











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkLos animales terrestres se desplazan principalmente gracias al movimiento cíclico de las extremidades. Si bien el rastreo, que se basa en la contracción de la musculatura axial, es una técnica alternativa para la locomoción en tierra, la que implica los miembros es la forma más antigua de locomoción terrestre (Biancardi, Fabrica, Polero, Loss, y Minetti, 2011).
La locomoción con miembros implica diferentes desafíos de tipo mecánico: en cada ciclo, los segmentos tienen que ser reposicionados anatómicamente, con fases de aceleración, deceleración y la consiguiente elevación y descenso del centro de masa (Cavagna, 2017b). Este artículo busca aclarar conceptos a través de estudios clásicos, explicar diferentes parámetros que sirven para comprender la locomoción y discutir algunos estudios desarrollados con animales. Estos son importantes para entender las investigaciones posteriores. El objetivo de este trabajo es hacer una revisión de la literatura, que discute aspectos mecánicos y energéticos de la locomoción, el efecto del tamaño y los diferentes patrones, con sus modelos mecánicos.
Locomoción bípeda y cuadrúpeda
Los humanos desarrollan, en sus actos locomotores, tres actividades que les permiten movilizarse y que comparten con los otros bípedos y los cuadrúpedos: caminar, correr (o trotar) y galopar (skipping) (Saibene y Minetti, 2003). En los humanos, caminar se reconoce porque existe una fase de contacto de ambos pies en el suelo. Lo mismo pasa en los pares delantero y trasero de los cuadrúpedos caminando. Correr está determinado por la ausencia del doble apoyo y la presencia de una fase balística, o de vuelo, más o menos predominante. En los cuadrúpedos, el mismo movimiento, con un desfase del 50% entre los pares delantero y trasero, determina el trote. El galope o skipping se distingue por estar compuesto por un paso con doble contacto, seguido de un salto (fase de vuelo) y, en los cuadrúpedos, representa un patrón eficiente para moverse a gran velocidad. En los humanos, el galope se desarrolla en la niñez, o al bajar una colina. Incluso, es la forma de locomoción utilizada por los astronautas cuando existe menor aceleración de gravedad (Minetti, ; Pavei, Biancardi, y Minetti, 2015).
Los procesos mecánicos y metabólicos son diferentes entre caminar, galopar y correr, y todos provocan diferentes costos energéticos en la locomoción (Margaria, 1976; Margaria, Cerretelli, Aghemo, y Sassi, ). A diferencia de los cuadrúpedos, donde trotar es tan económico como galopar, para los humanos realizar el galope es más costoso que correr (Hoyt, Wickler, Dutto, Catterfeld, y Johnsen, 2006; Minetti, Ardigò, Reinach, y Saibene, Minetti, Pavei, y Biancardi, ; Pavei et al., ).
La comparación de los costos de funcionamiento entre los humanos y otras especies animales ha sido de gran interés para los científicos que buscan entender relaciones entre estructura y función de las extremidades (Roberts, Kram, Weyand, y Taylor, 1998). Taylor, Schmidt-Nielsen y Raab () estimaron que el costo energético de carrera en humanos (C run, J kgˉ¹ mˉ¹, (ver la sección ''Costo de transporte'')) es dos veces mayor que el valor predicho para un animal de la misma masa. Fedak y Seeherman (1979) y luego Taylor, Heglund y Maloiy () informaron que, en realidad, el costo sería cercano a 43% más que el predicho por factores alométricos, los cuales se definen como la relación entre las proporciones del cuerpo y el tamaño total de este (Ranisavljev, Ilic, Soldatovic y Stefanovic, 2014). La principal conclusión de estos estudios es que la carrera humana tiene un alto costo comparado al trote y galope de los cuadrúpedos.
Lo contrario pasa con la marcha: caminar es más económico en los humanos que en los cuadrúpedos de masa similar (Bramble y Lieberman, 2004). En general, se acepta que el costo de transporte en la marcha de los humanos es más bajo y el de la carrera es más alto que en los animales de masa similar (Aiello y Wells, 2002; Bramble y Lieberman, ; Carrier et al., 1984; Leonard y Robertson, 1997; Rodman y McHenry, 1980; Schmidt-Nielsen, 1975; Steudel- Numbers, ; Steudel, ; Taylor, 1994). Alexander (2003) derivó una ecuación predictiva del costo de transporte en relación a la masa corporal basada en los datos de Taylor et al. ().
La Figura 1 presenta el costo de transporte de varias especies en relación con la masa corporal y la ecuación predictiva de Alexander (2003).
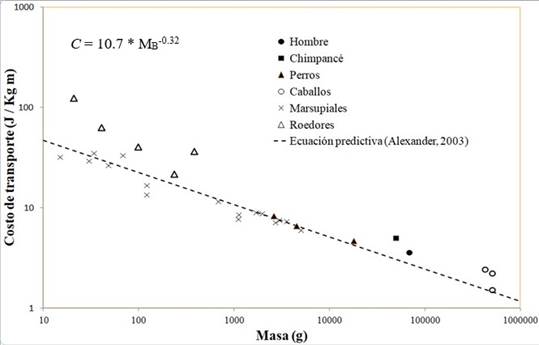
Figura 1 Costo de transporte (C) en relación con la masa corporal en algunos mamíferos terrestres y la ecuación predictiva de Alexander (2003), C = 10.7 * M -0.32 donde C = Costo de transporte (J/kg/m), MB = Masa corporal (kg). El C decrece al aumentar de la masa con una pendiente alrededor de -0.32 (los ejes se presentan en escala logarítmica). Fuente: elaboración propia a partir de (Minetti et al., 1999; Pontzer, Raichlen y Sockol 2009; Taylor et al., 1970; Taylor et al., 1982; Wunder y Morrison, 1974).
Desde el punto de vista histórico, el pionero en analizar aspectos biomecánicos humanos de la carrera y marcha fue Margaria (1938), quien estudió el efecto de diferentes velocidades e inclinaciones. En la década de 1940, los investigadores Wallace Fenn y Herbert Elftman hicieron muchas contribuciones, tales como el estudio de los cambios en energía cinética (E K ) y energía potencial (E P ), y métodos para calcular la fuerza en actividades de locomoción (Elftman, ; Fenn, ). El mismo Margaria, en la década de 1960, analizó aspectos como el costo energético usando un nomograma o nomógrafo, un instrumento gráfico de cálculo bidimensional que permitía calcular fácilmente el gasto de energía en función de la velocidad e inclinación (Margaria, Cerretelli, Aghemo, y Sassi, ).
Posteriormente, Cavagna, Saibene y Margaria (1963; ) volvieron a investigar las características mecánicas en la carrera y la marcha. Cavagna y Kaneko () mejoraron las estimaciones de trabajo mecánico de la carrera considerando la transferencia de energía cinética entre los segmentos para el cálculo del trabajo interno. Después, Minetti, Ardigò y Saibene (1994) concluyeron que existe un aumento en el trabajo mecánico total en la carrera con inclinación. No obstante, los valores obtenidos de trabajo mecánico para la carrera sobreestiman el trabajo muscular realizado debido a la dificultad de cuantificar la energía elástica durante ese movimiento (Saibene y Minetti, ).
Fue en los años 70 del siglo XX que Robert McNeill Alexander empezó a utilizar la fórmula del número de Froude (nFr) (ecuación 1) para comparar aspectos mecánicos y energéticos de la locomoción en especies diferentes o en sujetos de tamaño diferente dentro de la misma especie (Alexander, ).
nFr = v 2 /gL
Donde v es la velocidad, g es la aceleración de gravedad y L la longitud del miembro inferior.
El nFr es un número adimensional, parte de un modelo denominado similitud dinámica, el cual asume que dos cuerpos geométricamente semejantes, cuyos movimientos se basan en el intercambio entre E K y E P , se comportan de forma semejante dinámicamente y se mueven al mismo número de Froude (Alexander y Jayes, 1983). Posteriormente, Alberto Minetti aplicó el concepto de similitud dinámica en diferentes estudios de comparación de marcha en poblaciones con diferentes características (Minetti, Saibene et al., ) y también comparando la marcha a diferentes valores de aceleración gravitacional (Minetti, ; ). La fórmula de Froude determina un límite superior a la velocidad de marcha (caminar), equivalente a nFr = 1. Se supone que la velocidad espontánea de transición marcha-carrera (o marcha-trote en los cuadrúpedos) ocurre a nFr = .5; la velocidad adimensional que corresponde a la velocidad óptima de marcha es el nFr = .25 (Alexander, ).
Mecánica de los miembros
El esqueleto de los vertebrados funciona como un sistema de palancas articuladas, donde los músculos producen movimientos y transmiten fuerza al desarrollar momentos en las articulaciones (torque). Los músculos solo pueden generar fuerza de tracción, entonces la flexión y la extensión articulatorias son desarrolladas por conjuntos antagónicos de músculos (McMahon, 1984) o por el mismo grupo muscular trabajando de manera concéntrica y excéntrica. Por ejemplo, la flexión de rodilla después del contacto inicial del pie con el suelo durante la marcha o la carrera son controladas principalmente por el cuádriceps, actuando de manera excéntrica. En una articulación, el brazo de palanca (r), que es la distancia perpendicular desde el vector de la fuerza muscular al eje de rotación de la articulación, influye sobre el torque ejercido (M) por el músculo, como sobre la velocidad angular de la articulación (ω) y su rango de movimiento. Sin embargo, M depende de una relación directa de r con la fuerza del músculo (F) ecuación (2).
M = F r
Entonces, (r) varía conforme la articulación gira, ya que tanto el ángulo que forma la línea de acción de la fuerza (dirección entre el origen y la inserción), como el segmento corporal con respecto al cual se está midiendo el radio cambian al girar el segmento. Mientras, ω varía inversamente con r (ecuación 3), donde vs es la tasa de acortamiento de las fibras musculares.
ω = v s / r
En consecuencia, el torque que un músculo puede desarrollar varía inversamente con el rango de movimiento de la articulación y la velocidad del movimiento articular que puede producir. La mejor composición de fuerza y velocidad depende del nivel de la relación fuerza-velocidad del músculo mismo (Margaria, 1976; McMahon, 1984), como de la relación fuerza-velocidad resultante del mecanismo de palanca (Alexander, 2003; Biewener y Patek, 2018; McMahon, ).
Cuando el pie entra en contacto con el suelo, se puede medir la fuerza de reacción (G). El componente vertical Gz se opone a la aceleración de gravedad y provoca la aceleración en este eje. Por su parte, el componente antero-posterior y el medio-lateral (Gx y Gy) representan la fricción en el plano horizontal y permiten al humano o animal acelerar, desacelerar, maniobrar y equilibrarse; además, son las fuerzas que permiten el avance hacia el frente o hacia los lados (ver Figura 2).
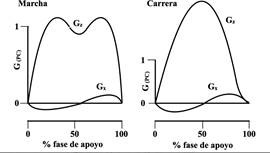
Figura 2 Fuerzas de reacción contra el piso en un paso de marcha y de carrera. G(PC): Fuerza expresada como fracción del peso corporal; Gz: Componente vertical de G; Gx: componente antero-posterior de G; en el eje horizontal el tiempo expresado como porcentaje del tiempo de apoyo. El área bajo las curvas representa el impulso. Fuente: elaboración propia.
En reposo, la fuerza Gz que actúa sobre cada miembro es aproximadamente igual al peso corporal dividido por el número de miembros que lo soportan. Cuando un animal se mueve, independientemente del tipo de paso y de la velocidad, las extremidades empujan al suelo, acelerando el cuerpo en la dirección deseada durante la fase llamada de apoyo. Luego, los miembros deben ser reposicionados para dar un paso adelante, dejando el suelo y acelerando también con respecto al centro de masa del cuerpo, en la fase llamada de oscilación.
Durante la locomoción, las fuerzas ejercidas por una extremidad en el suelo suben y bajan durante la fase de apoyo (cuando el pie está en contacto con el suelo) y es cero cuando la extremidad está en fase de oscilación; la fuerza vertical promedio ejercida por la suma de los miembros sobre un ciclo completo zancada tiene que ser igual al peso corporal para mantenerlo a una altura relativa del suelo. Se deduce que, si los miembros se mantienen en el suelo durante un período de tiempo más corto, se ejercerán fuerzas mayores sobre cada miembro (Alexander, 2003; Biewener, 1989).
La fracción relativa de una zancada cuando una extremidad particular está en la fase de apoyo se define como el Duty Factor de la extremidad (D F ) (Alexander, 1989b). Para moverse más rápido, un animal tiene que mover sus miembros más rápidamente, aumentando la frecuencia de la zancada; además, debe tomar pasos más largos, aumentando la longitud de la zancada, acoplando una o más fases de vuelo, cuando no hay miembros en fase de apoyo. En cualquier caso, el aumento de la velocidad de locomoción se logra reduciendo el D F y aumentando así la magnitud de la fuerza ejercida contra el suelo.
Los momentos que actúan en una articulación se tienen que equilibrar en condiciones estáticas o a velocidad angular constante, siendo F fuerza muscular, G fuerza de reacción, r el brazo de palanca y R la carga de la resistencia (ecuación 4).
F r = G R
Reordenando la ecuación (4):
r / R = G / F
Estas proporciones de brazos de palanca y fuerzas aportan una medida de la Ventaja Mecánica Efectiva (VME) de los músculos de las extremidades (Figura 2). La postura afecta la VME y, entonces, la alineación de los huesos articulados en la dirección de la fuerza de reacción del suelo cambia la ventaja mecánica, reduciendo la fuerza muscular empleada (Biewener, 1989; 1990); esto sucede cuando R disminuye. Tales cambios en la VME de las articulaciones de los miembros son importantes para permitir que mamíferos de diferentes tamaños mantengan el pico de tensión muscular y ósea dentro de un intervalo estrecho y, por tanto, tengan un factor de seguridad (Safety Factor) similar.
El factor de seguridad a menudo se define en términos de la resistencia de una estructura en relación con la tensión máxima probable que experimente durante su vida útil. El estrés óseo y muscular dependen del área transversal de los huesos y de las fibras musculares, que varían proporcionalmente a la masa corporal (Alexander, Jayes, Maloiy, y Wathuta, 1981). Por lo tanto, la reducción relativa a la masa en el pico de las fuerzas musculoesqueléticas se logra, en animales de gran tamaño, mediante la adopción de una postura más vertical, lo que cambia la VME (ver Figura 3) (Biewener, 1989).
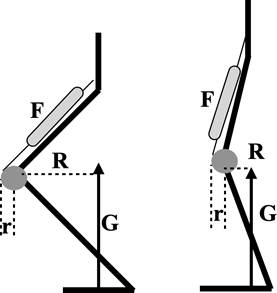
Figura 3 . Esquematización de un miembro inferior humano. G: Fuerza de reacción del suelo (ground reaction force); R: brazo de la fuerza G; F: fuerza del musculo cuádriceps; r: brazo de la fuerza F. La VME (r/R) a la rodilla resulta mayor en la postura más erguida (a la derecha). Fuente: elaboración propia.
Trabajo mecánico
Al final, los músculos convierten energía química producida por el metabolismo (energía metabólica, E Met ) en trabajo mecánico (W). En biomecánica de la locomoción se considera ''W TOT '' el trabajo mecánico total hecho para mover los segmentos corporales y desplazar todo el cuerpo. No se consideran los movimientos a nivel de tejido (fibras) muscular. Por supuesto, hay casos donde los músculos convierten E Met sin generar W TOT ; por ejemplo, en caso de contracciones isométricas o co-contracciones de músculos agonistas y antagonistas para estabilizar una articulación. Sin embargo, está generalmente aceptado que, durante la locomoción, el gasto energético E Met sea proporcional al trabajo mecánico W TOT (Cavagna, 2017b).
De acuerdo con Cavagna (2017a), ver la ecuación 5, donde W EXT es el trabajo externo, hecho para levantar y desplazar el centro de masa de todo el cuerpo (bCOM), y W INT es el trabajo interno, hecho para mover y reposicionar en cada ciclo los miembros.
WTOT = WEXT + WINT
El cálculo del trabajo mecánico se basa en el teorema trabajo-energía, el cual afirma que el trabajo mecánico hecho por un sistema en un dado proceso resulta ser la diferencia entre la energía mecánica total a final del proceso y la energía mecánica total (EMT) al inicio del proceso (ecuación 6).
W = EMT F - EMT I
Donde la EMT = E P (Energía potencial gravitacional) + E K (Energía cinética). EMT F = energía mecánica total final; EMT I = energía mecánica total inicial.
Si se suman los aumentos de EMT del b COM en el tiempo, se determina el trabajo externo positivo (W EXT ) (Cavagna y Kaneko, 1977; Cavagna et al., ; Willems, Cavagna y Heglund, 1995). El trabajo necesario para rotar y acelerar las extremidades respecto al b COM corresponde al W INT . El W INT constituye 25 a 40% del W TOT (Cavagna, ; Willems et al., ).
Dividiendo el W TOT por la E Met se obtiene la eficiencia mecánica de la locomoción (Eff). Dado que, como se explica en la siguiente sección, el gasto metabólico de la locomoción generalmente se expresa como costo de transporte (C; J kgˉ¹ m ˉ¹), por coherencia también el trabajo mecánico se divide por la masa del sujeto y por la distancia recorrida (Minetti et al., 1999).

Costo de transporte
El Costo de Transporte (C), es el costo metabólico necesario para mover una unidad de masa para una unidad de distancia (Schmidt-Nielsen, 1975). Fue estudiado en la marcha humana en 1963 (Margaria et al., 1963). El C (ecuación 7) se estima con medidas de calorimetría indirecta, es decir, con medidas de consumo de oxígeno y/o producción de dióxido de carbono. Generalmente, se prefiere convertir los mililitros de oxígeno a Joules, pues es la unidad de medida de la energía en el sistema internacional. Esto se realiza a través de un factor de conversión que depende del cociente respiratorio (mlCO2 / mlO2) (Di Prampero, 2015).
Donde C es el costo de transporte en J kgˉ¹ m ˉ¹; VO2 total (ml O2 kgˉ¹ s ˉ¹), es el consumo de oxígeno obtenido con un ergoespirómetro; VO2reposo (ml O2 kgˉ¹ s ˉ¹), es el consumo de oxígeno obtenido en reposo en bípedo durante 5 minutos; Velocidad (m s ˉ¹) es la velocidad media alcanzada y Eq(R) ( J mlˉ¹ O2ˉ¹) es el factor de conversión.
Se considera que, en sujetos adultos sin cambios del patrón de marcha, el mínimo C es cercano a los 2 J kgˉ¹ mˉ¹ y coincide con una velocidad de marcha óptima, muy cerca de la velocidad determinada por el ritmo de caminata natural (autoseleccionada) (Saibene y Minetti, 2003). El C aumenta cuando el valor de la velocidad es menor o mayor que el valor óptimo, determinando una característica curva con forma de ''U'', cuando se presenta el C en función de la velocidad (ver Figura 4).
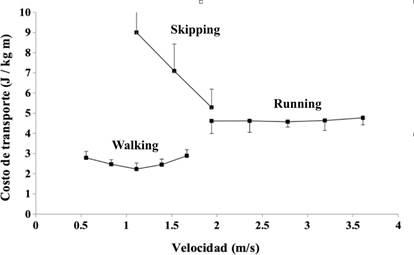
Figura 4 Costo de transporte (C) en relación del patrón y de la velocidad en el hombre. Fuente: elaboración propia basado en (Pequera et al., 2020).
En los adultos mayores, la curva del costo se desplaza hacia arriba, es decir, que el C aumenta con la velocidad (Mian, Thom, Ardigò, Narici, y Minetti, 2006). La velocidad autoseleccionada u óptima depende del tamaño, según la fórmula de Froude (ecuación 1), y coincide con un nFr = .25 (Saibene y Minetti, 2003).
Algunas condiciones o enfermedades pueden resultar en un alto C con alteraciones de su curva de distribución, con divergencias entre las velocidades de mínimo C y velocidad autoseleccionada (Bona et al., 2017; 2016; Bona, Gomeñuka, Storniolo, Bonezi, y Biancardi, ; Detrembleur, Dierick, Stoquart, Chantraine, y Lejeune, 2003; Detrembleur, Vanmarsenille,
De Cuyper, y Dierick, ). Estas condiciones cambian en ambientes de poca o nula aceleración de gravedad; por ejemplo, en la luna, donde se observa una reducción del trabajo mecánico y por lo tanto una disminución del C. Sin embargo, también la velocidad óptima y la velocidad límite de marcha bajan con la aceleración de gravedad, de acuerdo con la fórmula de Froude (). Por ejemplo, determinan la adopción de un patrón de galope en vez de marcha durante el desplazamiento de los astronautas en la luna (Minetti et al., 2012; Pavei et al., 2015). Esta es una consideración importante, puesto que en la actualidad existen terapias incipientes de rehabilitación de marcha basadas en condiciones de peso reducido o aceleración gravitacional reducida (Alkner y Bring, 2019; Salisbury, Baptista, Fei, Susin, y Russomano, 2015).
Como se ha afirmado antes, en condiciones terrestres el galope tiene un C muy alto, comparado con la marcha o la carrera (Minetti et al., 2012; Pavei et al., 2015) (Figura 4). Durante la carrera, el C es independiente de la velocidad (Pavei y Minetti, ) y en terreno plano a velocidad constante es del orden de 4 J kgˉ¹ m ˉ¹ (Di Prampero, Atchou, Brückner, y Moia, Lacour y Bourdin, 2015; Margaria, 1938; Pugliese et al., 2018) (Figura 4). Existen investigaciones que estiman que el C se podría obtener en base a ecuaciones que consideren la velocidad, la aceleración y la pendiente (McNeill Alexander, 2002; Minetti, Ardigò et al., ; Minetti y Pavei, ).
Patrones y modelos mecánicos
Si restamos los gastos ''de manutención'' del cuerpo, las actividades musculares y, por ende, la locomoción, consumen la mayor energía metabólica. Hay una fuerte presión selectiva hacia la evolución de mecanismos de optimización de los patrones de movimiento. En esta sección, se muestran los modelos mecánicos y los mecanismos minimizadores del gasto energético de los patrones más comunes de locomoción.
Marcha
La marcha se representa por un modelo de péndulo invertido, donde las extremidades inferiores son la estructura lineal y el bCOM es la masa unida a esta estructura (ver Figura 5) (Margaria, 1976; Saibene y Minetti, 2003). Durante la primera fase del paso, desde el apoyo del talón y hasta cuando la pierna de apoyo se encuentra vertical, el bCOM sube, aumenta su EP y disminuye su EK. Durante la segunda fase, al revés, ocurre una disminución de la EP y aumento de la EK (Cavagna, Thys y Zamboni, 1976).
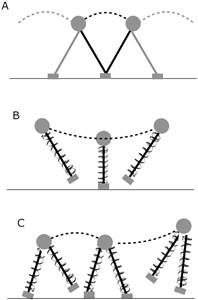
Figura 5 Modelos mecánicos de marcha (A: péndulo invertido), carrera y trote (B: masa-resorte), skipping y galope (C: péndulo y resorte). El círculo gris representa el centro de masa, y la línea discontinua su trayectoria. Fuente: elaboración propia.
Durante la marcha, la EP es alta cuando el bCOM está encima del punto de contacto del cuerpo con el suelo; pero, a partir de ese momento, EP comienza a disminuir y la EK aumenta gradualmente. Cuando el punto de contacto vuelve a estar en frente de bCOM, disminuye la EK y aumenta la EP. Las EK y EP se comportan en oposición de fase, generando un intercambio entre ellas, que permite reciclar energía mecánica y ahorrar energía metabólica (ver Figura 6) (Cavagna et al., 1976; Cavagna, Willems, Legramandi, y Heglund, ). El parámetro que mejor representa el modelo pendular es el porcentaje de recovery(R%) (recuperación o reconversión en su traducción), que cuantifica la capacidad de ahorrar energía mecánica a través de la reconversión entre las EK y EP (Cavagna, 2017c; Cavagna et al., 1976).
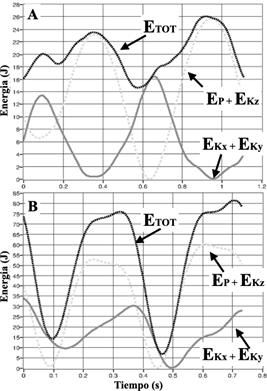
Figura 6 Ejemplo de trazados de energías del bCOM en un ciclo de marcha (A) y de carrera (B). EP = Energía potencial gravitacional; EKz = Energía cinética (componente vertical); EKx, EKy = Energía cinética (componentes horizontales); ETOT = Energía total. Fuente: elaboración propia.
En el modelo físico del péndulo ideal, la reconversión entre las energías es total (100%). En la marcha, el desfase debido al movimiento pendular puede llegar a un R% moderadamente elevado (hasta 60-70%) y dependiente de la velocidad de marcha (Cavagna et al., 1976). El máximo R% se encuentra alrededor de la velocidad óptima de marcha (que normalmente coincide con la velocidad autoseleccionada) y va bajando a velocidades mayores o menores. El máximo R% coincide con el mínimo C (Saibene y Minetti, 2003). Cuando es presentado en función de la velocidad de marcha, el R% presenta una característica de ''U'' invertida (ver Figura 7).
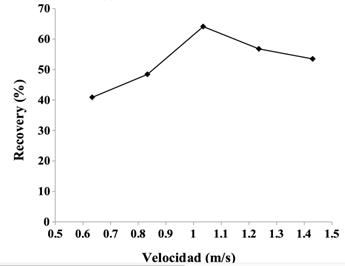
Figura 7 Porcentaje de recovery (recuperación) en función de la velocidad en un grupo de adultos, con su máximo a la velocidad de mínimo costo de transporte, alrededor de 1.1 ms-1 (Comparar con el C de la marcha presentado en la Figura 3). Fuente: elaboración propia con base en Bona et al. (2017).
Para describir el cálculo del R% (ecuación 8) se introducen dos nuevos parámetros: el trabajo vertical W V y el trabajo horizontal W H . El W V es la suma de los incrementos de EP + la componente vertical de EK. El W H es la suma de los incrementos de las componentes horizontales de EK.
R% = W H + W V - W EXT / W H + W V
El W EXT depende de las variaciones de energía total (EP + EK), que siempre son menores (o iguales) a la suma de las dos componentes (vertical y horizontal) tomadas por separado ( ).
Respecto a la contribución de la energía elástica (E E) ) Ishikawa, Komi, Grey, Lepola y Bruggemann (2005) explican la contribución a partir de la posición de músculos durante las fases de la marcha: el gastrocnemio medial estira durante la fase inicial de la fase de apoyo y luego permanece isométricamente durante la fase de apoyo final; el sóleo se alarga hasta el final de la fase de apoyo simple. Estos comportamientos sugieren que la E E se lleva a cabo no como un rebote (como ocurre en la carrera y saltos) en forma de resorte, sino como una acción de catapulta en el caminar. La interacción entre los fascículos musculares y los tejidos tendinosos desempeña un papel importante en el proceso de liberación de energía elástica. Lo anterior difiere de lo que se observa en la carrera, donde hay mayor participación de la E E
Carrera
La locomoción humana presenta mecanismos no completamente esclarecidos y las teorías buscan explicarlos. En la carrera, la teoría del sistema masa-resorte emerge como la más aplicable (Figura 5) (Blickhan, 1989; McMahon y Cheng, 1990). El modelo masa-resorte se fundamenta en él área de la física que estudia las condiciones de un oscilador armónico, el cual se basa en movimientos que se repiten en ciclos en torno de una posición de equilibrio. El comportamiento elástico de un resorte obedece a la Ley de Hooke (ecuación 9):
F = K x
Donde F es la magnitud de la fuerza elástica producida, K es la constante elástica del resorte (depende del tamaño y del material del resorte) (N.m-1); x es la variación de longitud del resorte (deformación). Cuando un resorte es alargado o comprimido, una cantidad de energía potencial elástica (E E ) puede ser almacenada y después liberada.
Blickhan (1989) y McMahon y Cheng (1990) fueron los primeros autores en proponer el modelo físico-matemático para carrera y saltos: una masa conectada a la extremidad de un resorte para simular patrones mecánicos de saltos y carrera y, así, cómo se minimiza la energía en estos movimientos. Ese modelo, masa-resorte, permite analizar la oscilación vertical del b COM . La oscilación del b COM describe una trayectoria similar a aquella de un sistema elástico denominado bouncing (rebote) (Cavagna, 2010). El sistema masa-resorte representa el modelo biomecánico de la energía presente preponderantemente durante esa actividad, caracterizado por un resorte capaz de almacenar energía elástica durante el contacto del pie con el suelo y liberarla durante el despegue (Blickhan, ).
Como se mencionó anteriormente, las energías asociadas al b COM son E K y E P . A través de la observación del comportamiento de estas energías se hace posible identificar cómo la dinámica del b COM minimiza el trabajo mecánico y maximiza la economía en la locomoción (Cavagna y Kaneko, 1977). Algunos parámetros del modelo masa-resorte son la rigidez vertical, frecuencia del sistema y asimetrías contacto-despegue; este modelo ha sido probado y ampliamente analizado, incluyendo diferentes situaciones y velocidades (Minetti, Ardigò et al., 1994).
Por su parte, en la marcha, los patrones rebotantes (bouncing gaits) como la carrera la E E juegan un papel importante en los mecanismos de ahorro de energía; su contribución se puede cuantificar de forma indirecta en humanos. Este hecho puede afectar el cálculo de la eficiencia mecánica de los patrones rebotantes (Minetti et al., 1999). En la carrera, la oscilación de la energía E P y E K es concomitantemente en fase (Figura 4). Es decir, no se observa una recuperación y conservación en el cambio entre las E P y E K .
Se ha postulado que, durante la primera mitad de la fase de apoyo de la carrera, principalmente, músculos y tendones se alargan almacenando energía elástica, mientras que los módulos de E P y E K disminuyen. Esa energía elástica, almacenada como un resorte, es sumada, aunque realmente es transformada en energía cinética potencial gravitacional, de modo gradual, durante la segunda mitad de la fase de apoyo (Minetti, Ardigò et al., 1994). Después de la fase de vuelo, parte de la energía total es nuevamente transformada en E E a través del alargamiento de la unidad músculo-tendón (Cavagna et al., 1964). Evidencias de E E almacenada y reconvertida también fueron encontradas en canguros y caballos (Alexander, 1989a). La característica anatómica del tendón permite ahorrar más o menos E E y, a diferencia de los canguros, las ratas canguros ahorran menos E E . Para obtener porcentajes de recuperación altos como los de los canguros, las ratas canguro necesitarían tendones extensores de tobillo mucho más delgados (Bienwener, Alexander, y Heglund, 1981).
El estudio en humanos es diferente al de animales. En estos últimos, a través de cirugía, fue posible implantar electrodos para medir fuerza y electromiografía, lo que posibilitó una medida directa (Biewener, 1998). Los estudios en humanos, normalmente, utilizan dinamometría, cinemática 2D y 3D, electromiografía, ultrasonografía, fibras ópticas, entre otros métodos (Alexander, 1984; Arndt, Komi, Brüggemann, y Lukkariniemi, 1998; Fina et al., 2000; Ishikawa et al., 2005).
Los ligamentos del arco del pie también responden, por el comportamiento elástico del sistema locomotor en la carrera. A pesar de la difícil estimación, evidencias indican que la E E es almacenada y liberada de los tejidos conectivos y contráctiles durante la carrera (Cavagna, 2010). Ker, Bennett, Bibby, Kester y Alexander (1987) muestran que, además de las estructuras elásticas (tendones y tejidos conectivos) que contribuyen en la inclusión de la energía elástica en la carrera, los arcos plantares también liberan energía elástica. Schleip y Müller (2013) estiman que para una persona de 70 kg a velocidad de 4.5 m s ˉ¹, el tendón de Aquiles almacena cerca de 35 J y los arcos plantares cerca de 17 J. Otros investigadores continúan recabando sobre la contribución de los tejidos conectivos en cuanto al ahorro de E E , como formas de mejora de la performance y cuidados preventivos para los tejidos conectivos y todo el sistema miofascial (Magnusson, Langberg y Kjaer, 2010; Shrier, 2004).
Como muestra la Figura 4, a diferencia de otros patrones de rebote, en la carrera no hay una velocidad de mínimo costo. Cuando el costo es evaluado en función de la distancia recorrida, el comportamiento es lineal (Saibene y Minetti, 2003), lo que parece haber favorecido la capacidad de correr largas distancias en la evolución humana, con individuos animados por cuestiones de sobrevivencia, fuga de depredadores, búsqueda de alimentos y un lugar seguro.
Este dato se asocia a la capacidad del eficiente mecanismo de absorción y liberación de energía elástica, con estructuras que permiten actuar como un resorte (modelo masa resorte), a diferencia de la marcha (poca contribución elástica, modelo pendular) (Bramble y Lieberman, 2004; Carrier et al., 1984; Di Prampero et al., 1986).
Aún no hay acuerdo sobre las determinantes mecánicas de la economía de la carrera en el ser humano. Parámetros espacio-temporales, velocidad, aceleración de gravedad y planos inclinados han sido ampliamente estudiados (Cavagna, 2010; Pavei et al., 2015). Si solo se toman parámetros biomecánicos internos, estos no consiguen explicar la economía de carrera (Kyrolainen, Belli, y Komi, 2001). Específicamente, la economía de carrera es definida como el consumo de oxígeno para una determinada velocidad submáxima de carrera y puede ser responsable de hasta 30% del rendimiento de un atleta de media y larga distancia (Kyröläinen, Belli, y Komi, ).
Así, para obtener un buen rendimiento en carreras de larga distancia, el atleta debe poseer alto consumo máximo de oxígeno (VO2MÁX), habilidad de mantenerse en un alto porcentaje de VO2MÁX y tener un pequeño gasto energético en determinada velocidad. Este último factor es evaluado a partir de la economía de carrera. Por lo tanto, el atleta más económico será aquel capaz de mantenerse en una velocidad submáxima con el menor gasto de energía metabólica. Varios estudios demuestran que el atleta es más económico cuando está en una longitud y frecuencia de pisada cerca de la autoseleccionada (Cavagna, 2010).
Así, el modelo masa-resorte es el mecanismo minimizador de energía de la carrera, auxiliando en la comprensión e interrelación con los beneficios de la práctica de ese deporte. Esos beneficios han sido documentados en el campo de la salud a través de la mejora en la capacidad cardiorrespiratoria, en la reducción de la presión arterial crónica, así como mejora del perfil lipídico. En cuanto al rendimiento deportivo, su entrenamiento implica mayor tolerancia a la fatiga, mayor consumo de oxígeno y mejora en la economía de movimiento (Capelli, Antonutto, Zamparo, Girardis y di Prampero, 1993; Costill, Branam, Eddy y Sparks, 1971).
Skipping (galope bípedo)
Es un término paralelo que asemeja al galope de los animales. Ha sido descrita bioenergética y mecánicamente por primera vez en humanos por Minetti (1998). Se presenta como una forma de andar observada con mayor frecuencia en niños (Roberton, 2013). El patrón de pasos se ejecuta con un paso y un salto en una pierna seguido por un paso y un salto en la pierna opuesta (similar a ir descendiendo por una pendiente muy inclinada). El patrón de skipping (galope) incorpora componentes de caminar y correr vista con una doble fase de apoyo, vuelo y apoyo de una sola extremidad, con un modelo mecánico que une elementos de la marcha y de la carrera (Figura 5) (Saibene y Minetti, 2003).
Una diferencia fundamental entre galopar y correr radica en la composición del paso; mientras correr repite un solo paso, idéntico, alternando entre las extremidades izquierda y derecha, galopar tiene dos pasos consecutivos, pero diferentes, realizados por cada miembro. El primer paso entra en contacto con la superficie antes del despegue de la otra extremidad, con esto se crea una breve fase de doble apoyo, seguida de salto del mismo pie. El aumento de la cadencia (número de pasos por minuto) muestra una mayor frecuencia de zancadas al comparar galope versus correr en todas las velocidades y gradientes (Minetti et al., 2012). El aumento de la cadencia y la disminución de la longitud del paso se debería directamente a las diferentes fuerzas de reacción del suelo entre los pasos (Heiderscheit, Chumanov, Michalski, Wille, y Ryan, 2011). Esta información es compatible con una disminución del pico vertical de fuerza de reacción que acompaña las longitudes de paso más pequeñas, observadas en el estudio de McDonnell y colaboradores en el 2017 (McDonnell, Willson, Zwetsloot, Houmard, y DeVita, ). Minetti et al. () informaron un costo metabólico invariablemente mayor al comparar con la carrera. Por esta razón, el galope podría ser un patrón de locomoción viable, ya que posee cargas musculoesqueléticas más bajas, con un costo energético más alto en comparación con correr (Minetti, ; Minetti et al., ).
En el contexto del deporte y salud, el galope, en comparación con la carrera, tiene picos de fuerzas de contacto de la articulación tibiofemoral y patelofemoral sustancialmente más bajas (30% y 98% respectivamente, según McDonnel et al., ), al igual que impulsos lineales por paso y por kilómetro (es decir, cargas acumulativas) más bajas, de cerca al 37% en la articulación patelofemoral; además, un costo metabólico un 30% más alto (McDonnell et al., 2019). Esto permite planificar ejercicios con menor carga articular y mayor gasto calórico. Si las fuerzas de contacto o los impulsos de la articulación de rodilla son factores causales de lesión, la sustitución de alguna cantidad de carrera con igual duración de galope puede permitir a los corredores reducir su potencial de lesión en la rodilla.
Conclusión
La aplicabilidad de todos esos conocimientos sobre la biomecánica de la marcha, carrera y galope tiene aplicación en distintas áreas del movimiento como educación física, fisioterapia o kinesiología, medicina, biología, física, ingeniería, entre otras áreas afines. Esto puede ser debido al hecho que actualmente diversas personas buscan el ejercicio físico como la caminata, carrera o galope bípedo con objetivos de mejor calidad de vida, ejercicio físico o rendimiento atlético (Capelli et al., 1993; Costill et al., 1971; Fries et al., 1994; Shipway y Holloway, 2010).

