de las 12 líneas se colocaron en solución de sulfato ferroso de amonio 0.003 M (Ballard 1973). El sitio de la cubierta seminal ennegrecido por el contacto con la solución indicó el lugar de entrada de humedad. Se realizaron observaciones cada 24 horas durante doce días, para seguir el patrón de imbibición y cuantificar el total de semillas ennegrecidas.
c) Determinación de las características morfo-anatómicas: Para la realización del estudio morfo-anatónico de la semilla se consideraron algunos de los caracteres señalados por Escala (1994) como de valor diagnóstico y que a su vez pudieran estar asociados con la dureza de la semilla, a saber: c.1. Morfológicos: Para el color y la forma se empleó una lupa estereoscópica. Un microscopio electrónico de barrido (Phillips XL-40) fue empleado para estudiar el tamaño (largo x ancho), las ornamentaciones de la cubierta seminal, longitud de la región hilar y longitud del estrofiolo. El término estrofiolo se refiere aquí a la depresión longitudinal de la cubierta seminal ubicada en el rafe entre el hilo y la chalaza (Werker 1980/81, Argel y Humprhyes 1983, Serrato-Valenti et al. 1993). Este término (estrofiolo) es equivalente a la palabra Lens empleada comúnmente por otros autores (Ballard 1975, Werker 1980/81, Argel y Humphryes 1983, Serrato-Valenti et al. 1993). A tal fin el material fue fijado en un portamuestras y metalizado con oro-paladio a 40 nm. Las observaciones se realizaron entre cinco y 10 kilovoltios, para luego obtener las respectivas impresiones fotográficas. c.2. Anatómicos: Grosor de la cubierta seminal, presencia y grosor de la cutícula, grosor de la epidermis, forma y tamaño de las osteoesclereidas (hipodermis), grosor del mesófilo, hilo (longitud de las células invertidas y tamaño de la isla de traqueidas), estrofiolo (largo de las células del tapón estrofiolar). Algunas semillas fueron a su vez fijadas en FAA por 24 horas, luego deshidratadas en una serie creciente de terbutanol e incluidas en parafina con punto de fusión 56-58ºC. Mediante el uso de un microtomo de rotación se obtuvieron secciones transversales de un grosor de 13 *m para posteriormente proceder a la tinción de las muestras con safranina-fastgreen (Johansen 1940) y su observación con un microscopio óptico a 4, 10 y 40X.
d) Determinación de las características histoquímicas: Para el estudio histoquímico fueron empleadas semillas previamente ablandadas por calentamiento (tres minutos en agua hirviendo). Las secciones se realizaron a mano alzada y de inmediato se aplicaron los reactivos señalados en el Cuadro 2. Las muestras empleadas para el reconocimiento de calosa y polisacáridos fueron sometidas a una extracción en serie de pectinas y de hemicelulosa empleando para ello oxalato de amonio al 0.5% por seis horas a 90 ºC e hidróxido de sodio al 4% por seis horas a 25 ºC (modificado de Bevilacqua et al. 1987). Las muestras fueron luego observadas en un microscopio de luz polarizada a 25X.
El diseño experimental empleado para comparar las distintas líneas fue un factorial 12x2 con efecto anidado. Los doce factores considerados fueron las doce líneas (siete diploides y cinco tetraploides) con dos niveles: posición: distal y basal. Cuando se compararon los dos niveles de ploidia el diseño fue igualmente un factorial, pero en este caso 2 x 2 y también con efecto anidado. En ambos casos se utilizó un arreglo completamente aleatorizado. Los resultados fueron sometidos a pruebas de normalidad y varianza y, cuando no se obtuvo normalidad en los datos obtenidos, se recurrió a la prueba no paramétrica de Kruskal-Wallis (Zar 1984).

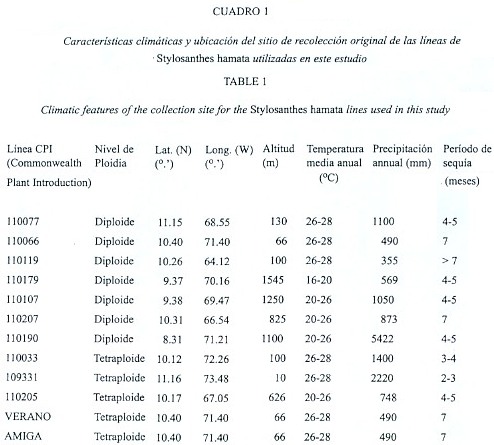
Resultados
Dureza de la semilla en función a la viabilidad: La Figura 1 muestra los porcentajes de dureza obtenidos para las doce líneas de S. hamata estudiadas. El promedio de dureza para las líneas diploides fue de 94.7% + 6.0 en la semilla distal, con una de variación entre 88% (CPI 110179) y 100% (CPI 110077, 110107). En la semilla basal el promedio de dureza fue de 88.9% + 11.5, con valores oscilando entre 64 (CPI 110179) y 97% (CPI 110077, 110190). En las líneas tetraploides la semilla distal alcanzó un porcentaje de dureza de 98.5% + 2.1 y la basal de 97.8% + 2.5, con un ámbito de variación para las dos posiciones entre 95% (cv. Amiga) y 100% (CPI 110033, 110205). No se encontraron diferencias estadísticamente significativas entre niveles de ploidia ni entre posiciones dentro del fruto, aunque el análisis estadístico demostró un clara interacción entre estos dos factores. Dentro de las líneas solo se encontraron diferencias significativas entre posiciones para CPI 110077 (p=0.005) y CPI 110179 (p=0.003), teniendo en ambos casos la semilla distal un mayor grado de dureza que la basal.
Sitio de entrada de humedad: En todas las semillas blandas, independientemente de su nivel de ploidía como de su posición en el fruto, la entrada de humedad se llevó a cabo a través del hilo, puesto que al colocar la semilla en la solución de sulfato ferroso de amonio, fue ésta la única región de la cubierta seminal ennegrecida por el contacto con la solución. Por otra parte, en las semillas duras no se observó cambio en la coloración de la cubierta seminal.
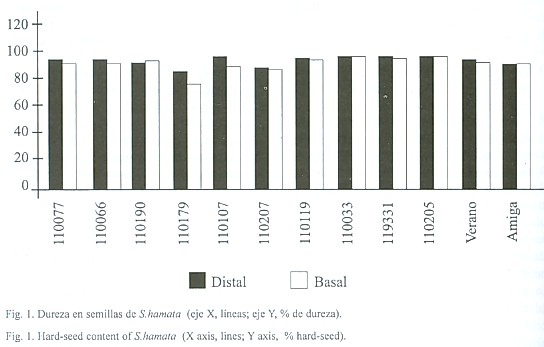
Características morfológicas de la semilla: El comparar la semilla de las distintas líneas y posiciones permitió determinar que la semilla de S. hamata es una semilla polimórfica, fue en general de forma ovoidea, cuyas dimensiones estuvieron por debajo de los 2 mm de largo por 1 mm de ancho por lo que se considera pequeña (Fig. 2) y su color varió de castaño claro a castaño oscuro casi negro. Se apreció una superficie rugosa, observándose una mayor rugosidad y mayor compactación celular en las semillas distales que en las basales (Fig. 2). ]]>
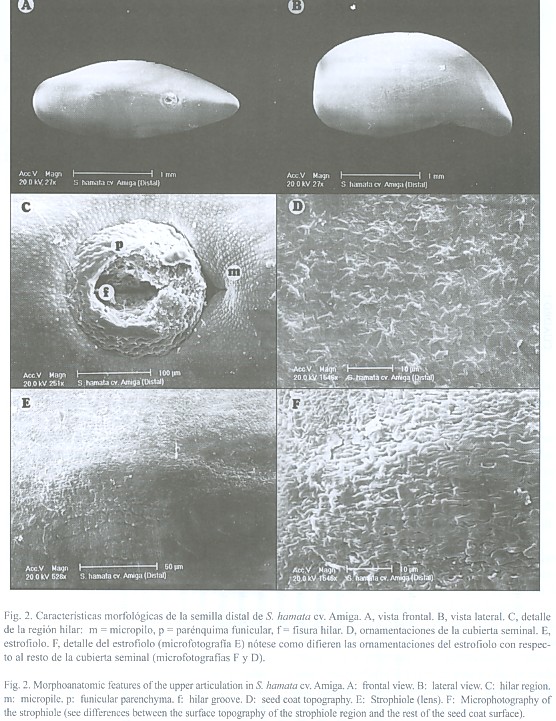
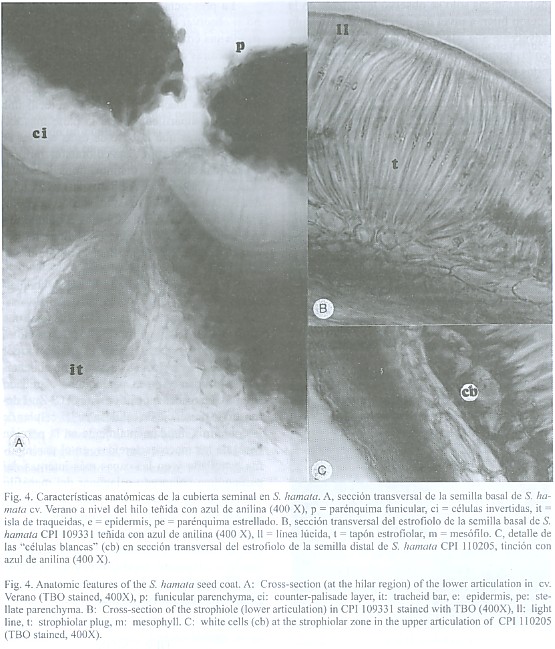
En la región hilar destacaron: a) el micropilo, el cual se observó como una depresión deltoide, generalmente cerrado y ubicado por encima del hilo; b) el hilo, ubicado en la porción apical, de color marrón oscuro y de forma circular, parcialmente cubierto por restos de parénquima funicular, sin halo distinguible y con un canal central cerrado, y c) el estrofiolo, de color castaño, más oscuro que el resto de la cubierta seminal ubicado en la porción opuesta al micropilo. El estrofiolo se destacó por ser una estructura prominente y de forma elipsoidal. En ésta última estructura la superficie de las células fue totalmente diferente al resto de la cubierta seminal. En tal sentido se observó que las macroesclereidas del estrofiolo no presentaron rugosidad.Características anatómicas de la cubierta seminal: Se observó una cutícula delgada y una epidermis uniestratificada de macroesclereidas en empalizada o células de Malpighi, con paredes engrosadas y radialmente alargadas. En la misma destacó la presencia de la línea lúcida, la cual está ubicada en la parte más externa de las macroesclereidas. La línea lúcida se apreció en toda la epidermis, a nivel del estrofiolo y en la región hilar. La misma fue distinguible tanto con el microscopio óptico como con el electrónico de barrido (Fig. 3). En las líneas tetraploides la línea lúcida se apreció más que en las diploides.
La hipodermis esta constituida por osteoesclereidas o células columnares o de reloj. Es uniestratificada y posee células de paredes engrosadas con grandes espacios intercelulares. Las osteoesclereidas de las líneas tetraploides se apreciaron menos estilizadas, con menores espacios intercelulares y menos ornamentadas. Así mismo, las semillas distales presentaron una hipodermis más gruesa que las semillas basales (Fig. 3).
El mesófilo conformado por varias capas de parénquima lagunar o colapsado, se encuentra constituido por células tangencialmente elongadas y de paredes engrosadas. En las líneas tetraploides se apreció que esta capa de tejido es más delgada que en las líneas diploides (Fig. 3).
El hilo presentó una doble capa de células de Malpighi, la superior invertida y superpuesta sobre la inferior. Ambas capas se ubican a los lados del canal central (fisura hilar) el cual se apreció totalmente cerrado. En la semilla distal de las líneas tetraploides se observó una tendencia a presentar esta estructura en forma más desarrollada. La isla de traqueidas ubicada por debajo del canal central se apreció más alargada en la semilla distal de las líneas tetraploides (Fig. 4).
El estrofiolo estuvo constituido por macroesclereidas mucho más largas y delgadas que el resto de las macroesclereidas que conforman la cubierta seminal. Se destacó la presencia de pequeñas células bien diferenciadas ubicadas a los lados de las macroesclereidas que conforman el tapón estrofiolar. Estas células denominadas células blancas (Fig. 4) fueron descritas por Serrato-Valenti et al. (1995) en el estrofiolo de la semilla de Leucaena leucocephla.
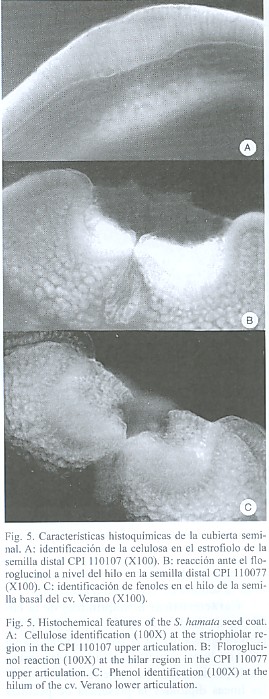
La presencia de polisacáridos se determinó al observar con el microscopio de luz polarizada una coloración roja en la isla de traqueidas mientras que las células invertidas se tiñeron de color verde y el parénquima funicular y el parénquima estrellado de color anaranjado a rojo. En las macroesclereidas se apreció una coloración amarillenta hacia la parte más externa mientras que la base de estas células se observó de color rojo. Esta coloración múltiple reveló la presencia de distintos tipos de polisacáridos en la cubierta seminal. Las observaciones realizadas con el microscopio de luz polarizada indicaron que la calosa es el compuesto que determina la presencia de la línea lúcida, encontrándose que en las líneas tetraploides la reacción fue mucho más intensa.
La lignina se detectó en la isla de traqueidas al observarse una coloración de rojo a morado y en las células invertidas al obtenerse una coloración rosada intensa (Fig. 5). Los fenoles estuvieron presentes en toda la epidermis por debajo de la línea lúcida, excepto a nivel del tapón estrofiolar y en el parénquima estrellado (Fig. 5). La reacción positiva ante el Cl-Zn-I demostró la presencia de celulosa en las células de la epidermis, fundamentalmente en la porción basal de las macroesclereidas, en el parénquima estrellado y en las capas más internas del parénquima colapsado o lagunar del mesófilo (Fig. 5). De igual manera se observó la presencia de este compuesto en la hipodermis.
Con respecto a la reacción de las muestras ante el sudan III y el sudan IV, sólo se obtuvo una leve coloración en el parénquima colapsado del mesófilo revelando así la presencia de grasas y cutina en este tejido.
Discusón
La presente investigación ha demostrado que la semilla de S. hamata (L.) Taub. desarrollada en condiciones tropicales de altas temperaturas (> 25 ºC) presenta un alto grado de dureza (>85%) y que el porcentaje de dureza en la semilla fue, en la mayoría de los casos, independiente del nivel de ploidia y de la posición de la semilla en el fruto (Fig. 1). Estos resultados confirman los obtenidos por Argel y Humphreys (1983) y Guenni et al. (1994). Sin embargo, se observó una tendencia en algunas líneas diploides a presentar un porcentaje mas bajo de semillas duras, tal como fue el caso de la línea CPI 110179 cuyos valores de dureza en la semilla basal fueron inferiores al promedio obtenido para la especie.
Desde un punto de vista ecológico, este tipo de latencia representa una propiedad adaptativa que promueve la sobrevivencia de las plantas bajo condiciones adversas y por lo tanto, es de suma importancia en términos evolutivos. La misma representa una ventaja para la especie cuando las condiciones ambientales para la germinación no son las más favorables. La dureza seminal es en consecuencia un mecanismo por medio del cual la germinación puede distribuirse tanto en el tiempo como en el espacio, aumentando así las posibilidades de éxito en el establecimiento de poblaciones sucesivas de plantas dentro de una misma especie (Guenni 1992). Este tipo de latencia asegura entonces, la permanencia de la semilla en el banco de semillas del suelo y la supervivencia de la especie a largo plazo (Bewley & Black 1982, Rice 1989).
En S. hamata, al igual que en otras especies de leguminosas, este tipo de latencia está altamente asociada a las características morfo-anatómicas e histoquímicas de las estructuras o tejidos que conforman la cubierta seminal. ]]>
En el presente estudio el hecho de haberse encontrado para S. hamata un conjunto de características particulares en la cubierta seminal, le confieren a la semilla su carácter impermeable, tal y como lo han informado distintos autores para otras leguminosas (Werker et al. 1979, Hanna 1983, Serrato-Valenti et al. 1989, Serrato-Valenti et al. 1992, Serrato-Valenti et al. 1994, Serrato-Valenti et al. 1995). Entre estas características se destacan la presencia de un micropilo e hilo totalmente cerrados y una epidermis rica en compuestos hidrofóbicos como los fenoles y conformada a su vez por macroesclereidas en empalizada muy compactadas. Igualmente se destacó en la epidermis la presencia de una línea lúcida producto de la deposición de calosa en las paredes más externas de este tejido. Aunado a esto, la existencia de compuestos hidrofóbicos tales como lignina, polisacáridos y pectinas fenólicas en la isla de traqueidas, y la presencia de lignina y calosa en las células invertidas contribuyen a determinar el carácter impermeable de la cubierta seminal. Resultados similares fueron infromados por Argel y Humphreys (1983) en S. hamata cv. Verano. En este cultivar el alto nivel de dureza en la semilla se asoció igualmente a un mayor desarrollo de la cutícula y una relación hemicelulosa/celulosa mucho más alta en las células de la epidermis. En S. scabra, Serrato-Valenti et al. (1993) observaron igualmente un patrón de composición química y de configuración anatómica de la cubierta seminal muy semejante al encontrado en este estudio.Por otra parte, se pudo determinar que la semilla de la especie estudiada posee un polimorfismo marcado en cuanto al tamaño y color. Este polimorfismo fue observado tanto entre líneas diploides y tetraploides, como entre semillas distales y basales dentro de una misma línea. La presente variabilidad fenotípica relacionada con la morfología de la semilla también se ha observado en Vigna ungiculata (Lush & Evans 1980), S. hamata cv. Verano (Argel & Humphreys 1983) y Lupinus angustifolius (Serrato-Valenti et al. 1989). Sin embargo, en S. hamata esta característica no parece estar asociada a la dureza, pues la tendencia general observada fue que independientemente del tamaño de la semilla, tanto las de color claro como las de color oscuro se comportaron como semillas duras.
Agradecimiento
Esta investigación formó parte de la Tesis de Maestría de la primera autora y fue parcialmente financiada por la Fundación para el Desarrollo de la Ciencia y la Tecnología del Estado Aragua, (FUNDACITE-Aragua), Venezuela. Agradecemos a los dos revisores anónimos por sus valiosos comentarios en la elaboración final del manuscrito.
Resumen
La semilla de Stylosanthes hamata (L.) Taub. se caracteriza por presentar un alto grado de latencia asociada a una restricción a la absorción de humedad (semilla dura). En esta investigación, se estudió la dureza de semillas distales y basales de siete líneas diploides y cinco líneas tetraploides, cultivadas bajo condiciones tropicales. Las pruebas pruebas de germinación (HR: 55-65%, temperatura constante: 27-30º C, fotoperíodo: 12/12 hr) llevadas a cabo en semillas desprovistas de vaina indicaron que el grado de dureza seminal estuvo en casi todos los casos por encima del 85%, independientemente del nivel de ploidia y de la posición de la semilla en el fruto. Así mismo, se realizó una caracterización morfo-anatómica de la cubierta seminal a través del uso del microscopio óptico y de barrido. Esta caracterización fue complementada con un estudio histoquímico para determinar la ubicación de compuestos hidrofóbicos en la cubierta seminal. Los resultados de estas pruebas señalaron que la dureza seminal estuvo relacionada con la presencia de un micropilo y un hilo totalmente cerrados, una epidermis compuesta por macroesclereidas en empalizada con una línea lúcida en las paredes más externas de estas células y a una alta concentración de compuestos hidrofóbicos en todos los tejidos que conforman la cubierta seminal. ]]>
Ballard, L.T.A. 1973. Physical barries to germination. Seed. Tech. 1: 285-303 [ Links ]
Baskin, C. &J. Baskin. 1989. Physiology of dormancy and germination in relation seed bank ecology. p. 553-556. In:M.A. Leck, V.T. Parker & R.L. Simpson (eds). Ecology of soil seed banks. California.
Bevilacqua, L., F. Fossati & G. Dondero. 1987. Calosa in the impermeable seed coat of Sesbania punicea. Ann. Bot. 59:335-341 [ Links ]
Bewley, J.D. & M. Black. 1982. Physiology and biochemistry of seeds in relation to germination. Vol. 2. Viability, dormancy and environmental control. Springer-Verlag, Berlín. 375 p. [ Links ]
Burt, R.L., D.G. Cameron, D.F. Cameron, L. TMannetje & J. Lenne. 1983. Stylosanthes. p. 141-181. In:R.L. Burt, P.P. Rotar, J.L. Walker & M.W. Silvey (eds.) The role ofCentrosema, Desmodium and Stylosanthes in proving tropical pastures. Colorado.
Castillo, R. 1997. Estudio de la dureza en semillas de Stylosanthes hamata (L.) Taub. y su realación con las características morfo-anatómicas de la cubierta seminal. Tesis de Maestría. Facultad de Agronomía. Universidad Central de Venezuela. Maracay. Venezuela. ]]>
Edye, L.A. & B. Groft. 1983. Selecting cultivars from naturally ocurring genotypes:evaluating Stylosanthes species. p. 217-232. In:J.G. McIvor y R.A. Bray (eds). Genetic resources of forage plants. CSIRO, Melbourne. [ Links ]
Edye, L.A. 1988. Stylosanthes collecting in Colombia and Venezuela. 16/01/86 to 05/09/86. Final Report. CSIRO, Townsville, Australia. 28 p. [ Links ]
Escala, M. 1994. Estudio morfoanatómico de frutos y semillas de leguminosas de los altos llanos centrales de Venezuela (Estación Biológica de los Llanos de la Sociedad Venezolana de Ciencias Naturales) Bol. Soc. Venezolana. Cien. Natur. 44(148): 259-316.
Ferder, N & T.P. OBrien. 1968. Plant microtechnique, some principles and new methods. Amer. J. Bot. 55 (1): 123-142. [ Links ]
Gardener, C.J. 1975. Mechanism regulating germination in seed of Stylosanthes. Aust. J. Agric. Res. 26: 281-294 [ Links ]
Guenni, O. 1992. Phenotypic variability within Stylosanthes hamata in relation to seed dormancy, growth and reproductive hehaviour. PhD Tesis. Griffith University. Australia. p. 86-123 [ Links ]
Guenni O, D.F. Cameron, L.A. Edye & C. Rose. 1994. Germinación y dureza en Stylosanthes hamata (Leguminosae). Ecotrópicos. 7(1): 1-12. [ Links ]
Hanna, P.J. 1983. Anatomical features of the seed-coat of Acacia kempeana (Muelier) which relate to increaseed germination reta induced by heat treatment. New Phytol. 96: 23-29. [ Links ]
Jensen, W.A. 1962. Botanical histochemistry. W.H. Freeman and Company. San Francisco. P. 23-56. [ Links ]
Johansen, D.A. 1940. Plant microtechnique. McGraw-Hill.New York. p. 14-45 [ Links ]
Lascano, C.E. 1991. Managing the grazing resource for animal production in savannas of tropical america. Trop. Grasslands. 25: 66-72 [ Links ]
Lascano, C.E. & C. Plazas. 1990. Banco de proteina y energía para complementar sabanas en los llanos de Colombia. Anais da 12a. Reuniao da Alpas. 22-27 Julho. Campinas, Sao Paulo, Brazil. p. 117
Lush, W.M. & L.T. Evans. 1980. The seed coats of Cowpeas and the order grain legumes:Structure in relation of the funtion. Fiel Crops Res. 3: 267-286. [ Links ]
Mott, J.J. 1979. High temperature contac tratament of hard seed in Stylosanthes. Aust. J. Agric. Res. 30: 845-874. [ Links ]
Rice, K.J. 1989. Impacts of seed banks on grassland community structure and population dynamics. p. 211-230. In:M.A. Leck, V.T. Parker & R.L. Simpson (eds). Ecology of soil seed banks. Chap. 10. Part 3. California.
Roth, I. 1964. Microtecnia vegetal. Ediciones de la Biblioteca de la Universidad Central de Venezuela. Caracas. p. 17-34.
Schneider, H. 1973. Methods for Botanical Sciences. p. 192-212. In:G. Clark (eds) Staining procedures used by the Biological stain Commision. 3th edition. The Williams & Wilkins Co. Baltimore, USA.
Serrato-Valenti. G., L. Melone, F. Ferro & A. Bozzini. 1989. Comparative studies on the testa structure of hard-seed and soft-seed varieties of Lupinus angustifolius L. (Leguminosae) and mechanims of water entry. Seed Sci. & Technolog. 17: 563-581 [ Links ]
Serrato-Valenti,G. L. Cornara, S. Lotito & L. Quagliotti. 1992. Seed coat structure and histochemistry of Abelmoschus esculentus chalazal region and water entry. Ann. Bot. 69: 313-321 [ Links ]
Serrato-Valenti, G. L. Cornara, M. Ferrando & P. Modensi. 1993. Structural and histochemical features of Stylosanthes scabra (Leguminosae-Papilionoideae) seed-coat as related to water entry. Can. J. Bot. 71: 834-840 [ Links ]
Serrato-Valenti, G. L. Cornara, P. Ghisellini & M. Ferrando. 1994. Testa structure and histochemistry related to water uptake in Leucaena leucocephala Lam (De Wit). Ann. Bot. 73: 513-537. [ Links ]
Serrato-Valenti, G. M. De Vries & L. Cornara. 1995. The region hilar in Leucaena leucocephala Lam (De Wit) seed:structure, histochemistry and the role of lens in germination. Ann. of Bot. 73:569-574 [ Links ]
Werker, E. 1980/81. Seed dormancy as explained by the anaomy of embryo envelopes. Israel J. Bot. 29: 22-44 [ Links ]
Werker, E., I. Marbach & A.M. Mayer. 1979. Relation between the antomy of the testa, water permeability and the presence of phenolics in the genus Pisum. Ann. Bot. 43: 765-771 [ Links ]
Williams, R., R. Reid, R. Schultze-Kraft, N.M. Sousa-Costa & B.D. Thomas. 1984. Natural distribution of Stylosanthes. p. 73-101. In:H.M. Stace & L.A. Edye (eds). The biology and agronomy of Stylosanthes. Sydney.
Zar, J.H. 1984. Biostatistical Analisys. Second edition. Prentice-Hall International,New Jersey. 718 [ Links ]p.
1 Universidad Simón Bolívar. Departemento de Estudios Ambientales. Apartado postal 89000, Caracas 1080, Venezuela. Fax 58-2-9063064. e-mail: mailto:rosanacastillo@hotmail.com ]]>