











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO  uBio
uBio 
 Permalink
Permalink
Intro
El zorro gris pampeano (Lycalopex gymnocercus) es un cánido silvestre nativo de Sudamérica, que en Argentina presenta una gran abundancia y una amplia distribución (Luengos, 2003). Este carnívoro posee una dieta omnívora con un elevado consumo de frutos los cuales pueden llegar a constituir más del 50% de la misma (Nuñez & Bozzolo, 2006; Varela, Cormenzana-Méndez, Krapovickas & Bucher, 2008). En los bosques serranos de la provincia de Córdoba se ha observado que los zorros consumen tanto frutos de especies nativas como de especies exóticas entre los que se encuentran los frutos de mora (Morus nigra) (obs. pers.). Esta especie pertenece a la familia Moraceae, es nativa de Asia Menor y se distribuye en regiones tropicales y subtropicales de ambos hemisferios; en Argentina fue introducida con fines ornamentales llegando a convertirse en una planta invasora (Tolaba & Novara, 1996).
Morus nigra florece y fructifica en primavera (octubre y noviembre) y cada fruto posee entre 15 a 32 semillas en su interior de 2,5mm de largo por 1mm de ancho promedio (obs. per.). Al ser pequeñas, son fáciles de tragar por los animales frugívoros, lo cual puede resultar ventajoso a la hora de dispersarse (Galindo, 1998). Sin embargo, el hecho de que estos frutos sean consumidos por los zorros no implica necesariamente que sus semillas sean dispersadas legítimamente, es decir sanas y/o viables (Fleming & Sosa, 1994) debido a que el paso a través del tracto digestivo puede afectarlas negativamente, ya sea en forma mecánica y/o afectando su viabilidad.
El presente estudio tuvo por objetivos conocer si las semillas de M. nigra sufren daño al pasar a través del tracto digestivo de los zorros y verificar si dicho paso afecta la viabilidad, el poder germinativo, el inicio y tasa de germinación.
MATERIALES Y MÉTODOS
Para estudiar el efecto del paso de las semillas a través del tracto digestivo del zorro se comparó la germinación entre semillas recolectadas directamente de los frutos y semillas obtenidas de fecas del zorro. Para ello se recolectaron frutos de mora a orillas del Río Chocancharava en la zona noreste de la ciudad de Río Cuarto, provincia de Córdoba, Argentina.
El área de estudio posee una superficie de 12 hectáreas y la vegetación presente corresponde a la región fitogeográfica pampeana - distrito occidental (Cabrera, 1976). La temperatura media anual es de 18°C con una precipitación media anual de 800mm. En total se recolectaron 2000 frutos de M. nigra de 10 plantas diferentes.
Se seleccionaron al azar 1000 frutos a los cuales se les extrajeron las semillas. Posteriormente, se seleccionaron al azar 100 semillas a las cuales se les realizó el test de viabilidad de tetrazolio y 300 semillas las cuales fueron puestas a germinar en 10 placas de Petri de 30 semillas cada una. El resto de los frutos (1000) se utilizaron para alimentar a tres zorros en cautiverio del Parque Ecológico Urbano de la ciudad de Río Cuarto (PEU). Al día siguiente se recolectaron las fecas del recinto, y se llevaron al laboratorio donde fueron desarmadas cuidadosamente a mano.
Todas las semillas extraídas fueron analizadas bajo lupa estereoscópica para registrar posibles daños físicos (quiebres y/o exposición del embrión.). Se seleccionaron al azar 300 semillas extraídas de frutos y 300 semillas extraídas de fecas. En cada tratamiento, se sembraron 10 placas de Petri de 30 semillas cada una.
Las placas de Petri de las semillas de frutos y de las fecas del zorro se colocaron en germinador a temperatura constante de 25°C y se regaron diariamente. Dichas placas se revisaron cada dos días, durante 100 días, en búsqueda de evidencias de germinación. El criterio de germinación fue la emergencia de la radícula.
El inicio de la germinación (GS) y la tasa de germinación (GR) fueron evaluadas siguiendo el método de Izhaki & Ne’eman (1997). De acuerdo con estos autores el inicio de la germinación se define como el intervalo (días) entre la siembra y la germinación y se calculó como:
GS = 1/6 * P
donde P es el porcentaje final de germinación.
La tasa de germinación se calculó como:
GR = (5/6 * P) / (T2 - T1)
donde T1 es el intervalo de tiempo (días) entre la siembra y la germinación de 1/6 * P de las semillas, y T2 es el intervalo de tiempo (días) entre la siembra y la emergencia de 5/6 * P de plántulas.
El poder germinativo (PG) representa el porcentaje final de semillas que germinan (Sg) con relación al número total de semillas sembradas (Ss).
PG = (Sg/Ss) x 100%
Se calculó el porcentaje acumulado de germinación entre las semillas de los frutos y las fecas a lo largo del tiempo. Para los análisis estadísticos se utilizó la prueba de la t para muestras independientes para detectar diferencias significativas entre los tratamientos.
Por último, se aplicó la prueba de tetrazolio a las semillas que no germinaron al finalizar el experimento para conocer su viabilidad (Cottrell, 1947; MacKay, 1972). Para ello las semillas fueron cortadas por la mitad y se sumergieron en una solución acuosa al 0,1% de cloruro de 2, 3, 5 - trifenil - tetrazolio, pH 7, en la oscuridad durante 72 horas a una temperatura constante de 25°C. Finalmente, el embrión se observó bajo lupa estereoscópica; los teñidos de rojo eran viables y los no coloreados eran inviables (Bradbeer, 2013).
RESULTADOS
El 100% de las semillas recolectadas de las fecas de zorro estaban sanas; es decir no presentaron evidencias de daño físico luego de pasar por el tracto digestivo de los animales.
El 100% de las semillas provenientes de los frutos estaban vivas previo a comenzar el experimento.
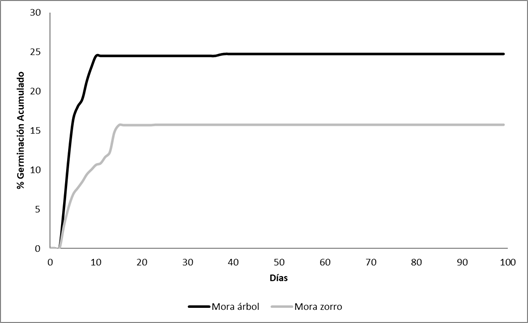
El inicio de germinación (GS) fue en el día tres tanto para las semillas provenientes de los frutos como para las obtenidas de las fecas de los zorros (t=-1,24; p=0,23; Fig. 1).
Los porcentajes de germinación acumulada de las semillas de los frutos y las obtenidas de las fecas de los zorros fueron significativamente diferentes (t=16,33; p<0,0001; Fig. 1). Esta diferencia entre los tratamientos se debería a diferencias en los porcentajes de germinación (PG) y a la tasa de germinación (GR). El PG fue significativamente mayor para las semillas de los frutos (PG= 82,5±3,41) que para las semillas de las fecas de zorro (PG=52,3±4,42) (t=5,42; p<0,0001; Fig. 1). La tasa de germinación (GR) fue mayor para las semillas de los frutos (13,75±4,27) que para las semillas de las fecas (3,96±3,85), mostrando diferencias significativas entre ellas (t=2,39; p=0,02).
Al finalizar el experimento 105 semillas obtenidas de los frutos (17,5%) y 286 semillas obtenidas de las fecas (47,7%) no habían germinado. A dichas semillas se les aplicó la prueba de tetrazolio para conocer su viabilidad y se encontró que el 47±9% y 21±4% de las semillas de los frutos y fecas respectivamente estaban vivas. Estos resultados muestran diferencias estadísticamente significativas (t=-2,68; p=0,02).
DISCUSIÓN
Las semillas de Morus nigra no sufren daño al pasar por el tracto digestivo del zorro, lo cual podría deberse a que las semillas son muy pequeñas en relación al tamaño de la boca de estos carnívoros, lo que permitiría que las semillas ingeridas sean generalmente defecadas sin daños mecánicos (Herrera, 1989; Galindo, 1998; Suárez‐Esteban, Delibes, & Fedriani, 2013). De acuerdo con estos resultados, el zorro sería un dispersor legítimo de M. nigra.
El inicio de la germinación fue similar para las semillas de los frutos que para las de las fecas de los zorros, ya que en ambos casos comenzaron a germinar poco tiempo después de ser sembradas. Resultados similares se han observados en otros carnívoros donde el 73% de las especies de frutos consumidos no manifiestan diferencias en el inicio de germinación (Lieberman & Lieberman, 1986; Traveset, 1998). El poder germinativo fue significativamente mayor cuando las semillas se extrajeron directamente de los frutos que las provenientes de las fecas. Lo mismo ocurrió con la tasa de germinación que, si bien en ambos casos sigue un patrón de crecimiento exponencial los primeros días y luego se detiene, es mayor para las semillas extraídas de los frutos que para las que fueron consumidas por los zorros.
Esto podría estar indicando que existe algún factor en el tracto digestivo de estos mamíferos que provoca una disminución en el número de semillas germinadas y una reducción en la velocidad a la que lo hacen. Dichos resultados, sumados al hecho de que las semillas de las fecas no presentaron daños externos y que el número de semillas viables al finalizar el estudio fue significativamente menor para las provenientes de las fecas, nos llevan a suponer que en el tracto digestivo de los zorros ocurre algún proceso químico que resulta negativo para las semillas de mora.
Diversos estudios han demostrado que el tamaño de una semilla puede determinar el tiempo que permanece en el tracto digestivo de un animal (Levey & Grajal, 1991; Izhaki & Arad, 1995), y que las semillas de menor tamaño tienden a estar más tiempo en el sistema digestivo, por lo cual es más probable que sufran daños por abrasión (Traveset, 1998). Estos resultados coinciden con el 17% de los estudios sobre el efecto de la endozoocoria, donde el paso por el tracto digestivo de diferentes animales perjudica la germinación de las semillas (Traveset, 1998). Mayores estudios son necesarios para conocer cuál o cuáles son las causas específicas de esta disminución en el poder germinativo y tasa de germinación.
M. nigra, como la mayoría de las plantas exóticas, genera una gran cantidad de frutos y cada uno de ellos puede tener hasta 32 semillas (obs. pers.), por ende, el consumo de una pequeña cantidad de frutos por parte del zorro implicaría que un número considerable de ellas estén siendo dispersadas sanas. Esto sumado a la amplia movilidad de este mamífero (Luengos, 2003) haría que las semillas consumidas puedan llegar a sitios alejados de la planta madre, lo cual le conferiría numerosos beneficios, ya que este transporte disminuye la competencia intra-específica (Galindo, 1998) y el riesgo de depredación (Janzen, 1970; Fleming & Heithaus, 1981; Fleming & Williams, 1990). La llegada de las semillas a áreas lejanas a la planta madre promovería, además, el flujo genético entre poblaciones de plantas (Heithaus, 1982). Es decir que, a pesar de que el zorro disminuye su poder germinativo y tasa de germinación, la interacción mutualista sería positiva para la planta ya que el zorro realizaría una importante lluvia de semillas y permitiría el ingreso de esta especie a nuevas áreas abiertas a la colonización. Teniendo en cuenta que la mora es una especie exótica invasora, serían necesarios estudios más profundos para conocer el efecto de su dispersión por endozoocoria sobre las especies nativas de la zona, las cuales probablemente se verán perjudicadas.

