











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink1. Introducción
Las abejas sin aguijón (ASA) se encuentran en las zonas tropicales y subtropicales del mundo, principalmente en Sur América, donde se halla la mayor diversidad de especies (Prendas, 2013). En total se conocen cerca de 400 especies (Freitas et al., 2009), de las cuales 300 se encuentran en el Neotrópico, desde México hasta Argentina; en Costa Rica se han identificado al menos 20 géneros y 59 especies, lo cual representa cerca del 15 % de la diversidad mundial de estos insectos (Aguilar, 2013; Figueroa-Mata et al., 2016).
Las ASA juegan un importante papel en la polinización de las plantas y en la agricultura en particular (Araújo et al., 2004; Batista et al., 2016, Frankie et al., 2018); su labor polinizadora contribuye a la formación de frutos y semillas y, por tanto, a la provisión de alimento para la fauna frugívora, tanto silvestre como doméstica e incluso para poblaciones humanas (Sparagnino et al., 2014). Son, además, una importante alternativa al uso de las abejas Apis mellifera en la polinización comercial de cultivos agrícolas (Slaa et al., 2006).
Se desconocen las razones por las cuales las abejas sin aguijón seleccionan un determinado sitio para establecer sus nidos o la densidad de colonias que pueden desarrollarse de manera adecuada en un espacio determinado. Varios estudios de esos tópicos concluyen que, en el caso de los insectos sociales que con frecuencia viven en colonias permanentes y muy pobladas, la densidad de colonias locales puede verse limitada por la disponibilidad de sitios adecuados para anidar (Eltz et al., 2002); a cambio, igualmente, reciben el beneficio de la polinización (Armbruster, 1984, citado por Roubik, 2006). Wille (1983) menciona que la mayoría de los árboles de las selvas tropicales no son polinizados por el viento, sino por otros agentes, entre ellos las abejas que juegan un papel importante; él pudo comprobar la presencia de ASA polinizando árboles del dosel en Corcovado, Costa Rica e indica que, sin ellas, las comunidades de árboles de la selva tropical serían muy modificadas.
Vergara et al. (1986) mencionan que la mayoría de las especies de ASA prefiere árboles vivos para establecer sus nidos y la mayor altura a la que encontraron un nido en su estudio fue a 12 m del suelo. En relación con la densidad de colonias, Ravelo et al. (2014) estudiaron en Cuba la Melipona beechii en parcelas de 400 m2 de distintas formaciones vegetales y encontraron 1.75 nidos/ha-1 en vegetación de mogotes, 0.86 nidos/ha-1 en pinares, 1.24 nidos/ha-1 en bosque semideciduo, 0.71 nidos/ha-1 en encinares y 0.45 nidos/ha-1 en bosque de galería, concluyendo que la densidad puede estar asociada al tipo de formación vegetal, a la posibilidad de encontrar alimento, protección y alcanzar valores altos de reproducción. Por su parte, Eltz et al. (2002) encontraron en los bosques de dipterocarpáceas de Malasia una densidad de nidos alta (media de 8.4 nidos/ha-1) en los sectores que bordean manglares y densidades bajas (entre 0 y 2.1 nidos/ha-1, promedio de 0.5 nidos/ha-1) en los bosques continuos; estos datos muestran que la densidad de nidos es muy variable entre diferentes formaciones vegetales y hábitats en un mismo ecosistema.
Con respecto a las especies de árboles que las ASA utilizan para establecer sus nidos, Roubik (1983) menciona que estos insectos prefieren las especies de madera dura a las de madera suave, pero se desconoce si las abejas seleccionan las cavidades de los árboles según las especies. Ravelo et al., (2014) mencionan que las abejas sin aguijón usan un número de especies arbóreas relativamente bajo para la nidificación; Van Veen (1999) analizó 19 colonias de Melipona beecheii en Costa Rica, las cuales anidaban en siete diferentes especies de árboles, con una preferencia marcada por Gliricidia sepium (madero negro), lo cual coincide con los hallazgos de Moreno y Cardozo (1997), quienes concluyeron que, entre las especies arbóreas de importancia para M. beecheii en el establecimiento de sus nidos, se encuentran Gliricidia sepium y Bursera simaruba. Fonte et al. (2009) también determinaron una aparente preferencia de la M. beecheii por anidar en troncos de Gliricidia sepium, razón por la cual resaltan el importante papel de esa leguminosa forrajera utilizada en cercas vivas, en el mantenimiento de la biodiversidad en los sistemas de producción.
Varias investigaciones han estudiado las abejas nativas en ecosistemas de bosque seco en Guanacaste; por ejemplo, Frankie et al. (1997) responsabilizan a la destrucción del hábitat y el desarrollo agrícola como los principales factores causantes de la disminución de la riqueza y abundancia de especies de abejas nativas en sus sitios de estudio, situación que evidenciaron incluso en remanentes boscosos de áreas protegidas. Mencionan que la presencia de periodos lluviosos con baja precipitación, seguidos de períodos de sequías, en combinación con el cambio de uso del suelo, son responsables de la declinación general de las poblaciones de abejas nativas en las tierras bajas de Guanacaste. Estos datos son más preocupantes, al considerar, como lo establecen Frankie et al. (2018), que en Costa Rica hay cerca de 8 500 especies de plantas de floración y cerca de un 60 % de ellas son polinizadas por especies de abejas nativas.
El presente estudio busca documentar las interacciones entre especies de abejas meliponidas y las plantas que utilizan como sitio de anidación y alimento en tres diferentes ecosistemas del PNG, Costa Rica. Asimismo, se procedió a caracterizar la vegetación arbórea y arbustiva asociada a este importante grupo de insectos y a realizar el análisis palinológico de las cargas de polen portadas por las abejas o recolectadas en sus nidos.
2. Metodología
2.1 Sitio de estudio
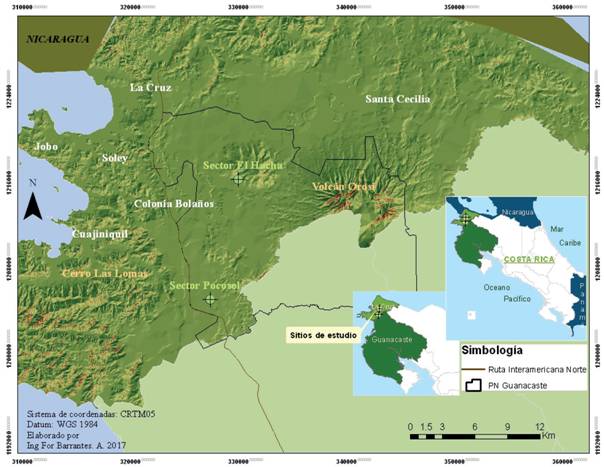
El estudio se llevó a cabo en el Área de Conservación Guanacaste (ACG), Parque Nacional Guanacaste (PNG), en los sectores Pocosol y cerro El Hacha (Figura 1). En el sector Pocosol se trabajó en bosque secundario (BS) en diferentes estados de sucesión; se tiene referencias de que esos sectores fueron abandonados desde hace 39 y 56 años, lo cual propició su proceso de regeneración. Se ubica en las coordenadas geográficas N 10° 88' 90'' y W 85° 58' 60'', a 255 m s.n.m., en la zona de vida bosque húmedo premontano transición a basal, según la clasificación de Holdridge (Tosi, 1969). En este sitio la precipitación promedio anual es de 1 800 mm y la temperatura media anual de 28 °C (Solano y Villalobos, s.f.); presenta un periodo seco que va de noviembre a abril (o de diciembre a mayo, dependiendo de la influencia del ENOS).
El sector cerro El Hacha presenta un bosque primario intervenido (BPI) y se ubica en las coordenadas geográficas N 10° 99' 12'' y W 85° 56' 14'‘, entre 300 y 500 m s.n.m., en la zona de vida bosque húmedo tropical, según la clasificación de Holdridge (Tosi, 1969), con una precipitación promedio anual de 1 800 mm y una temperatura media anual de 28 °C, cuyo período seco va de noviembre a abril (Solano y Villalobos, s. f.).
2.2 Disponibilidad de sitios potenciales para anidación
En el bosque secundario del sector Pocosol la topografía es plana y los suelos son entisoles; en cada sector (39 y 52 años) se inventariaron once parcelas de 20 x 20 m (400 m²) para un área total de muestreo de 0.88 ha. Por su parte, en el cerro El Hacha se muestrearon once parcelas de 400 m2 (0.44 ha) en lo que corresponde al bosque primario intervenido (Figura 2a).
Dado que se cuenta con parcelas permanentes de muestreo (PPM) en bosques secundarios de dos diferentes estados sucesionales, se buscó establecer si existen diferencias en las características del bosque y en la presencia, tanto de nidos como de especies de ASA. Por tal razón, se procedió a documentar las características dasométricas de todos los individuos, según el protocolo establecido para este fin (Sánchez, 2013).
En cada parcela se midió el diámetro a 1.3 m de altura (DAP) de todos los individuos con dap ≥ 10 cm (árboles); se registró si se encontraba vivo o muerto en pie y se verificó la información taxonómica, colectando muestras botánicas en caso de que fuera necesario corroborar la taxonomía (Figura 2b). Se remidieron todos los árboles que ya estaban marcados y mapeados, y se incluyeron los nuevos ingresos, o sea, aquellos árboles que para esta medición ya habían alcanzado los 10 cm de DAP.
2.3 Densidad y distribución espacial de nidos, abejas y colecta de polen
Para cada árbol censado con la metodología descrita en la sección anterior, se determinó, visualmente, la presencia de nidos. Cada nido se georreferenció por medio de un receptor GPS marca Garmin, modelo GPSmap 60CSx, y se registró información como la especie de ASA, altura sobre el tronco a la cual se encontraba el nido, tipo y altura de la piquera (entrada del nido) (Figura 2c).
Mediante la utilización de redes entomológicas se colectaron abejas en la entrada de los nidos, las cuales fueron colocadas en pequeños tubos (viales) y trasladadas al laboratorio para su identificación taxonómica, mediante la utilización de claves y comparación con especímenes de la colección de referencia de abejas del CINAT.
Para la determinación de las plantas alimenticias utilizadas por las abejas, se realizó un muestreo en tres nidos de la especie Scaptotrigona pectoralis ubicados en los alrededores de las PPM del cerro El Hacha; lo anterior para tener más representatividad en el muestreo, al estar ubicados los nidos de esta especie cercanos entre sí y accesibles en un mismo sector. El muestreo se realizó en la época seca, en tres visitas al área de estudio durante los meses de febrero, marzo y abril de 2015. Durante cada visita se recolectaron 20 abejas obreras por nido. La mitad en un muestreo matutino de las 8:00 a las 11:00 h y la otra mitad en uno vespertino de 15:00 a 18:00 h. Las abejas fueron colectadas durante su llegada al nido, para lo cual se verificó que tuvieran polen en las corbículas. Un total de 180 muestras fueron colectadas (Figura 2d).
Durante la época lluviosa se muestrearon abejas obreras pecoreadoras en el sector Pocosol, en dos nidos de la especie Trigona fulviventris ubicados igualmente cercanos entre sí y accesibles para verificar el muestreo durante el mismo periodo de tiempo. Estos se muestrearon durante los meses de setiembre y octubre del año 2015, coincidiendo en este caso con el periodo de lluvias. Se colectó un total de 30 abejas por nido al día (tres días en setiembre y dos en octubre), con una distribución aproximada de la mitad de las abejas colectadas entre las 7:00 y las 10:00 h y el resto de las 14:00 a las 17:00 h, ya que su actividad de pecoreo tiende a disminuir en las horas en que el sol se encuentra en su punto más alto, debido a la elevación de la temperatura (Aguilar y Smith, 2009).
Adicionalmente, en cada una de las visitas, coincidiendo con los periodos de muestreo de abejas, se colectó el polen de plantas en flor dentro de las PPM, para lo que se utilizó, como área de muestreo, transectos aleatorios alrededor de los nidos, tomando, como referencia promedio, un radio óptimo de 500 m, acorde con la actividad de pecoreo de las abejas. Este polen, a su vez, sirvió como material de referencia para su comparación con el polen que se clasificó a partir de las cargas portadas por las abejas.

2.4 Análisis palinológico
Para determinar las especies de flora de las cuales las abejas portaban su polen, se utilizaron métodos de análisis palinológico que permitieron establecer, mediante las características morfológicas del grano de polen (polaridad, tamaño, aberturas en la exina y tipo de ornamentación), la ubicación taxonómica a nivel de familia, género y especie. Dichos resultados fueron verificados mediante la comparación de las microfotografías de los granos con las claves taxonómicas de granos de polen del neotrópico, así como la colección de referencia de granos de polen de las plantas del sitio y la del CINAT. Los granos de polen fueron procesados mediante la utilización del método de tinción con safranina (Sánchez, 2001). Este método utiliza las cargas de polen colectadas por las abejas en sus corbículas. Asimismo, en el caso del muestreo de T. fulviventris, se modificó dicho método, optando por embeber a la abeja por completo en una solución de jalea de glicerina, previamente teñida con safranina para extraer cualquier grano de polen adherido a su cuerpo; la solución fue colocada en láminas fijas de vidrio y homogenizada mediante calor, para su posterior observación e identificación del polen al microscopio. Esta modificación se realizó debido a que durante la época de muestreo no se observaron cargas de polen corbiculares en esta especie de abeja.
Para el análisis del polen colectado de las plantas en flor, se realizó una disección y separación de las anteras y estructuras florales de cada una de las especies colectadas e identificadas con anterioridad; las anteras se colocaron en cajas petri rotuladas para evitar la combinación del polen de cada planta, se introdujeron en un beaker con 15 ml de agua destilada para llevar a ebullición a una temperatura de 400 °C, con el propósito de separar los granos de polen de las anteras. Posteriormente se realizó una filtración de la solución para separar las estructuras florales de mayor tamaño, la solución filtrada se vertió en tubos de ensayo utilizando una balanza para igualar volúmenes antes de centrifugar durante siete minutos a 3 500 rpm, para luego eliminar el líquido sobrenadante por decantación de un golpe; este paso se repitió dos veces para recuperar la mayoría del polen por sedimentación. Adicionalmente, se preparó una solución de glicerol (50 % glicerina + 50 % agua, 25 ml respectivamente), la cual permite que el polen se mantenga fresco durante más tiempo, se adicionaron 10 gotas de safranina a cada tubo, para teñir las células vivas y se colocaron los tubos en un espacio carente de luz, para lograr que la solución de safranina penetrara en la pared de la célula del polen. De igual forma los tubos se centrifugaron, decantaron y dejaron en reposo en posición completamente vertical; de esta manera, los granos de polen quedan embebidos en la parte superior del tubo. Posteriormente se colocaron a secar en la estufa a 60 °C durante 15 minutos. Finalmente, con la ayuda de una aguja de disección, se frotó con porciones pequeñas de gel de glicerina en la parte superior del tubo, donde se encontraban concentrados los granos de polen y se procedió a elaborar las láminas fijas para la observación del polen al microscopio.
3. Resultados
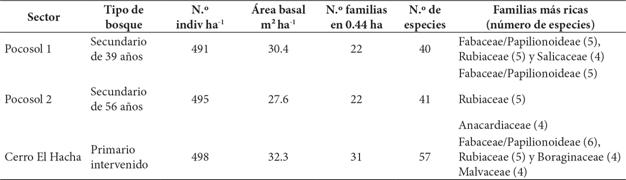
Con respecto a la composición florística y área basal de los sitios de estudio, se determinaron las características de cada tipo de bosque, según se indica en el Cuadro 1. Los datos sobre composición florística de los tres sitios en su conjunto indican la presencia de 37 familias, 70 géneros y 84 especies de árboles con DAP ≥ 10 cm, donde es evidente la riqueza de la familia Fabaceae en los tres sitios de estudio. La cantidad de especies y la riqueza por familias fue muy similar en los dos tipos de bosque secundario, a pesar de existir una diferencia de 17 años entre uno y otro. En el Apéndice 1 se muestra la lista con los nombres comunes y científicos de todas las especies arbóreas inventariadas en las tres parcelas.
Adicionalmente, en los alrededores de las parcelas, se realizaron colectas de plantas en floración, lo cual dio como resultado la presencia de 30 especies adicionales, entre ellas una especie arbórea, nueve arbustos, doce hierbas, cinco lianas herbáceas y tres lianas leñosas. La lista completa de las especies mencionadas se muestra en el Apéndice 2.
El bosque secundario de 56 años muestra una mayor cantidad de individuos por ha con respecto al bosque secundario de 35 años, pero una disminución en el área basal por ha; el bosque primario intervenido es el que presenta mayor cantidad de individuos y mayor área basal por ha. Por otra parte, en el bosque secundario de Pocosol la especie Q. oleoides es la que presenta el mayor índice de valor de importancia (IVI = 69.71), lo cual significa el 23.23 % del total del IVI e indica la importancia ecológica relativa de la especie en este ecosistema. La situación es similar en el cerro El Hacha, pues la especie Q. oleoides alcanza un IVI de 43.10, correspondiente al 14 % del IVI del bosque primario intervenido.
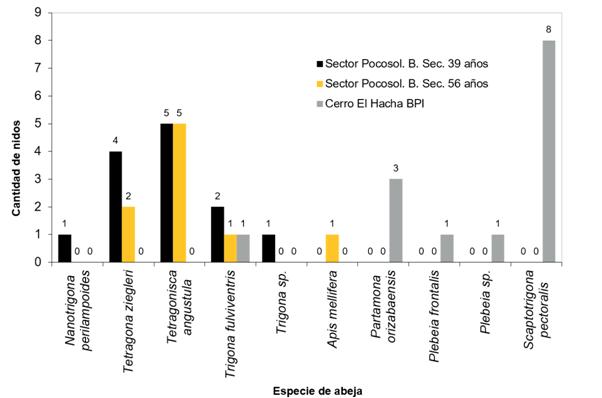
Dado que los datos de composición florística y área basal mostraron pocas diferencias en los tres sitios de estudio, se optó por analizar los datos del inventario de nidos de ASA de manera conjunta, lo cual dio como resultado que se localizaron un total de 35 nidos de abejas sin aguijón, más una colmena de Apis mellifera, para una densidad total de 4.43 nidos/ ha-1 (Figura 3).
En el bosque primario intervenido se hallaron las especies Scaptotrigona pectoralis, Partamona orizabaensis, Plebeia frontalis y Plebeia sp., las cuales fueron exclusivas de ese tipo de ecosistema; solamente una especie de ASA se ubicó en los tres tipos de bosque estudiados: Trigona fulviventris. En el bosque secundario (sectores 1 y 2 de Pocosol) se logró ubicar las especies Tetragonisca angustula, Tetragona ziegleri y Nannotrigona perilampoides (Figura 3).
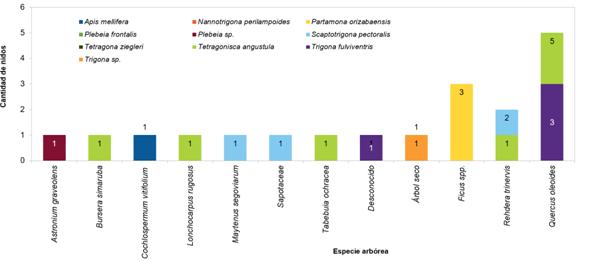
En la Figura 4 se muestra la preferencia de las abejas por las distintas especies arbóreas. El 50 % de los nidos se encontraron en el Quercus oleoides, además de ello, seis de las 10 especies de abejas encontradas los construyeron en los troncos de esta misma especie. Todos los nidos de Partamona orizabaensis encontrados fueron construidos en los troncos del Ficus sp. En troncos de árboles muertos se ubicaron tres nidos de tres especies de ASA (Tetragona ziegleri, Tetragonisca angustula y Trigona sp.). La especie arbórea Rehdera trinervis, un árbol de porte bajo, de fuste acanalado, que permite a las ASA construir sus nidos en medio de los intersticios del tronco, fue la segunda especie más utilizada, con un 14 % de los nidos. Las abejas de la especie Tetragonisca angustula utilizaron varias especies arbóreas para construir sus nidos (cinco los hicieron en el encino y uno en cada una de las siguientes especies: Bursera simaruba, Lonchocarpus rugosus, Tabebuia ochraceae, Rehedera trinervis y en un árbol de especie indeterminada, pues se encontraba seco.
Las especies de abejas con mayor cantidad de nidos (considerando los tres sitios) fueron: Tetragonisca angustula (n=10); Scaptotrigona pectoralis (n=8); Tetragona ziegleri (n=6); Trigona fulviventris (n=3) y Partamona orizabaensis (n=3).
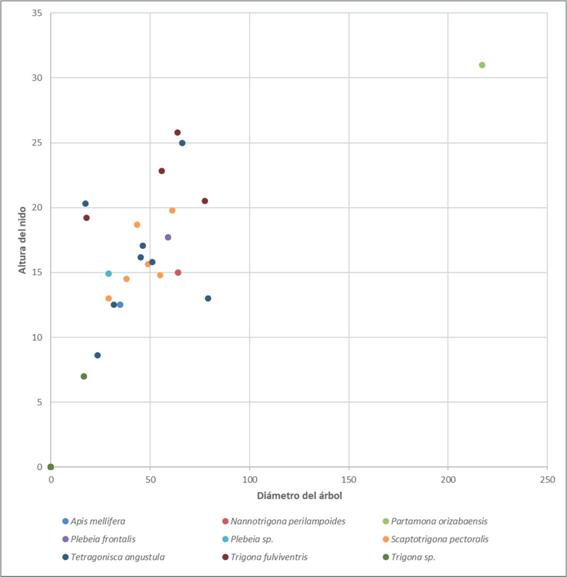
La Figura 5 muestra la selección del sitio (por especie de abeja) según el DAP del árbol donde construyeron sus nidos. La especie Partamona orizabaensis fue la que utilizó el árbol más grueso, de más de 2 m de DAP (la especie arbórea no se indica en el gráfico, pero se trató de un Ficus sp.); por su parte la especie Scaptotrigona pectoralis utilizó diferentes estratos del bosque para ubicar sus nidos, pues estos mismos se encontraron desde alturas menores a un metro hasta los 12 m de altura, resultado coincidente con lo reportado por Vergara et al. (1986). La especie T. fulviventris seleccionó alturas a nivel del suelo (tierra o raíces); los nidos de Tetragonisca angustula que se logró inventariar estaban ubicados desde el nivel del suelo hasta alturas menores a 3.5 metros.
En el estudio palinológico realizado con muestras de polen de Scaptotrigona pectoralis del cerro El Hacha y Trigona fulviventris del sector Pocosol, de las 184 muestras analizadas, 32 de ellas (17 %) no evidenciaron ningún tipo polínico presente. En los dos nidos de Trigona fulviventris se determinó un total de 26 tipos polínicos distintos, pertenecientes a 19 familias botánicas, de los cuales 14 fueron determinados a nivel de especie y los restantes 12 solamente a nivel de género. La familia que presentó mayor número de especies fue Fabaceae, subfamilia Papilionoideae, con cuatro especies, el resto de las familias visitadas por T. fulviventris estuvo representado en su mayoría por una o dos especies. La especie más representativa en los estudios de las cargas polínicas corresponde a Baltimora recta, una planta de pequeño porte, perteneciente a la familia Asteraceae, con una abundancia del 35 % para el nido 2 y cerca del 20 % para el nido 1. Los restantes recursos polínicos se encontraron en proporciones menores al 20 %. Entre los árboles identificados mediante el polen analizado se encuentran Astronium graveolens (ron-ron), Cecropia sp. (guarumo), Trema micrantha (jucó), Inga sp. (guaba), Andira inermis (almendro de montaña), Lonchocarpus sp. (chaperno), Miconia argentea (María colorada), Eugenia sp. y Triplaris sp. (hormigo). De esas especies solamente cuatro fueron inventariadas dentro de las parcelas (A. graveolens, M. argentea, Lonchocarpus sp. y Eugenia sp.).
El estudio del comportamiento de la especie Scaptotrigona pectoralis, en la época seca (meses de febrero, marzo y abril), mediante la colecta de abejas en tres nidos del BPI del cerro El Hacha permitió identificar un total de 26 morfotipos de tipos de polen en las 180 muestras recolectadas; de los 26 morfotipos de polen se logró identificar 17 a nivel de especie, 9 permanecieron a nivel de género de un total de 15 familias. Se determinó que las especies de árboles más visitados por Scaptotrigona pectoralis durante los muestreos fueron Cupania guatemalensis (cantarillo), Byrsonima crassifolia (nance), Eugenia sp., Miconia argentea, Brosimun alicastrum (ojoche) y en menos proporción Cedrela odorata (cedro) y Quercus oleoides (encino). Con respecto a la cantidad de polen encontrado por especie dentro de las diferentes muestras, se obtuvo que la mayor parte del polen en las muestras pertenece a las especies Byrsonima crassifolia (nance), Cupania guatemalensis (cantarillo) y Brosimun alicastrum (ojoche).
4. Discusión
Al analizar la composición florística de los tres sitios, se destaca la riqueza de la familia Fabaceae, lo cual es particularmente importante para la alimentación de las abejas, pues sus especies (que van desde plantas de pequeño porte, lianas, arbustos y árboles) presentan floraciones muy abundantes y constantes a lo largo del año.
La cantidad de especies y la riqueza por familias fue muy similar en los dos tipos de bosque secundario, a pesar de existir una diferencia de 17 años entre uno y otro. Esto puede estar asociado a que en el bosque seco los ciclos de crecimiento se ven limitados por la periodicidad de lluvias; lo que puede generar que existan pocas diferencias perceptibles en periodos de tiempo menores a 50 años.
La densidad de nidos encontrada fue de 4.43 nidos/ ha-1; por su parte Roubik (1983), en un estudio realizado en bosque húmedo y lluvioso tropical en Panamá, encontró 6.0 nidos/ha-1; mientras que Eltz et al. (2002) encontraron en los bosques de dipterocarpaceas de Malasia, densidades altas (media de 8.4 nidos/ha-1) en los sectores que bordean manglares y densidades bajas (entre 0 y 2.1 nidos/ha-1, promedio de 0.5 nidos/ha-1) en los bosques continuos.
En el presente estudio, en troncos del Quercus oleoides se identificaron nidos de seis especies de ASA, lo cual da indicios de una preferencia por la especie. Estos troncos usualmente presentan cavidades, las cuales podrían originarse debido a los daños causados por los recurrentes incendios forestales de la época seca. Otras especies arbóreas también utilizadas fueron Astronium graveolens (ron-ron), Bursera simaruba (indio pelado) y Ficus sp. (higuerón). Por su parte, Roubik (1983) reportó la presencia de nidos en los troncos del Brosimum alicastrum en bosque tropical lluvioso de Panamá y Ravelo et al. (2014) determinaron que Melipona beecheii, en Cuba, hace uso de un total de 15 especies de árboles para su nidificación, entre los cuales, los más frecuentemente utilizados fueron: Bursera simaruba, Guazuma ulmifolia, Psidium guajava, Mangifera indica y la palma Roystonea regia. Leal (2010, citado por Ravelo et al., 2014) también encontró una mayor cantidad de nidos en árboles de Bursera simaruba en las formaciones boscosas galería y siempreverde en Cuba. Según lo mencionado por Aguilar et al. (2013), los árboles del género Ficus son usados por las abejas sin aguijón de Costa Rica como sustrato para el establecimiento de sus colonias.
Tal como se muestra en la Figura 3, en el BPI fue donde se encontró la mayor cantidad de nidos (14); hay cuatro especies de ASA que son exclusivas del BPI del cerro El Hacha y no se encontraron en ninguno de los bosques secundarios estudiados en el sector Pocosol. La especie Scaptotrigona pectoralis, la cual es exclusiva del BPI, fue la que presentó la mayor abundancia de nidos, con un total de ocho.
En el presente estudio, la altura de construcción de los nidos fue variable; por ejemplo, los nidos de Partamona orizabaensis se ubicaron en el dosel intermedio e inferior (4 a 7 m), lo cual coincide con lo reportado por Jarau & Barth (2008), quienes los encontraron en cavidades en el suelo, en los troncos de los árboles, entre las raíces de epífitas o incluso en nidos de aves abandonados. Llama la atención la adaptabilidad de la especie S. pectoralis, la cual utilizó diferentes estratos del bosque, desde alturas menores a un metro hasta los 12 m de altura; mientras que los nidos de la T. fulviventris se encontraron en el suelo o en la base de árboles, coincidiendo con lo reportado por Roubik (2006), o por Jarau & Barth (2008) en su estudio en la región de Golfo Dulce, Costa Rica.
En el cerro El Hacha se colectaron y procesaron muestras de polen de tres nidos de Scaptotrigona pectoralis, durante el período de máxima floración (estación seca) de febrero a abril de 2015; mientras que en el sector Pocosol, las muestras de polen fueron colectadas de dos nidos de Trigona fulviventris en la época de menor floración (estación lluviosa) de setiembre a octubre de 2015.
Dado que las muestras de polen de nidos de ASA se colectaron en distintas épocas (estación seca en cerro El Hacha y estación lluviosa en sector Pocosol), esto podría estar generando diferencias muy significativas en los resultados para cada sitio, ya que la mayor explosión de floraciones, en general, se da durante la época seca (noviembre a mayo) y son pocas las especies con floración entre junio y octubre, por ejemplo, Acosmium (árbol perteneciente a la familia Fabaceae) y algunas Sapotaceas como Manilkara o Sideroxylon. Al respecto, Ramalho et al. (1990) indican que las preferencias en las colectas suelen estar influenciadas por la estacionalidad climática y la disposición del recurso floral en el tiempo.
Es importante indicar que los resultados obtenidos para Scaptotrigona pectoralis en el cerro El Hacha coinciden con otros estudios que revelan la importancia alimenticia de las familias Myrtaceae, Melastomataceae. Moraceae y Sapindaceae para las abejas sin aguijón en el Neotrópico (Ramalho et al., 1990; Wilms y Wiechers, 1997). A diferencia de lo reportado también en esas investigaciones, se encontraron pocas plantas de la familia Asteraceae con un papel relevante para las abejas en el sector cerro El Hacha, posiblemente debido a que se trata de un bosque en un estado más maduro, donde dicha familia está poco representada. En contraposición, en el sector Pocosol, para la especie Trigona fulviventris, la especie Baltimora recta, (familia Asteraceae) resultó ser la especie más abundante. En los diferentes muestreos realizados en Pocosol, esta planta herbácea apareció como recurso más importante en las colectas durante la época lluviosa, lo cual brindó un indicio sobre el comportamiento fenológico de la planta, pues al presentarse un invierno más seco, influenciado por el efecto del Niño, su periodo de floración se amplió, posiblemente como repuesta adaptativa a esa variación climática. Usualmente esta planta florece en junio y julio, alrededor del “veranillo” y luego cuando las lluvias se intensifican las flores tienden a desaparecer (Espina y Ordetx, 1983).
La cantidad de familias identificadas en el estudio palinológico coincide con los resultados obtenidos por Ferreira y Absy (2013), quienes analizaron polen de Melipona (Melikerria) interrupta en el municipio de Manacapuru, Estado de Amazonas, Brasil y, en las muestras de residuos evaluadas, identificaron 32 tipos polínicos, que representaban 19 familias botánicas, y tres tipos de polen no estaban determinados. Al igual que en el presente estudio, Ferreira y Absy (2013) encontraron que la familia más representativa fue Fabaceae (Mimosoideae), con ocho tipos de polen.
5. Conclusiones
Desde el punto de vista florístico, la familia botánica dominante en las tres condiciones de bosque fue la Fabaceae, la cual es de gran relevancia para el ecosistema por su abundante floración, utilizada por las abejas sin aguijón como fuente de polen y néctar; aspecto que se logró comprobar, pues de los tipos polínicos identificados que portaban las abejas, cuatro pertenecían a esta familia.
La especie Quercus oleoides fue la preferida por las abejas sin aguijón como sitio de anidación, es además la que presenta el mayor IVI en todos los sectores y los mayores DAP en las PPM, por lo que cumple un papel importante para la conservación de las poblaciones de abejas sin aguijón en estos bosques.
En el caso de la Scaptotrigona pectoralis, se sugiere la existencia de una especificidad cuantitativa y cualitativa en las preferencias de visita floral de la especie. Por su parte, para Trigona fulviventris no es posible asegurar una especificidad en la dominancia presente en las colectas para la época del año analizada, en lo relativo a la posible relación entre esta especie y las plantas más representativas del estudio, ya que las limitaciones de recursos nutricionales que se presentaron en el sitio, en conjunto con las necesidades alimenticias de la población de las colonias, obliga a las abejas a recurrir a múltiples fuentes de nutrición.
El conocimiento generado en este estudio sobre las poblaciones de abejas sin aguijón en el ACG y su asociación con el hábitat se convierte en una herramienta para la determinación de la vulnerabilidad de estos insectos en los escenarios que se presentan, producto de la variabilidad ambiental y la acción antropogénica directa. Además, se podrán generar, en el futuro, criterios de uso sustentable y sostenible para las abejas en su entorno y favorecer, así, que continúen brindando los servicios ecosistémicos de polinización al bosque seco tropical.

