











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La cubierta forestal de Brasil es de 463 millones de hectáreas, de las que el 98 % son ecosistemas naturales situados en el norte del país. El 2 % restante son plantaciones, principalmente de los géneros Pinus y Eucalyptus, de las que se obtiene el 81 % de la madera que se aprovecha en el país. La Mata Atlântica o bosque tropical atlántico es el cuarto ecosistema forestal por importancia, con 30,9 millones de hectáreas (10 % del territorio nacional) (1).
Hasta hace unos setenta años el bosque tropical atlántico se extendía por toda la costa brasileña pero actualmente solo se encuentran algunos fragmentos, sobre todo en áreas montañosas costeras de las regiones sur y sureste del país (2).
La gestión dirigida a conservar dichos recursos forestales debe basarse en un conocimiento suficiente de los mismos. Para lograr ese conocimiento es necesario descubrir la estructura del vuelo mediante técnicas de inventario forestal, siendo la distribución diamétrica una de las características que mejor describen dicha estructura (3).
El presente trabajo tuvo por objetivo evaluar el progreso de la regeneración natural del dosel de un área, colindante con una masa de bosque atlántico, en la Reserva Biológica Sítio dos Palmares, Estado de Espírito Santo (Brasil) como información de apoyo para fundamentar las correspondientes medidas de conservación.
Materiales y métodos
El área de estudio se localizó en la Reserva Biológica Sitio dos Palmares (Santa Maria de Jetibá: 20°02`27”S 40°44`45”O), específicamente en el distrito de Rio Claro del municipio de Santa Maria de Jetibá en el estado de Espírito Santo (Brasil), a una altitud de 692 m sobre el nivel del mar. De acuerdo con la clasificación de Köppen (4), el clima es de tipo tropical de altitud (Cwb) con estaciones lluviosa y seca bien definidas. La temperatura media anual es de 18 °C y la precipitación media anual de 1 800 mm.
La Reserva Biológica Sítio dos Palmares conserva algunos fragmentos bien preservados del bosque atlántico original. En la reserva, hay también algunas áreas en el entorno de dichos fragmentos, que durante muchos años estuvieron sometidas a una intensa actividad agrícola, específicamente con cultivos de café (5). Hace más de 33 años que este cultivo fue abandonado en la zona, iniciándose así un proceso de regeneración natural de la vegetación nativa. Para el presente estudio fue escogida aleatoriamente una de esas áreas en regeneración natural, que ocupaba aproximadamente seis hectáreas. En ella se establecieron seis parcelas de 400 m2 cada una (20 x 20 m), distribuidas aleatoriamente y separadas entre sí. El área de muestreo total fue de 2 400 m2.
Todos los árboles vivos, incluyendo palmeras, con diámetro normal igual o superior a 5 cm fueron identificados taxonómicamente en campo por un dendrólogo, y posteriormente se verificó la identificación botánica en el Herbario de la Universidad de Brasilia a partir de muestras recogidas en la zona de estudio. La nomenclatura de los ejemplares identificados siguió la clasificación w3Trópicos del Jardín Botánico de la Universidad de Missouri (6).
Se midió el diámetro normal de cada árbol identificado, calculándose la sección normal, el número de árboles por hectárea y el área basal (7). Para modelar el patrón de distribución de los diámetros normales de la población en su conjunto y por cada especie, se ajustó al número de individuos arbóreos por clase diamétrica los modelos matemáticos exponenciales de Meyer (8)(Eq 1)
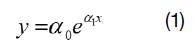
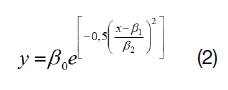
dónde: y es el estimador del número total de árboles en la clase diamétrica x, expresada por su marca de clase en centímetros ai, ßi, son los parámetros del modelo, calculados para la muestra completa y para cada una de las especies más abundantes.
Como estimadores de la bondad del ajuste se emplearon el coeficiente de determinación ajustado (R2), la raíz del error medio cuadrático (RMSE, en porcentaje) y el sesgo relativo (E, en porcentaje), ejecutados con el programa SigmaPlot (10).
A partir de las dotaciones reales de cada clase diamétrica se calculó la constante q de Liocourt como cociente entre el número de árboles por hectárea de una clase diamétrica y el número de árboles por hectárea de la clase inmediata superior (8).
El volumen maderable con corteza fue calculado para cada clase diamétrica aplicando un coeficiente mórfico de masa igual a 0,6 (7). Determinados la densidad poblacional y el volumen de madera por clase diamétrica se calculó para toda la masa el coeficiente de homogeneidad (11).
A partir de bibliografía se obtuvo para cada especie muestreada el síndrome de dispersión (DIS) (anemócoria, autocoria y zoócoria) y el grupo ecológico (GE) (especie pionera, secundaria inicial, secundaria tardía y clímax) como variables cualitativas; así como, la densidad de la madera como variable cuantitativa. A partir de dichos parámetros, las especies fueron reunidas en grupos funcionales (GF) según la metodología propuesta por Díaz y Cabido (12), construyéndose la matriz de distancia (13), utilizando el paquete ade4 en el programa R project. A partir de la matriz de distancia fue construido un dendrograma funcional (14) adoptando el nivel de corte de 0,6.
Resultados y discusión
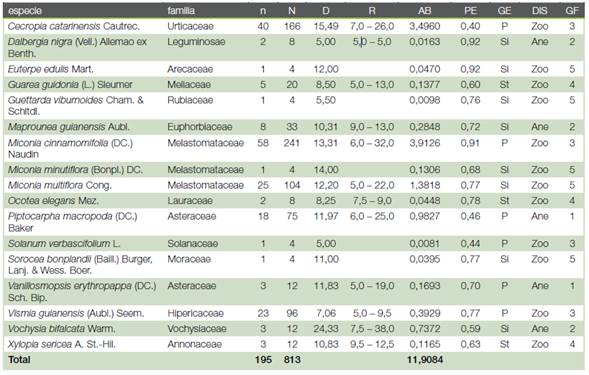
En el inventario florístico del área estudiada (2400 m2) se encontraron 195 árboles (813 árboles ha-1, 47.8 ± 95.6 árboles por hectárea), pertenecientes a 17 especies arbóreas, distribuidas en 15 familias (Cuadro 1 y Cuadro 2). La familia con mayor riqueza en especies fue Melastomataceae, con tres especies del género Miconia. Aunque se trata de una masa mixta pluriespecífica la mayor parte de las existencias recae en muy pocas especies. De hecho, las mayores densidades poblacionales se concentraron en las especies M. cinnamomifolia (242 árboles por hectárea; 14,2 ± 28,4 árboles por ha-1) y C. catarinensis (167 árboles por hectárea; 9,8 ± 19,6 árboles por ha-1), que reunían el 50 % del total de ejemplares de la población. Incluyendo M. multiflora (104 árboles por hectárea; 6,1 ± 12,2 árboles por ha-1) y V. guianensis (96 árboles por hectárea; 5,6 ± 11,3 árboles por ha-1) las cuatro especies acumulaban el 75 % de los ejemplares y el 78 % del área basal.
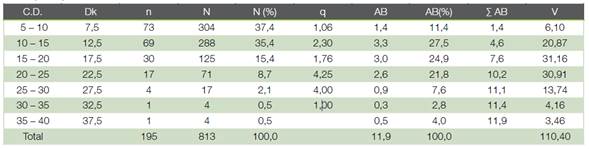
Cuadro 1 Distribución diamétrica de la población arbórea en la zona objeto de estudio (área en regeneración natural del bosque atlántico en la Reserva Biológica Sítio Palmares, Brasil). C.D.: clase diamétrica (cm); Dk: diámetro marca de clase (cm); n: número de individuos contabilizados; N: número de árboles por hectárea; N (%): porcentaje del número de árboles totales; q: constante de Liocourt; AB: área basal (m2 ha-1); AB (%): porcentaje del área basal total; Σ AB: área basal acumulada (m2 ha-1); V: volumen estimado (m3 ha-1).
Cuadro 2 Caracterización del arbolado en el área en regeneración natural de la Reserva Biológica Sítio Palmares, Brasil. n: número de árboles; N: número de árboles por hectárea; D: diámetro medio aritmético (cm); R: rango diamétrico (cm); AB: área basal (m2 ha-1); PE: densidad de la madera (g cm-3); GE: grupo ecológico; DIS: sistema de diseminación y GF: grupo funcional. P: pionera, Si: secundaria inicial, St: secundaria tardía, An: anemócora, Zo: zoócora.
A pesar de su pluriespecificidad, el área de estudio se halla en unos niveles de espesura y riqueza florística del arbolado claramente inferiores a los que refieren otros estudios en ámbitos ecológicos similares (Cuadro 3), lo que permite afirmar que el área de estudio se encuentra aún en una fase inicial de la estructuración hacia la fase clímax de desarrollo, puesto que no hay constancia de etapas de regresión durante los 33 años de regeneración natural de la zona de estudio.
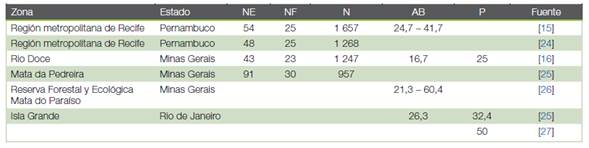
Cuadro 3 Estructura de la masa forestal en diversos estudios realizados sobre fragmentos de bosque atlántico brasileño. NE: número de especies; NF: número de familias; N: número de árboles por hectárea; AB: área basal (m2 ha-1); P: periodo de regeneración, en años.
De las 17 especies, tres de ellas tienen carácter de secundarias tardías (Cuadro 2). Incorporando a D. nigra (acumulan 50 árboles por hectárea entre las cuatro), podrán formar la base dominante de la estructura del futuro bosque secundario. Este grupo de cuatro especies presentan un diámetro medio de 8,5 cm (11,9/813 = 0,015 ± 0,03 cm) y se estima que su crecimiento diametral es de unos 4 mm año-1, que es el calculado para las especies pioneras del Cuadro 2. Por tanto, el grupo de especies secundarias tardías alcanzarían en unos 17-18 años un diámetro cuadrático medio de 15,5 cm, que es el estimado a partir de los resultados de Alves Júnior et al. (15) y Drumond y Meira Neto (16) para bosques más evolucionados.
Por lo tanto, 33 años de regeneración natural y crecimiento para la zona analizada parece ser un periodo aún insuficiente, como para que la dinámica de la masa alcance una estructura vegetal de comunidad arbolada pluriespecífica y disetánea con distribución diamétrica en J invertida, característica típica de los bosques primarios del bioma atlántico (15, 16). Es probable que, en ausencia de perturbaciones significativas durante las dos próximas décadas, se produzca un aumento en el número de especies; aunque, se cree que no será muy significativo debido a la falta de corredores naturales para la llegada de material reproductivo desde bosques próximos con mayor diversidad florística. De hecho, en el bosque primario adyacente al área en regeneración natural analizada fueron encontradas 70 especies distribuidas en 32 familias (17) y ninguna de las especies más abundantes aparecía en la zona de estudio. Por otra parte, se espera la densificación de la masa forestal por incorporación de nuevas cohortes anuales de regeneración, que lleven la distribución diamétrica de la masa al modelo decreciente en J invertida, como indicador de una posible estabilización de la estructura del vuelo.
Distribuidos los diámetros normales de la población en clases de 5 cm de amplitud Figura 1 y Cuadro 1 se constituyeron siete clases diamétricas, en un rango de 5 a 38 cm, con un diámetro cuadrático medio de 13.7 cm. Las dos primeras clases diamétricas, de 5 a 15 cm, concentraban el 73 % de la población y entre las tres primeras clases concentraban el 88 % del arbolado. La abundancia de las clases dimensionales inferiores, posiblemente clases de edad juveniles, deben orientar al gestor forestal hacia intervenciones de consolidación de dicha regeneración natural (2); así como, hacia acciones de restauración del tipo de plantaciones de enriquecimiento, tal como fue recomendado para algunos bosques secundarios establecidos naturalmente en potreros abandonados en el norte de Costa Rica (18).
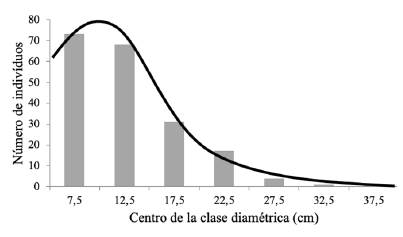
Figure 1. Stand diameter distribution and fitted Gauss curve.
El área basal (11,9 m2 ha-1; 0,01 ± 0,03 m2 ha-1) se hallaba concentrada en un 74 % en las clases diamétricas centrales (de 10 a 25 cm) (Cuadro 1). La distribución claramente asimétrica del número de árboles y la centralidad del área basal por clases diamétricas permitía suponer que la distribución normal o gaussiana con tendencia a la asimetría sería un modelo aceptable para describir la tendencia de la distribución objeto de estudio. El ajuste de la curva de distribución de Gauss mostró un coeficiente de determinación ajustado (R2) igual a 0,98, con un valor de la raíz del error medio cuadrático (RMSE) del 15,6 % y un sesgo relativo (E) del 2,0 %. En la Figura 1 se observa que solo las cuatro primeras clases diamétricas se ajustan aproximadamente al modelo y que éste sobreestima las dotaciones de las clases superiores, pudiéndose inferir que en las clases diamétricas superiores algunos árboles fueron probablemente cortados y las especies presentes (Cecropia catarinensis con 12 árboles por hectárea, M. cinnamomifolia con 8 árboles por hectárea y P. macropoda con 8 árboles por hectárea) constituyen el testimonio de las especies pioneras que aparecieron en la zona de estudio en un primer momento de la sucesión ecológica secundaria (Cuadro 4 y Cuadro 5).
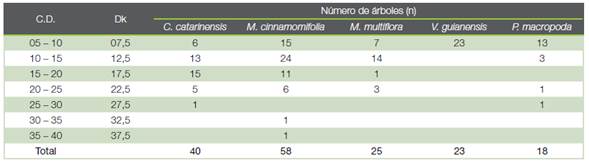
Cuadro 4 Número de individuos en cada clase diamétrica para las cinco especies arbóreas más abundantes. C.D.: clase diamétrica (cm); Dk: diámetro marca de clase (cm).
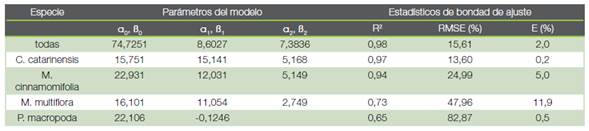
Cuadro 5. Parámetros de ajuste del modelo (1) para la población completa y para las tres especies más representadas (C. catarinensis, M. cinnamomifolia y M. multiflora), junto con el modelo (2) para P. macropoda. Se aportan los estadísticos de bondad de ajuste de las distribuciones.
Un valor del cociente q de Liocourt sensiblemente constante es un indicador de estabilidad de la distribución diamétrica (8). Se calculó para la masa forestal en estudio (Cuadro 1), comprobándose que en las clases extremas la q difería apreciablemente del resto. Esto permite conjeturar que un número apreciable de ejemplares de dichas clases fueron extraídos recientemente, quizá a causa de incendios de los que no se han encontrado evidencias. El área basal era muy diferente entre clases diamétricas (Cuadro 1), lo que hacía descartar que la estructura de la masa se hallara en situación de equilibrio según el criterio areal de Beranger (19).
En el análisis de la relación porcentual del número de individuos por clase diamétrica y correspondiente volumen de madera, se encontró un coeficiente de homogeneidad igual a 1,15, lo cual indica que la comunidad vegetal, de acuerdo con De Camino (11), se ajusta a la definición de bosque disetáneo irregular.
De las cinco especies que presentaban una abundancia superior a 75 árboles por hectárea, V. guianensis solo se encontraba en la primera clase diamétrica (5-10 cm). Para las otras cuatro especies: M. cinnamomifolia (242 árboles por hectárea), C. catarinensis (167 árboles por hectárea), M. multiflora (104 árboles por hectárea) y P. macropoda (75 árboles por hectárea) se ajustaron los modelos (1) y (2) a sus distribuciones diamétricas (Cuadros 2, 4 y 5; Figura 2). De las cuatro especies, C. catarinensis era la que más se aproximaba por su distribución a la ley normal en tanto que P. macropoda presentaba una distribución monótona decreciente que la hacía aproximarse a una curva en J invertida.
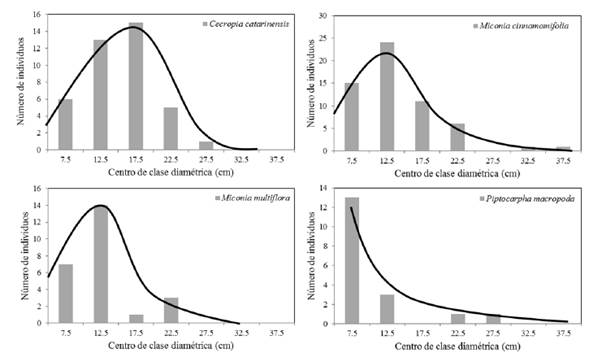
Figura 2 Distribución diamétrica de la población y modelos ajustados para las cuatro especies más abundantes de la muestra y con representación en dos o más clases diamétricas.
Figure 2. Stand diameter distribution and fitted models for the four most abundant species in the sample and represented in two or more diameter classes.
En función de los atributos cualitativos asignados a las especies y las características cuantitativas evaluadas en la población arbórea, a pesar de la presencia de pocas especies en la muestra, fue posible identificar cinco grupos funcionales en el conjunto de especies que componen la regeneración natural del bosque atlántico en la Reserva Biológica Sítio dos Palmares (Cuadro 1 y Figura 3). El grupo funcional 1 (GF1) quedó constituido por especies pioneras anemócoras. En el grupo GF2 quedaron las especies anemócoras y secundarias iniciales. El grupo GF3 quedó compuesto por especies pioneras zoócoras. En el grupo GF4 se establecieron especies zoócoras secundarias tardías con densidad de la madera entre 0.60 y 0.78 g cm-3. El grupo GF5 quedó formado por especies zoócoras y secundarias iniciales.
Dentro de cada grupo funcional se sitúan especies que poseen funciones semejantes dentro del ecosistema, corroborando las afirmaciones de Tilman et al. (20) y de Hooper y Vitousek (21). Teóricamente se puede deducir que las especies pertenecientes al mismo grupo funcional (GF) son competidoras por recursos similares, como se ha observado en lo referente al proceso de dispersión (12). Consecuentemente las especies de los demás grupos no son competidoras, pudiendo darse incluso un proceso de mutualismo que no dificulta el desarrollo de las demás especies (22). Sin embargo, podría estar produciéndose una competencia intergrupal para ciertos recursos más escasos en la estación, específicamente los relacionados con la riqueza de nutrientes del suelo (23).
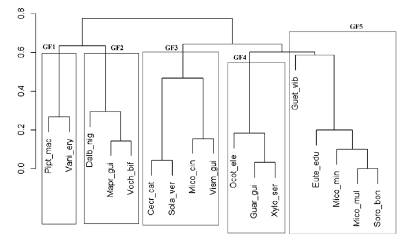
Figura 3 Dendrograma funcional con el agrupamiento de las 17 especies. El nombre científico está abreviado. Ejemplo: Pipt_mac = P. macropoda.
Figure 3. Functional dendrogram with the grouping of the 17 species. The scientific name is abbreviated. Example: Pipt_mac = P. macropoda.
Conclusiones y recomendaciones
A pesar de haber transcurrido 33 años en proceso de regeneración natural, el área de estudio aún se encontraba en una etapa inicial de la sucesión ecológica del arbolado, según atestiguaban la pobreza florística, la abundancia de regenerado de escaso tamaño, la baja espesura (tanto en número de árboles por hectárea como en área basal) y la tendencia a la normalidad de la distribución diamétrica de la población en su conjunto y de cada una de las especies arbóreas más abundantes. Se estima que tendrán que pasar todavía unos 17-18 años hasta que la estructura de la masa se asemeje a la de otros bosques atlánticos maduros. Se sugiere adoptar los criterios de manejo que aplica CATIE en los procesos de restauración forestal de bosques secundarios formados sobre potreros abandonados.
Se espera que el avance de la sucesión natural secundaria ya iniciada conduzca a una mayor dotación de las clases diamétricas superiores, con un mejor ajuste de la distribución diamétrica exponencial decreciente de Meyer. Es más incierta la posible transición de la masa forestal analizada hacia una mayor diversidad florística ya que las masas maduras de bosque atlántico más próximas no son colindantes y no hay corredores biológicos a través de los que pudieran trasladarse los propágulos de nuevas especies arbóreas. Por tanto, será muy aconsejable la realización de futuros inventarios para valorar si la evolución natural de estos enclaves de vegetación natural se dirige hacia estructuras semejantes al bosque atlántico originario.

