











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los bosques secos tropicales son uno de los ecosistemas tropicales más amenazados y degradados (1,2,3,4). Los procesos de restauración en estos bosques son complejos, en primer lugar, debido a una precipitación limitada (600 a 1 800 mm/año) y a una fuerte estación seca de al menos 3-6 meses donde cae poca o ninguna lluvia (5, 6). En segundo lugar, a pesar de que el reclutamiento inicial de plántulas vía semilla es bastante alto (7, 8), la tasa de mortalidad a menudo excede al reclutamiento (9), siendo en algunos casos extremadamente variable entre el 0 y 92,5 % (10). Finalmente, las plántulas que se utilizan para iniciativas de restauración o plantaciones comerciales son de mediana o baja calidad.
Para garantizar la sobrevivencia inicial de las plántulas en sitios de alto estrés ambiental, la clave podría ser producir plántulas de alta calidad. La calidad de planta se define como la capacidad que estas tienen para adaptarse y desarrollarse en las condiciones climáticas y edáficas del sitio donde se establecen (11), es la resultante de la calidad genética, morfológica, fisiológica y sanitaria (12), que permiten alcanzar un desarrollo (supervivencia y crecimiento) optimo en un sitio forestal particular y, por tanto, cumplir los objetivos establecidos en los planes de restauración (13).
La calidad de una planta se basa en medidas sencillas de caracteres morfológicos relacionados con su forma, estructura y distribución de la biomasa (12). Durante la etapa de vivero, existen diversas prácticas culturales que pueden modificar los parámetros morfológicos y fisiológicos de las plántulas, y por tanto cambiar sus estándares de calidad (14). Por ejemplo, el endurecimiento en la fase de vivero, pretende activar determinados mecanismos fisiológicos y morfológicos relacionados con la resistencia a factores de estrés, especialmente al estrés hídrico, térmico, nutricional y mecánico, en consecuencia, mejorar la supervivencia y el crecimiento tras el trasplante (15).
El endurecimiento o aumento de la resistencia a un determinado estrés se logra sometiendo a las plántulas a dosis sub-letales de ese mismo estrés (16). El endurecimiento por estrés hídrico pretende detener el crecimiento de las plantas y activar sus mecanismos de resistencia a condiciones de déficit hídrico para lograr un equilibrio entre la absorción y la perdida, especialmente evitando el estrés, tolerándolo o ambos (15). Esta práctica podría ser útil en los bosques secos tropicales, donde la disponibilidad de agua en el suelo, se considera el factor limitante más importante en el establecimiento, la supervivencia y el desarrollo de las plantas (17, 18).
Finalmente, la exigencia de plántulas de alta calidad y resistentes a determinados factores de estrés es mayor cuanto más limitante sean las condiciones del sitio donde se ejecutará plantación. En este sentido, una planta de buena calidad, debe tener un diámetro al cuello de raíz grande, bajo valor de esbeltez (cociente altura / diámetro al cuello de la raíz), un sistema radical fibroso y un valor alto del cociente biomasa de raíz / biomasa aérea (19). Por otro lado, el Índice de Calidad de Dickson (ICD) (20) es el mejor parámetro para indicar la calidad de plántulas, ya que expresa el equilibrio de la distribución de la masa y la robustez, lo que evita seleccionar plantas más altas pero débiles y descartar ejemplares de menor altura, pero con mayor vigor (19). Desde esta perspectiva, el objetivo de este trabajo fue determinar el efecto del riego y la fertilización en vivero sobre las características morfológicas de plántulas de especies con uso potencial para la restauración de áreas degradadas en el bosque seco Neotropical.
Materiales y métodos
Área de estudio y clima
La investigación se desarrolló en el invernadero de la Estación Experimental “El Limón” de la Facultad Regional Multidisciplinaria de Esteli, Nicaragua. Esta se encuentra ubicada a 1,5 km al suroeste de la ciudad de Esteli, entre las coordenadas 13° 03’ 44” N y 86° 21’ 57” O a una altura de 888 m.s.n.m. La región se caracteriza por su clima seco, con una temperatura que varía entre 16 y los 33 °C, con una precipitación aproximada de 830 mm, de los cuales el 90 % caen durante 6 meses de la estación húmeda comprendidos entre mayo y noviembre (21).
Colecta de semilla
Las semillas de Swietenia humilis Zucc, Cedrela odorata L. (ambas Meliaceae) y Ceiba Pentandra, C.F. Gaertn (Malvaceae) se recolectaron directamente de árboles en el campus de la FAREM-Esteli y en los alrededores de la Estación Experimental el Limón durante la estación seca del año 2017 (enero - marzo). Las semillas se secaron en un invernadero durante 20 días y se almacenaron en bolsas herméticas dentro de una nevera a 8°C aproximadamente durante 60 días (abril - mayo).
Germinación
En junio 2017, se llenaron bolsas de polietileno de 15,2 × 20,3 cm con un sustrato compuesto por una mezcla de tierra negra (suelo vertisol) y cascarilla de arroz en una relación de 3:1. En julio 2017 se inició la germinación de las semillas de S. humilis, C. odorata y C. pentandra, dos semillas de cada especie fueron depositadas por bolsa a una profundidad del doble de su tamaño. Las bolsas fueron humedecidas periódicamente hasta la germinación (15 - 25 días aproximadamente). Las plántulas fueron aclimatadas dentro de un invernadero durante 45 días (primera semana de septiembre 2017) y se mantuvieron dentro del invernadero hasta el final del experimento (25 de noviembre 2017).
Riego y fertilización
El tratamiento de riego y fertilización fue aplicado a las plántulas a los 45 días después de la siembra (segunda semana de septiembre 2017). Para definir la cantidad de agua a aplicar a cada plántula, se determinó la capacidad de campo del sustrato. Para ello, se escogió una muestra al azar de 10 bolsas llenas con el mismo sustrato que sería usado en el experimento. Cada bolsa fue saturada con agua y se dejó escurrir el exceso de agua durante 24 h. Posteriormente, se registró el peso inicial (g) de cada muestra y se colocaron en una estufa a 105 °C durante 24 h; pasado este tiempo, cada bolsa fue nuevamente pesada (peso final), y la diferencia entre el peso inicial y final se usó como la dosis de agua a aplicar a las plántulas, considerando que 1 g = 1 ml.
La cantidad de riego promedio estimada a aplicar a cada bolsa fue de 170 ml de agua. El riego fue aplicado individualmente a cada plántula, con frecuencia de 3 días para las que no tendrán estrés (+Riego) y cada siete días para las que tendrán estrés (-Riego) utilizando agua de la red interna de la Estación Experimental El Limón. Adicionalmente, a cada frecuencia de riego se le aplico dos niveles de fertilización (+Nutrientes y -Nutrientes) vía Drench (fertilizante disuelto en agua aplicado sobre la superficie del suelo) con una formula completa marca comercial FOLIAR PLUS NPK 15.8.6 con microelemento quelatados y vitamina B1, producido por ALBA MILAGRO International S.p.A. Parabiago (MI) ITALIA - Vía Corridoni 19 (www.albamilagro.com). Para el tratamiento de +Nutrientes, el fertilizante fue diluido a razón de 2,5 ml por litro de agua y se aplicó durante las tres primeras frecuencias de riego a partir de los 45 días después de la siembra.
Variables evaluadas
Las mediciones de los parámetros morfológicos para cada plántula se tomaron después de 75 días de haber aplicado los tratamientos de fertilización y riego. La altura en centímetros (cm) de las plántulas se midió desde la base hasta el meristemo apical del tallo más alto, utilizando una cinta métrica. El diámetro al cuello de la raíz se midió a 1 cm desde la base de la planta, utilizando un calibrador digital con un error de 0,01 mm. Para la evaluación del peso fresco y seco del sistema radicular y de la parte aérea, se separó el sustrato de la raíz en un recipiente con agua. Luego cada raíz fue lavada con agua hasta que quedo libre de partículas de suelo y se secó el exceso de agua con papel absorbente.
Posteriormente se separó el sistema radicular de la parte aérea y cada una de las partes en fresco fue pesada por separado en una balanza de ACCULAB VICON con precisión 0,001 g. La biomasa de la parte aérea y del sistema radicular de cada plántula fue colocada en una bolsa de papel y llevada a estufa a 70 °C por un periodo de 48 h y luego se pesaron cada uno de los componentes. Para determinar la calidad de las plántulas en vivero, se utilizó el ICD (20). Este índice integra la relación entre la masa seca total de la planta, la suma de la esbeltez y la relación parte seca aérea/parte seca radicular, el cual se presenta en la ecuación 1.

Diseño del experimento y análisis estadístico
El diseño experimental de este estudio es un Completamente Aleatorizado (DCA) con estructura factorial de tratamientos. Los doce tratamientos surgen de la combinación de dos intensidades de riego (+Riego = cada tres y -Riego = siete días) y dos de fertilización (N+ = con y N- = sin fertilizante) y las tres especies evaluadas. La unidad experimental consistió en una plántula sembrada en una bolsa plástica con 30 repeticiones por cada tratamiento (2 intensidades de riego × 2 dosis de fertilización × 3 especies × 30 repeticiones, n = 360).
Todos los análisis estadísticos y gráficos fueron ejecutados con InfoStat (22) mediante modelos lineales generales y mixtos; las especies, el riego, la fertilización, las interacciones dobles y triple se declararon como efectos fijos. Se probaron diferentes estructuras de varianza residual para considerar la falta de homogeneidad de varianzas (23). El supuesto de normalidad fue evaluado usando gráficos cuantil-cuantil y la prueba de Shapiro-Wilks. Los mejores modelos se seleccionaron de acuerdo con los criterios de información de Akaike (AIC) y Schwarz (BIC). En todos los análisis se reportan las medias ± 1 error estándar y las medias se compararon usando la prueba LSD Fisher (P < 0,05).
Resultados
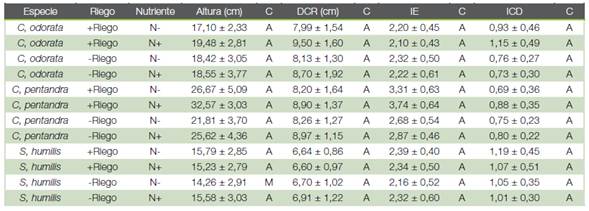
Los patrones de asignación de biomasa de las tres especies en respuesta a los tratamientos indican que son plántulas de alta calidad según la clasificación de Rueda-Sánchez, et al. (24), salvo en el caso S. humilis con -Riego y N- que mostro una calidad media (Cuadro 1 ). Para el índice de calidad de plántulas de Dickson (ICD) no se encontró interacción significativa entre especie, riego y fertilización (F2,347 =1,64; P > 0,05).
Cuadro 1 Efecto del riego y la fertilización sobre los principales parámetros morfológicos de calidad de plántulas forestales (media ± E.E), durante 75 días en vivero, n = 30 plántulas por especie.
El riego influyo significativamente sobre el índice de calidad de plántulas de Dickson (ICD) pero de distinta manera a una de las especies (especie × riego en ICD, F2,347 = 4,65; P < 0,05; figura 1). La especie S. humilis fue la de mayor ICD promedio; en el otro extremo C. pentandra fue la especie con menor ICD, pero estas dos especies no fueron afectadas por el nivel de riego, mientras que C. odorata redujo su ICD hasta en 0,3 unidades cuando el riego fue limitado (-Riego). La adición de nutrientes no afecto significativamente al índice de Dickson en ninguna de las especies (F2,347 = 1,28; P > 0,05).
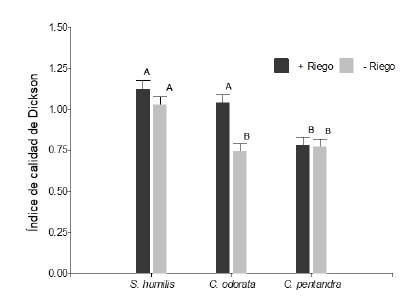
Figura 1 Índice de Calidad de Dickson para plántulas de Swietenia humilis, Cedrela odorata y Ceiba pentandra después de 75 días de aplicado del tratamiento riego (+Riego = 170 ml y -Riego = 170 ml cada 3 y 7 días respectivamente). Letras iguales indican medias iguales (P> 0.05).
Figure 1. Dickson’s Quality Index for Swietenia humilis, Cedrela odorata and Ceiba pentandra seedlings after 75 days of applying the irrigation treatment (+Irrigation = 170 ml and -Irrigation = 170 ml every 3 and 7 days respectively). Equal letters indicate equal means (P >0.05).
Para la fracción de masa de raíz (FMR en g/g) no se encontró interacción de entre los tres factores (F2,347 =0,80; P > 0,05). Para esta variable se encontró interacción significativa entre la dosis de fertilizante y las especies (F2,347 = 4,66; P < 0,05; figura 2). En promedio S. humilis y C. pentandra fueron las especies con mayor FMR, mientras C. odorata invirtió una menor cantidad de biomasa de raíz respecto al total de biomasa. No obstante S. humilis respondió al tratamiento de fertilización produciendo una fracción menor de raíces. De manera general, con riego limitado (-Riego) las especies presentaron una mayor FMR en comparación a cuando el riego no fue limitante (F1, 348 = 7,18; P < 0,01).
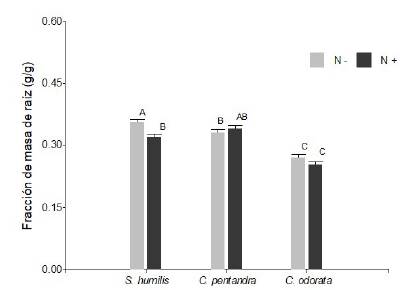
Figura 2 Fraccion de masa de raiz (g / g) para plantulas de Swietenia humilis, Ceiba pentandra y Cedrela odorata despues de 75 dias de aplicado del tratamiento de fertilizacion via Drench (N- = sin adicion de nutrientes y N+ = con adicion de nutrientes). Letras iguales indican medias iguales (P > 0.05).
Figure 2. Root mass fraction (g / g) for Swietenia humilis, Ceiba pentandra and Cedrela odorata seedlings after 75 days of applying the fertilization treatment via Drench (N- = without addition of nutrients and N+ = with addition of nutrients). Equal letters indicate equal means (P > 0.05).
No se encontró interacción entre los tres factores (F2,347 = 1,34; P > 0,05) para la relación entre la altura y el diámetro al cuello de la raíz expresada como índice de esbeltez (IE). Esta variable difirió entre las especies en función del nivel de riego (especie × riego en IE, F2, 347 =21,22; P < 0,0001; figura 3a) y también con la fertilización adicional (especie x nutrientes en IE, F2,347 = 4,53; P <0,05; figura 3b). En ambos casos, C. pentandra presento mayor índice de esbeltez con riego frecuente (3,52 ± 0,07) y nutrientes adicionales (3,31 ± 0,07), mientras que este índice no difirió entre S. humilis y C. odorata independientemente del nivel de riego y de fertilización.
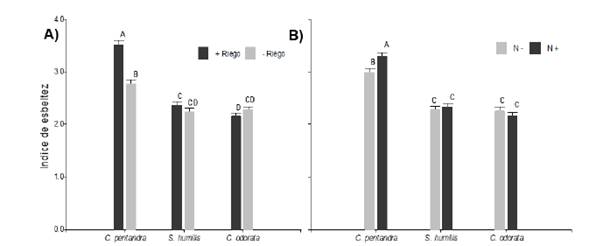
Figura 3 Índice de esbeltez en respuesta a los tratamientos experimentales (a) dependiendo de las frecuencias de riego (+Riego = 170 ml y -Riego = 170 ml cada 3 y 7 días respectivamente), (b) dependiendo de la disponibilidad de nutrientes vía Drench (N- = sin adición de nutrientes y N+ = con adición de nutrientes). Letras iguales indican medias iguales (P > 0.05).
Figure 3. Slenderness index in response to experimental treatments (a) depending on the irrigation frequencies (+Irrigation = 170 ml and -Irrigation = 170 ml every 3 and 7 days respectively), (b) depending on the availability of nutrients via Drench (N- = without addition of nutrients and N+ = with addition of nutrients). Equal letters indicate equal means (P > 0.05).
No se encontró interacción entre los tres factores (F2,347 = 1,64; P > 0,05) para la relación entre el diámetro al cuello de la raíz (DCR en mm). La aplicación de nutrientes adicionales provoco un incremento significativo en esta variable, pero de manera diferente en las tres especies (especies × nutrientes en DCR, F2,347 = 3,82; P < 0,05; figura 4). C. odorata y C. pentandra fueron las especies con mayor DCR y además sus plántulas respondieron positivamente a la adición de nutrientes. Concretamente, C. odorata fue la especie más sensible a la fertilización y su DCR aumento en 1,04 mm en promedio cuando se aplicaron nutrientes adicionales. No obstante, las plántulas de S. humilis alcanzaron un DCR siempre menor sin importar la adición de nutrientes. Por otra parte, el riego no tuvo ningún efecto sobre el DCR para ninguna especie (F1,347 = 0,03; P > 0,05).
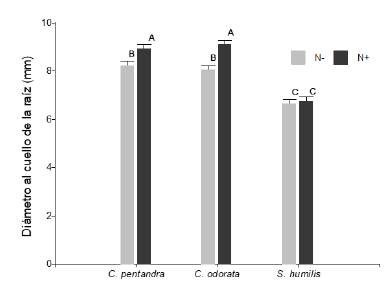
Figura 4 Diámetro al cuello de la raíz (mm) de las plántulas de Ceiba pentandra, Cedrela odorata y Swietenia humilis, en respuesta la disponibilidad de fertilización vía Drench (N- = sin nutrientes adicionales; N+ = con nutrientes adicionales). Letras iguales indican medias iguales (P > 0.05).
Figure 4. Root neck diameter (mm) of Ceiba pentandra, Cedrela odorata and Swietenia humilis seedlings, in response to the availability of fertilization via Drench (N- = without additional nutrients; N+ = with additional nutrients). Equal letters indicate equal means (P > 0.05).
Encontramos un efecto diferenciado de los nutrientes en la relación entre biomasa seca de raíz sobre la biomasa seca aérea (R:S) entre las especies (interacción especie × nutrientes en R:S, F2,345 = 3,68; P < 0,05; figura 5). De las tres especies evaluadas, C. odorata presento la menor R:S la cual mostro una cierta tendencia a la reducción cuando la disponibilidad de nutrientes fue mayor (N+ = 0,34 ± 0,02 y N- = 0,38 ± 0,02 respectivamente). Esta respuesta fue todavía más acusada en las plántulas de S. humilis, las cuales redujeron un promedio de 0,07 unidades su R:S cuando recibieron nutrientes adicionales. Por otra parte, C. pentandra, presento una R:S similar sin importar la disponibilidad de nutrientes.
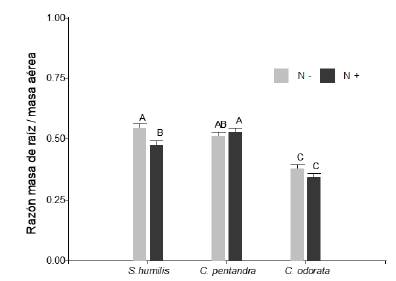
Figura 5 Relación de la biomasa seca de raíz respecto a la biomasa seca aérea (R:S) de plántulas de Swietenia humilis, Ceiba pentandra y Cedrela odorata, en respuesta la disponibilidad de fertilización vía Drench (N- = sin nutrientes adicionales; N+ = con nutrientes adicionales). Letras iguales indican medias iguales (P > 0.05).
Figure 5. Root to shoot ratio (R:S) of Swietenia humilis, Ceiba pentandra and Cedrela odorata seedlings, in response to the availability of fertilization via Drench (N- = without additional nutrients; N+ = with additional nutrients). Equal letters indicate equal means (P > 0.05).
El peso seco total (g) de las plántulas fue mayor cuando tuvieron nutrientes adicionales y riego constante (interacción especie × nutrientes × riego en PST, F2,347 = 3,37; P < 0,05; figura 6). Las plántulas de C. odorata y C. pentandra en condiciones de +Riego respondieron positivamente a la adición de nutrientes mientras que en -Riego el C. odorata no y la C. pentandra ligeramente. No obstante, en la S. humilis el patrón es algo distinto ya que la respuesta a los nutrientes fue relativamente escasa y en todo caso solo en condiciones de -Riego, por tanto, es en esas condiciones de riego donde las plántulas de esta especie (a diferencia de las otras dos especies) se benefician del fertilizante.
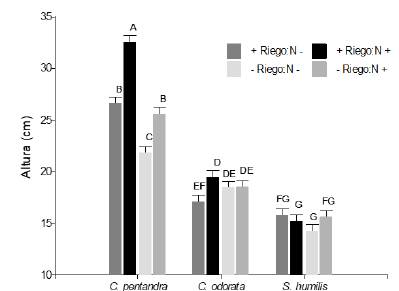
Figura 6 Altura de las plántulas (cm) de Ceiba pentandra, Cedrela odorata y Swietenia humilis en respuesta la disponibilidad de fertilización vía Drench (N- = sin nutrientes adicionales; N+ = con nutrientes adicionales) y el riego (+Riego = 170 ml y -Riego = 170 ml cada 3 y 7 días respectivamente). Letras iguales indican medias iguales (P > 0.05).
Figure 6. Height of the seedlings (cm) of Ceiba pentandra, Cedrela odorata and Swietenia humilis in response to the availability of fertilization via Drench (N- = without additional nutrients; N + = with additional nutrients) and irrigation (+ Irrigation = 170 ml and -Irrigation = 170 ml every 3 and 7 days respectively). Equal letters indicate equal means (P> 0.05).
Igual que para el PST, la altura de las plántulas (cm) presento interacción entre riego, nutrientes y especies (F2,347 = 3,57; P < 0,05; figura 7). La altura de las plántulas de S. humilis parece insensible a los tratamientos. En el caso de C. odorata parece que la fertilización aumento la altura de las plántulas, pero este efecto fue menos aparente en con -Riego. Mientras que las plántulas de C. pentandra fueron las más plásticas en cuanto a su altura se refiere. Cuando tienen nutrientes extra tienen una fuerte tendencia a aumentar su altura y de nuevo, este fuerte efecto es algo menor en condiciones de -Riego.
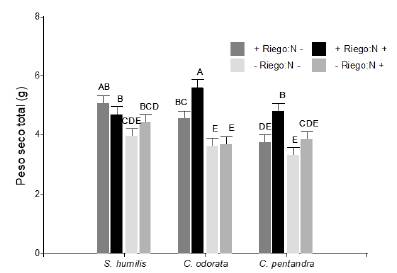
Figura 7 Peso seco total (g) de Swietenia humilis, Ceiba pentandra y Cedrela odorata, en respuesta la disponibilidad de fertilización vía Drench (N- = sin nutrientes adicionales; N+ = con nutrientes adicionales) y el riego (+Riego = 170 ml y -Riego = 170 ml cada 3 y 7 días respectivamente). Letras iguales indican medias iguales (P > 0.05).
Figure 7. Total dry weight (g) of Swietenia humilis, Ceiba pentandra and Cedrela odorata, in response to the availability of fertilization via Drench (N- = without additional nutrients; N+ = with additional nutrients) and irrigation (+Irrigation = 170 ml and -Irrigation = 170 ml every 3 and 7 days respectively). Equal letters indicate equal means (P > 0.05).
Discusión
El éxito de una plantación forestal comienza en el vivero, por lo tanto, la clave es producir plántulas de alta calidad, que cumplan con niveles óptimos de rendimiento (supervivencia y crecimiento) para contribuir a las metas establecidas en los planes de restauración (12, 13). Los datos de calidad de plántulas para los atributos morfológicos y funcionales ICD, IE, R:S, DCR, Altura, indican que las plántulas obtenidas son de alta calidad según Rueda-Sánchez, et al. (24). En general, valores pequeños de R:S, IE y altos de ICD indican una mayor probabilidad de supervivencia en condiciones de campo (20).
El ICD, es el indicador más importante que expresa el equilibrio de la distribución de biomasa y la robustez; mediante su aplicación, se evita seleccionar plantas desproporcionadas y descartar plantas de menor altura, pero con mayor vigor (20, 25). En este estudio, se encontró un efecto del riego sobre el ICD entre las especies, pero no de los nutrientes. A pesar de ello, las plántulas son de óptima calidad, ya que mostraron un ICD ≥ 0,5 (24). Las medidas de ICD en plántulas de C. odorata luego de 75 días, fueron similares a los reportados por Mateo-Sánchez, et al. (26), pero superiores a los reportados por Rueda-Sánchez, et al. (24), Basave Villalobos, et al. (27).
Así mismo, las plántulas de C. odorata presentaron un ICD superior a las reportadas por Villacis, et al. (28), pero fueron inferiores a los reportados por Gaburro, et al. (29). Se encontraron valores superiores de ICD para S. humilis a los reportados por Rueda-Sánchez, et al. (24), Basave Villalobos, et al. (30) para plántulas de 3 meses. Probablemente, las diferencias del ICD de las tres especies evaluadas en este estudio y los demás, se deba al tipo de sustrato, la forma e intensidad del riego y la aplicación de nutrientes adicionales.
En este estudio C. odorata presento valores inferiores de esbeltez respecto a los reportados por Basave Villalobos, et al. (27). De igual manera, los datos de C. odorata fueron inferiores a los datos reportados por Rueda-Sánchez, et al. (24) para plántulas de 3,5 meses e inferiores a los datos Mateo-Sánchez, et al. (26) para plántulas cultivadas en con diferentes sustratos. Para plántulas de S. humilis se encontró un IE similar a los reportados por Rueda-Sánchez, et al. (24), y a los reportados por Basave Villalobos, et al. (30). En ambos casos, la esbeltez de plántulas de C. odorata y S. humilis se mantuvieron dentro del intervalo optimo (< 6,0) sugerido por Rueda-Sánchez, et al. (24) para plántulas de óptima calidad. La esbeltez relaciona la resistencia de la planta con su capacidad fotosintética (31), por tanto, es preferible que los valores sean bajos, ya que esto indica que una planta es más robusta y con menos probabilidad de daño físico por la acción del viento y la sequía en los sitios de plantación (26, 32) como pueden ser los bosques secos tropicales.
Los datos indican una disminución de la FMR para S. humilis, C. odorata y en menor medida para C. pentandra cuando se aplicó riego frecuente y nutrientes adicionales. Estos datos son consistentes con el supuesto de que las plantas deberían disminuir el RMF en respuesta al aumento fertilidad del suelo, como una respuesta adaptativa a la limitación de recursos subterráneos (33, 34, 35, 36). En este sentido, las plántulas ajustan claramente su inversión relativa en el desarrollo de raíces en respuesta a las condiciones heterogéneas de crecimiento (37). Esta plasticidad en el patrón de crecimiento, expresada en forma de variabilidad intraespecifica en distintos atributos morfo-funcionales, puede ser clave para aumentar la probabilidad de establecimiento en escenarios de restauración con microclimas variables (37).
La relación de la raíz respecto al brote (R:S) es un indicador del potencial de supervivencia de las plántulas y ha sido concebido como una medida de equilibrio entre el área de transpiración (brote) y el área de absorción de agua y nutrientes (raíz) de una plántula (32). Los resultados indican un efecto de la fertilización, pero no de la sequía simulada sobre la relación R:S. En todos los casos, las plántulas se consideran dentro del rango optimo (<2,0) para plántulas de alta calidad (24). Sin embargo, la relación R:S, no debería exceder a 2,5, en especial cuando el factor mas influyente sobre la supervivencia del primer año es una larga y cálida estación seca o sitios con escasa precipitación (14, 32). Este es el caso de los bosques tropicales secos donde la precipitación anual es limitada, entre 600 a 1 800 mm, y además experimentan una fuerte estación seca de al menos 3-6 meses donde cae muy poca o ninguna lluvia (5, 6).
De las tres especies evaluadas, C. odorata fue la especie con menor R:S con y sin nutrientes adicionales, estos datos son similares a los encontrados por Basave Villalobos, et al. (27), pero superiores a las reportadas por Rueda-Sánchez, et al. (24). Los valores de R:S para S humilis corresponden a plántulas de alta calidad y son similares a los reportados por Rueda-Sánchez, et al. (24). Las plántulas de S. humilis y C. odorata presentaron un importante descenso de la R:S cuando no se aplicó fertilización. Es bien conocido que la R:S tiende a incrementar en respuesta a los tratamientos de sequía simulada (35, 38) y limitación de nutrientes (13, 33).
Esta respuesta, además es consistente con la hipótesis clásica del equilibrio funcional que predice que las plantas bajo un estrés dado maximizan su área de superficie para adquirir el recurso más limitante (39). Es decir, las plantas pueden adaptar su patrón de asignación de biomasa en función de las condiciones ambientales externas (33, 35, 40, 41).
En consecuencia, las plantas asignan más biomasa a las raíces cuando el agua y los nutrientes son los factores más limitantes y promueven la masa foliar y del tallo cuando falta luz o para compensar las pérdidas por defoliación debido a la herbívora (33, 35, 37, 41, 42, 43).
Así, por ejemplo, bajo condiciones de déficit hídrico ambiental o sequia controlada, la plasticidad morfológica permite reducir el diámetro de las raíces, incrementando la longitud y el área de superficie radicular por gramo de tejido invertido para maximizar la adquisición de agua y nutrientes del suelo (44, 45). De igual manera, Mokany, et al. (46) reportaron una mayor R:S cuando la precipitación era limitada (<1 000 mm/año).
Recientemente, Ledo, et al. (43) reportaron una variación global de la R:S ampliamente dominada por el tamaño del árbol y déficit hídrico climático. Mientras que, Norby, et al. (47) demostraron que la limitación progresiva del N limita el crecimiento de las plántulas.
Sin embargo, las plantas tienen la capacidad de asimilar más nutrientes al expandir su sistema radicular, mediante simbiosis con micorrizas o ambas (40). El endurecimiento en vivero de las plántulas para fines de restauración activa podría aumentar la supervivencia post trasplante debido a una mayor resistencia a algunos factores de estrés, especialmente hídrico, nutricional y mecánico.
El DCR, es probablemente el mejor y más fácil predictor global de crecimiento posterior y supervivencia de las plántulas en campo (32). El diámetro es un indicador del comportamiento de la altura y ambos definen la producción de biomasa en la parte aérea y la raíz (48). Este índice, relaciona la resistencia de la planta con la capacitad fotosintética de la misma (31). En este ensayo, el DCR de todas las plántulas, se mantuvo por encima del rango optimo (DCR ≥ 5,0 mm) para plántulas de alta calidad (24). Además, encontramos un efecto de los nutrientes sobre el DCR de todas las plántulas en vivero; en todos los casos, S. humilis fue la especie con menor DCR con y sin nutrientes, mientras que C. odorata y C. pentandra fueron las especies con mayor DCR.
Los resultados de DCR fueron inferiores a los reportados por Gaburro, et al. (29) para plántulas de C. odorata, pero son superiores a los reportados por Rueda-Sánchez, et al. (24) en plántulas de 128 días y a los reportados por Basave Villalobos, et al. (30) en diferentes sustratos. Para plántulas forestales con fines de restauración de bosque tropical seco, sería ideal llevar a campo plántulas con diámetros grandes, las cuales resisten mejor a la flexión, toleran más danos de insectos y animales, y están mejor aisladas del calor que aquellas con diámetros mas pequeños (49).
En relación a la altura, es conocido que las plántulas más altas pueden lidiar mejor con su vegetación competidora, aunque esto implica una buena salud fisiológica y un sistema radicular adecuado (50). Contrario a lo esperado, no se encontró efecto del riego, ni de la fertilización sobre la altura de las plántulas de C. odorata, S. humilis, pero si para C. pentandra. Sin embargo, en todos los casos, las alturas corresponden a plántulas de alta calidad, es decir, plántulas con altura ≥ 15 cm (24). Reportamos datos inferiores de altura en C. odorata a los presentados por Mateo-Sánchez, et al. (26) y a los reportados por Gaburro, et al. (29), pero similares a los presentados por Basave Villalobos, et al. (27). Los datos de S. humilis fueron superiores a los reportados por Rueda-Sánchez, et al. (24) para plántulas de 3 meses. La altura está asociada con la forma de crecimiento, la posición en un gradiente vertical de exposición a la luz, el vigor competitivo, el tamaño reproductivo, la fecundidad de la planta, la longevidad promedio y si la especie es capaz de establecerse y alcanzar un tamaño reproductivo (51).
Conclusiones
En vivero se verifico que el endurecimiento por estrés hídrico y nutricional mejora la calidad morfológica de plántulas forestales. El ICD difirió entre especies y por efecto del riego en especial para C. odorata. La FMR disminuyo en respuesta al aumento de los nutrientes, pero aumento con riego limitado. Por otro lado, la esbeltez difirió entre especies en función del riego y también de la fertilización, mientras que la altura y el PST de las plántulas fue mayor cuando tenían nutrientes adicionales y +riego. La medición de parámetros morfológicos relacionados con la sobrevivencia como la esbeltez, la FMR, R:S, DCR, altura, PST y en especial el ICD, permite mantener un control sobre el estado nutricional de las plántulas en vivero y ayudan a garantizar plántulas de óptima calidad para la fase de campo.

