











 Português (pdf)
Português (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntrodução
A competição arbórea é um fenômeno biológico que influencia a dinâmica de crescimento e produção em ecossistemas florestais. Informações sobre a competitividade subsidiam o planejamento e definição de estratégias conservacionistas para o estímulo da formação lenhosa em fragmentos sob recuperação e manejo florestal sustentável.
O estoque de crescimento lenhoso reflete os efeitos cumulativos da competição, influenciado pelas condições de sítio (aspectos climáticos, edáficos, topográficos, nutricionais) (1), (2), (3). A intensidade da competição por sua vez, oscila conforme a relação espacial entre árvores, disponibilidade de recursos para árvores competidoras e da plasticidade arquitetônica e fisiológica vegetal (1), (4). Contudo, ainda, persistem dúvidas sobre a demarcação da zona de influência competitiva de árvores, especialmente em fragmentos degradados de Mata Atlântica.
Diante da complexidade das interações bióticas e abióticas envolvidas na expressão da capacidade competitiva lenhosa, a competição tem sido convencionalmente resumida na forma de um único valor ou índice (5), (6), (7), (8). Os índices de competição são relações algébricas que refletem o grau de supressão de cada árvore em relação àquelas vizinhas competidoras, úteis para a quantificação de competitividade em um dado ambiente, normalmente, calculados para cada árvore em uma comunidade (8), (9), (10), (11), (12).
Diversos índices de competição se encontram disponíveis na literatura (2), (10), (11), (13), (14), porém sua escolha é um grande desafio de gestão e deve ser vista com cautela. É improvável que haja um índice ideal para todos os sítios florestais (1), mas se recomenda o uso daquele que seja mais consistente biologicamente (7). Índices dependentes da distância se baseiam em dados de espacialização e dimensões entre árvores, ou seja, exprimem a ligação mecanicista entre o tamanho, abundância e distribuição espacial das árvores vizinhas (8), (15). Esta característica permite a análise da competitividade assumindo distintos raios de competição, subsidiando interpretações sobre interações entre árvores quanto a aquisição de recursos limitados (3), (16).
Duas abordagens têm sido utilizadas para a definição do raio de competição, o método de raio fixo e o método de teste com diferentes tamanhos de raio. O método de raio fixo é uma abordagem amplamente adotada que considera empiricamente um valor de raio e/ou utiliza a referência de outros trabalhos. No entanto, é plausível assumir que tal raio pode ser distinto entre ecossistemas devido a sua variabilidade estrutural (2), (9), (14). A quantificação do efeito competitivo de árvores vizinhas não é uma tarefa fácil devido às variabilidades ambiental e genética (3), sendo o mais indicado a avaliação de desempenho de um conjunto de raios de competição para a escolha daquele desejável.
Tendo em vista a predominância de pesquisas que definem índices e seus raios de competição em razão de análises de correlação e regressão com atributos biométricos (2), (4), (13), (15), (16), torna-se relevante a modelagem de tais índices em função de diferentes raios de competição. Maiores amplitudes de variação entre raios de competição são importantes para melhor delinear o efeito da competição entre árvores. É importante salientar a importância do uso de modelos sigmoidais, como o logístico, para a identificação precisa do raio de influência competitiva. Mediante exposto, o objetivo do presente trabalho foi avaliar índices de competição dependentes da distância e definir um raio de influência competitiva entre árvores em um fragmento de Mata Atlântica sob recuperação florestal.
Material e métodos
O trabalho foi conduzido em um fragmento de Mata Atlântica no município de São João Evangelista - MG, em área do Instituto Federal de Educação, Ciência e Tecnologia de Minas Gerais, nas coordenadas de 18º33’5,05” Sul e 42º45’50,40” de longitude Oeste (17). A região possui clima do tipo Cwa pela classificação do sistema internacional de Köppen, com verão chuvoso e inverno seco. As médias anuais de temperatura e precipitação são de 20,2º C (médias variando de 16,7ºC em julho a 22,8ºC em janeiro e fevereiro) e 1.377 mm (médias variando de 10 mm em julho e agosto a 308 mm em dezembro), respectivamente. Os dados climáticos foram oriundos de registros anuais de 1982 a 2012, disponibilizados gratuitamente por Climate-data.org
O fragmento de Mata Atlântica possui 2,53 ha e se encontra em fase de recuperação florestal desde novembro de 2013. O histórico de uso e ocupação do solo contempla sucessivos monocultivos de Coffea arabica L. por período de, aproximadamente, 10 anos. O tipo de solo predominante é o Latossolo Vermelho-Amarelo distrófico com o horizonte A proeminente, textura arenosa, relevo ondulado e a 690 m de altitude.
O inventário florestal foi realizado aos 80 meses após início da recuperação do fragmento de Mata Atlântica. Diante da assimetria de forma e tamanho reduzido do fragmento, optou-se pelo lançamento de uma parcela experimental retangular de 100 × 40 m (4.000 m2, equivalente a uma intensidade amostral de 15,81%) no centro e representativa da área de estudo, dividida em 10 setores quadrados de 20 × 20 m (400 m2). Todos os fustes dos indivíduos arbóreos com circunferência a 1,30 m de altura do solo (CAP, cm) igual ou superior a 10 cm foram mensurados e mapeados, tendo as coordenadas referenciadas em um plano cartesiano (X, Y), onde X e Y é a distância em metros da projeção dos pontos em eixos para o vértice de origem (0, 0). O CAP e a altura total foram obtidos com auxílio de fita métrica e vara telescópica, respectivamente.
O diâmetro à 1,30 m de altura do solo (DAP, cm) foi calculado pela relação entre (CAP) e o valor de π (3,141592654...). Adotou-se o diâmetro equivalente (DAPEQ) para as árvores com bifurcação, com uso da expressão Eq.1 em que DAP1 ... n é o diâmetro de cada fuste. A altura (H, m) foi obtida pelo método de superposição de ângulos iguais. Para a estimativa volumétrica (V, m3), foi empregada a equação fornecida pelo (17), para mata secundária eq. 2.
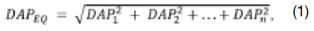
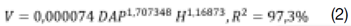
Para o cálculo de índices dependentes da distância, é necessário o estabelecimento de um raio de competição entre a árvore objetivo e suas competidoras. Avaliaram-se três índices de competição dependentes da distância Quadro 1) e vinte distâncias para o raio de competição (1, 2, 3... e 20 m); menores valores de índice expressaram menor competição sobre a árvore objetivo. O efeito de bordadura dos índices foi corrigido pelo método do deslocamento (Shifting), conforme recomendado por (9). Este procedimento consiste na emissão da cópia exata dos setores limítrofes da parcela experimental para a sua respectiva adjacência, cujas árvores são desconhecidas e estarão presentes fora da área amostrada.
Quadro 1 Indices de competicao dependentes da distancia avaliados em um fragmento de Mata Atlantica sob recuperacao florestal.
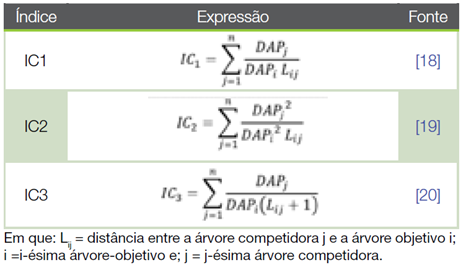
Os índices de competição foram submetidos às análises de correlação de Pearson e de regressão não linear através do método iterativo de Levenberg-Marquardt. O modelo logístico de três parâmetros foi ajustado para a estimativa do índice de competição em função do raio de competição, em metros eq. 3.
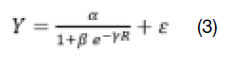
Em que: Y = índice de competição; R = raio de competição (m); α, β e γ = parâmetros do modelo logístico; e = constante neperiana; e ε = erro aleatório.
Para cada índice de competição, calcularam-se a distância correspondente a assíntota (m). A qualidade dos ajustes foi analisada pela significância dos parâmetros pelo teste t, Raiz Quadrada do Erro Médio (RQEM) e Média dos Desvios Absolutos (MDA). A análise gráfica consistiu na inspeção estatística de curvas com margem de confiança.
Calculou-se o coeficiente de variação (%) dos índices de competição em nível de parcela para cada um dos raios de competição em estudo. Posteriormente, estes dados foram submetidos ao teste de normalidade conforme Shapiro Wilk e análise de Boxplot. Para diagnóstico de efeito estatístico, foi empregado o nível de confiança de 95% de probabilidade em todas as análises. Estas foram efetuadas com auxílio do software R versão 3.5.2 (20).
Resultados
O inventário florestal contemplou 973 fustes ha-1 e 740 árvores ha-1, as quais apresentaram de 3,18 a 28,52 cm de DAP e altura total de 2,50 a 13,50 m. As correlações entre índices de competição e atributos biométricos foram discriminadas para distintos raios de competição, com distâncias variando de 1 a 20 m Quadro 2). Todas as correlações foram negativas, ou seja, os índices avaliados aumentaram com a diminuição dos valores de DAP e volume das árvores. A associação dos índices de competição e atributos biométricos se tornaram mais fortes à medida que aumentou o raio de competição, seguindo tendência logarítmica assintótica.
Quadro 2 Correlacoes entre atributos biometricos e indices de competicao dependentes da distancia para diferentes raios de competicao em um fragmento de Mata Atlantica sob recuperacao florestal.
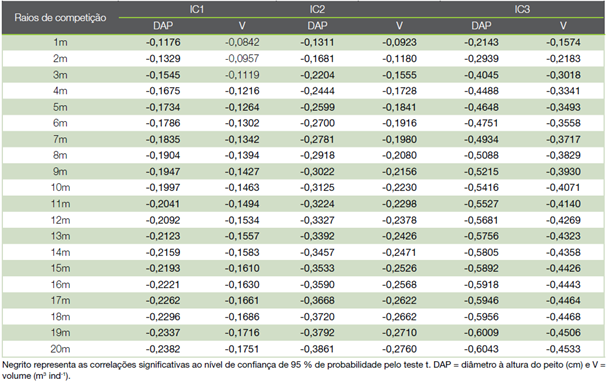
Notou-se que todos os índices de competição exibiram correlações significativas (p ≤ 0,05) a partir do raio de competição de 4 m, sendo mais fortes para o índice IC3. Este índice de competição foi o único que se correlacionou significativamente com a biometria das árvores mesmo diante de raios de competição reduzidos, como de 1 a 3 m de distância.
As relações funcionais estabelecidas para a estimativa de cada índice de competição em função do raio de competição se encontram no Quadro 3. A significância de parâmetros pelo teste t (p ≤ 0,05) e baixos valores de MDA e RQEM dos ajustes evidenciaram o comportamento sigmoidal, esperado para curvas provenientes do modelo logístico Figura 1). A assíntota das equações, representada pelo parâmetro α, se localizou entre as distâncias de 4,18 (IC1; α = 12,541423 m) a 5,18 m (IC2; α = 11,958259 m), para os índices IC1 e IC2, respectivamente.
Quadro 3 Coeficientes e qualidade do ajuste do modelo logistico para estimativa de indice de competicao dependente da distancia em funcao do raio de competicao para um fragmento de Mata Atlantica em recuperacao florestal.

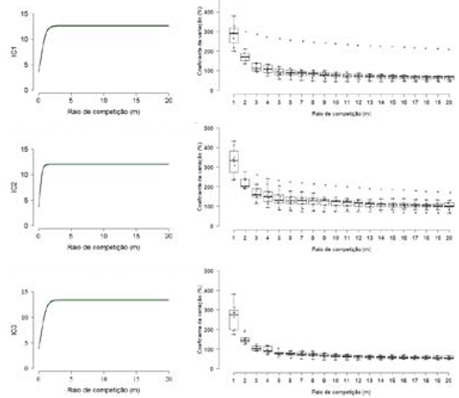
Figura 1 Representacoes graficas dos indices de competicao dependentes da distancia (a esquerda) e Boxplot do coeficiente de variacao (a direita) em funcao do raio de competicao para um fragmento de Mata Atlantica em recuperacao florestal. Margens de confianca (verde escuro) estabelecidas ao nivel de significancia de 5 % de probabilidade.
Figure 1. Graphic representations of distance-dependent competition indices (on the left) and Boxplot of the coefficient of variation (on the right) as a function of the competition radius for an Atlantic Forest fragment under forest recovery. Confidence interval (dark green) at the 5 % probability level of significance.
Em geral, a variabilidade dos índices de competição tendeu à estabilização a partir do raio de competição de 6 m Figura 1). O índice IC3 apresentou a menor variabilidade à medida que aumentou o raio de competição, concentrando valores mais próximos à medida de tendência central (mediana). Apenas os índices IC2 e IC3 exibiram normalidade para todos os 20 raios de competição pelo teste de Shapiro Wilk (p ≥ 0,05).
Discussão
A competição influenciou a biometria das árvores do fragmento florestal. A consistência biológica dos índices de competição foi demonstrada na própria expressão algébrica; o efeito da competição reduziu à medida que aumentou a distância entre árvores objetivo e a competidora, porém se intensificou com o aumento do diâmetro daquela vizinha competidora. Ressalta-se que a circunstância do valor de um índice ser maior ou menor de acordo com a competição segue princípios matemáticos, dependendo da expressão algébrica e relações diretas e inversamente proporcionais à competitividade (2). Obteve-se correlação negativa em todas as combinações entre índices de competição e atributos biométricos Quadro 2). Este fato está condizente biologicamente, pois os índices avaliados refletem a competição sobre a árvore objetivo (árvores maiores tendem apresentar menores índices de competição). Maiores taxas de crescimento são comumente relacionadas à disponibilidade de recursos, como o espaço, resultando em menor competição sobre a árvore objetivo (1), (4), (21).
Índices de competição podem se associar com atributos biométricos de modo distinto segundo a definição do raio de competição para árvores individuais Quadro 2). Nem todos os índices exibiram correlações significativas (p ≤ 0,05) para raios menores (de 1 a 3m), que consideram reduzidas zonas de influência competitiva. Tem-se a expectativa de que a zona de influência competitiva de uma árvore aumente com as suas dimensões e, consequentemente, qualidade de sítio, porte do dossel e estágio sucessional de formações florestais (21).
Pela análise de correlação, verificou-se que o índice IC3 (20) se associou significativamente (p ≤ 0,05) com o DAP e volume das árvores objetivo, mesmo adotando um raio de competição de apenas 1m Quadro 2).). Apenas a partir do raio de 4 m que todos os três índices de competição se correlacionaram com os atributos biométricos, sendo mais fraca para os índices IC1 e IC2. Enfatiza-se que a correlação se tornou mais forte nos maiores raios de competição, fato que pode favorecer a modelagem do crescimento e produção em nível de árvores individuais, contribuindo para a predição de produção em tempos futuros, partindo-se das condições atuais (8). Essa relação direta entre biometria vegetal e o raio de competição também foi observada para um remanescente de floresta ombrófila mista no sul do Brasil por (22).
Estabeleceram-se três relações funcionais para a estimativa da competição sobre a árvore objetivo em função do tamanho de raios de competição. A qualidade preditiva oscilou pouco entre as equações geradas, com coeficiente de variação de 3,79 e 3,66% para RQEM e MDA, respectivamente Quadro 3). Em razão da complexidade envolvida na representação da competição em florestas naturais (3), (4), (5), (6), (7), (15), (16), a qualidade dos ajustes realizados foi considerada satisfatória. O comportamento sigmoidal crescente das curvas obtidas com o ajuste do modelo logístico foi evidenciado na Figura 1. O raio de competição em que todos os índices de competição tenderam a estabilização foi de 5,18 m (IC2). Por questões práticas e facilidade de adoção do sistema métrico, sugere-se que o raio de competição não seja inferior a 6 m (equivalente a 28,27 m2), sobretudo, para fragmentos de Mata Atlântica em fase de recuperação florestal. Este resultado também corrobora com o proposto por (13) e (23), que estudaram raios de competição de 3 e 6 m, indicando o raio de 6 m para maiores correlações em um fragmento de floresta estacional semidecidual no sudeste do Brasil e em povoamentos mistos no leste do Canadá, respectivamente. Todavia, salienta-se que, apesar da tendência assintótica, raios de competição demasiadamente longos podem não representar biologicamente a real zona de influência competitiva da árvore objetivo.
Pela análise de Boxplot Figura 1), observou-se que a variabilidade dos índices tendeu à diminuição e estabilização com o aumento dos raios de competição. Visualmente, o ponto de estabilização da variabilidade também tendeu a ocorrer próximo ao raio de competição de 6 m. Nesse sentido, indica-se o planejamento de maiores raios de competição desde que estejam em conformidade com as dimensões da parcela a fim de se evitar amostragens externas à unidade amostral. Isto se deve ao empecilho de que índices baseados em raios de competição (dependentes ou semidependentes da distância) se tornam cada vez mais imprecisos para árvores próximas à borda da parcela, em virtude do desconhecimento daquelas competidoras além da parcela, sendo preconizado o emprego do método de deslocamento para correção de bordadura (9).
O índice IC3 se mostrou o mais adequado em relação aos demais (IC1 e IC2) para representar a competição na área de estudo, pois se correlacionou mais fortemente com os atributos biométricos, apresentou fundamentação biológica e menor variabilidade com o aumento do raio de competição. A adição de uma unidade ao componente da distância entre árvores competidora e objetivo, contida no índice IC3, tem o propósito de evitar a ocorrência de valores nulos no denominador da expressão algébrica proveniente de ocasiões de distâncias iguais à zero. Não foram encontradas distâncias iguais à zero entre árvores objetivo e a competidora na amostragem realizada no fragmento de Mata Atlântica sob recuperação florestal, indicando que tal adição de valor numérico 1 (um) pode, em alguns casos, favorecer a obtenção de dados normalmente distribuídos e a associação com variáveis biométricas. É conveniente ressaltar que a ausência de aditividade no denominador do índice IC3, representa exatamente a expressão de IC1.
Neste trabalho, a modelagem para estimativa de índice dependente da distância em função do raio de competição se mostrou eficaz para a definição da zona de influência competitiva de árvores contidas em um fragmento de Mata Atlântica sob recuperação florestal. Os resultados obtidos fornecem informações úteis para o desenvolvimento de futuras pesquisas sobre o efeito da competitividade vegetal, auxiliando predições do crescimento e produção em nível de árvore individual.
É pertinente destacar, ainda, o potencial uso da análise competitiva arbórea para a definição de estratégias silviculturais e manejo florestal sustentável. Uma abordagem promissora é o direcionamento da técnica de liberação para árvores objetivo de interesse comercial ou ambiental sujeitas à maior competitividade, eliminando-se a vegetação concorrente adjacente para promover o acúmulo de sua biomassa (24). Outra abordagem é o manejo de lianas, especialmente, aplicado a árvores em ambientes mais competitivos (25). Além disso, a implementação racional de técnicas de enriquecimento em ambientes menos competitivos é essencial para promover receitas e o equilíbrio do ecossistema (26).
Conclusões
O índice de competição dependente da distância IC3 é indicado para análises sobre relações entre a competitividade e aspectos biométricos no fragmento de Mata Atlântica sob recuperação florestal em estudo. O raio de competição de 6 m é indicado para estimação de índices de competição dependentes da distância na área de estudo. A modelagem para identificação de tendências assintóticas de índices dependentes da distância, utilizando raios de competição como variável preditora, auxilia a definição de zonas de influência competitiva.

