











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLa relación mutualista más frecuente entre plantas y vertebrados es la dispersión de frutos y semillas, la cual involucra una gran variedad de mecanismos y de relaciones funcionales, ecológicas y evolutivas (Bascompte & Jordano, 2007). Entre los vertebrados, las aves juegan un rol fundamental en muchos ecosistemas ya que a través de mecanismos como la endozoocoria, pueden dispersar una gran variedad de frutos y semillas (Traveset et al., 2001; Figueroa & Castro, 2002; Amico & Aizen, 2005). Además, pueden tener un papel fundamental en la germinación y establecimiento de numerosas especies vegetales.
En los bosques tropicales la dispersión de frutos y semillas por vertebrados es superior al 90% y las aves se encuentran entre los principales dispersores (Frankie et al, 1974). Lo mismo ocurre en los bosques templados del hemisferio sur donde se ha observado que la dispersión de frutos y semillas por animales puede llegar a ser tan elevada como en las regiones tropicales (Gentry, 1982; Willson, 1991). Entre los bosques templados del hemisferio sur el bosque y matorral serrano de la provincia de Córdoba, Argentina, se caracterizan por presentar una importante diversidad florística y un gran número de especies de aves (Luti et al., 1979; Nores & Yzurieta, 1983; Nores et al, 1983; Nores, 1996;). Sin embargo, la dispersión de frutos y semillas por aves han sido escasamente estudiadas.
Recientemente se ha observado que un gran número de semillas están siendo dispersadas en los excrementos de aves y entre dichas semillas se encuentran las de Pyracantha atalantoides (Dellafiore, obs. per). Esta especie pertenece a la familia Rosaceae y comprende alrededor de diez especies de arbustos espinosos nativos del sur de China, Asia Menor y Europa (USDA, 2010). Numerosas especies de este género han sido introducidas en distintos países con fines ornamentales y en la mayoría de ellos se han naturalizado e incluso convertido en malezas invasora. Esta especie junto con P. angustifolia invaden zonas arbustivas y comparten dominancia con los arbustos nativos de frutos carnosos de los bosques abiertos y matorrales de las sierras de Córdoba (Tecco et al., 2006; Giorgis et al., 2011).
Los frutos de P. atalantoides son muy atractivos para las aves, las cuales han sido consideradas como responsables de su propagación en algunos ecosistemas naturales (Delucchi, 1991; Gurvich et al., 2005). Sin embargo, el hecho de que las aves dispersen sus semillas, no implica necesariamente que dicha dispersión sea efectiva, ya que el paso a través del tracto digestivo puede afectar negativamente su germinación. El presente estudio tuvo por objetivo conocer si las semillas de P. atalantoide sufren daño al pasar a través del tracto digestivo de las aves y si dicho paso afecta el poder germinativo, la velocidad o energía germinativa y el inicio y tasa de germinación.
Métodos
El área de estudio comprende una superficie de 3,5km2 en la ciudad de Alpa Corral, Provincia de Córdoba, Argentina y pertenece a la Región Fitogeográfica del Chaco Serrano (Cabrera, 1976); la cual se caracteriza por presentar una alta diversidad de aves (Nores, 1996; Miatello, 2003; Di Giacomo, et al., 2007) y en el área de estudio se censaron veintiséis especies nativas de América del Sur y al menos dieciséis de ellas consumen frutos y semillas de forma regular (Saltator aurantiirostris, Poospiza melanoleuca, Coryphospingus cucullatus, Zonotrichia capensis, Turdus rufiventris, Dellafiore obs. pers).
Entre las especies vegetales nativas registradas se encuentran Lithraea ternifolia, Fagara coco, Celtis ehrenbergiana, Schinus areira, Prosopis torquata y varias especies de la familia Poaceae como Stipa sp. y Festuca sp. Además, se encontraron especies exóticas como Ligustrum lucidum, Rubus ulmifolis, Pyracanthaatalantoides, Rosa eglanteria y Gleditsia triacanthos.
P. atalantoides florece y fructifica durante el verano y el otoño y en invierno los frutos suelen permanecer adheridos a sus ramas. Esta especie produce una gran cantidad de frutos y cada uno de ellos posee 5 semillas en su interior. Las semillas son alargadas, de color negro oscuro y brillante y poseen una longitud de 1,31 + 0,11 mm (media ± SE, n = 100) y un ancho de 0,87 + 0,09 mm (media ± SE, n = 100).
Para estudiar el efecto del paso de las semillas a través del tracto digestivo de las aves se hizo una comparación entre semillas recolectadas directamente de los frutos y puestas a germinar, con aquellas obtenidas de las excretas de las aves que también fueron puestas a germinar. En el primer caso se recolectaron 1 000 frutos de P. atalantoides de 20 plantas diferentes (50 frutos por planta). A dichos frutos se les extrajeron las semillas y se seleccionaron al azar nueve muestras de 100 semillas cada una. Cada muestra fue sembrada en placas germinadoras individuales. Para el segundo caso, se recolectaron muestras de excretas frescas de aves a lo largo de una transepto de 500m de largo por 5m de ancho. Debido a que las aves suelen defecar varios pellets en un mismo lugar se consideró como una muestra a todos aquellos pellets recogidos en la misma posición geográfica. En total se recolectaron 19 muestras las cuales fueron pesadas en el laboratorio y se desarmaron suavemente a mano. Posteriormente se observaron bajo lupa estereoscópica y las semillas de P. atalantoides fueron separadas. Dichas semillas fueron analizadas en detalle para registrar posibles daños físicos (semillas partida, tegumento sano o roto, exposición del embrión, deshidratación, etc.) y posteriormente las semillas sin daño aparente fueron sembradas para su germinación.
El tiempo transcurrido entre la recolección de las muestras y la siembra fue de un mes tanto en el caso de los frutos como de las excretas. Durante este período las muestras fueron conservadas en sobres de papel a temperatura ambiente. Las semillas recolectadas se colocaron a germinar en placas de plástico con algodón y papel secante. Las placas se regaron diariamente y se mantuvieron a temperatura constante de 20-25 ºC. El criterio de germinación fue la emergencia de la radícula. Se revisaron las semillas cada 2 días en búsqueda de evidencias de germinación durante 60 días.
Se calculó el porcentaje acumulado de germinación entre las semillas de los frutos y las excretas a lo largo del tiempo y se usó una prueba de Mann-Whitney para detectar diferencias entre los tratamientos.
El poder germinativo (PG) representa el porcentaje final de semillas que germinan (Sg) con relación al número total de semillas sembradas (Ss).
PG = (Sg/Ss) x 100%
El inicio de la germinación (GS) y la tasa de germinación (GR) fueron evaluadas siguiendo el método de Izhaki y Ne'eman (1997). De acuerdo con estos autores el inicio de la germinación se define como el intervalo (días) entre la siembra y la germinación y se calculó como:
GS = 1/6 * P
Donde P es el porcentaje final de germinación.
La tasa de germinación se calculó como:
GR = (5/6 * P) / (T2 - T1)
Donde T1 es el intervalo de tiempo (días) entre la siembra y la germinación de 1/6 * P de las semillas, y T2 es el intervalo de tiempo (días) entre la siembra y la emergencia de 5/6 * P de plántulas.
El tiempo medio de germinación (TMG) fue calculada como:
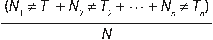
Donde Tn= número de días transcurridos desde el inicio de la germinación hasta el día n, Nn= número de semillas germinadas en el día n, y N número total de semillas germinadas.
Por último, se aplicó la prueba de tetrazolio a las semillas que no germinaron al finalizar el experimento (seis meses más tarde) para conocer su viabilidad (Cottrell, 1947; MacKay, 1972). Para ello las semillas fueron cortadas por la mitad y se sumergieron en una solución acuosa al 1 % de cloruro de 2, 3, 5 - trifenil - tetrazolio, pH 7, en la oscuridad durante 24 horas a una temperatura constante de 25 °C. Finalmente, el embrión se observó bajo lupa estereoscópica; los teñidos de rojo eran viables y los no coloreados eran inviables (Bradbeer, 1998).
Resultados
Se recolectaron 106 semillas de P. atalantoides de las 19 muestras de excretas de aves. Dichas muestras pesaron 41,7 g y se observaron 2,6 semillas por gramo; P. atalantoides estuvo presente en el 47,4 % de las muestras analizadas y representaron el 25,9 % del total de semillas contadas. Ninguna semilla de P. atalantoides mostró signos de daño físico o de deshidratación.
El poder germinativo (PG) fue similar entre las semillas provenientes de los frutos (50%) y las semillas de las excretas de las aves (46%). Sin embargo, fueron observadas diferencias significativas entre el porcentaje de germinación acumulado de las semillas de los frutos y de las excretas a través del tiempo (prueba de la U de Mann- Whitney, Z = 4,23, p < 0,001) (Figura 1). Estas diferencias se debieron a divergencias en el inicio (GS) y tiempo medio de germinación (TMG) y en la tasa de germinación (GR).
El inicio de la germinación (GS) de las semillas de los frutos fue de 5 + 1 día y continuaron germinando hasta el día 18. Al cabo de este período el 50 % de las semillas sembradas había germinado (Figura 1). El inicio de la germinación (GS) de las semillas provenientes de las fecas fue de 12 + 3 días y continuaron germinando hasta el día 26 cuando el 46 % de las semillas habían germinado (Figura 1).
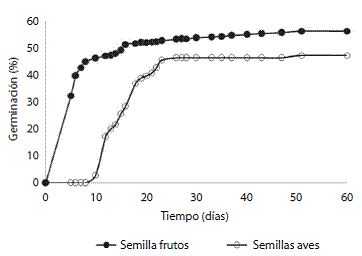
Figura 1: Tasa de germinación acumulada a través del tiempo de las semillas de Pyracanthaatalantoides.
La tasa de germinación fue menor para las semillas provenientes de las excretas (GR=5,2+1,3) que para semillas de los frutos (GR=21,0 + 4,3). Mientras que el tiempo medio de germinación fue menor para las semillas provenientes de los frutos (TMG=6,04+0,5) que para las semillas de las excretas (TMG=16,04+1,18).
La prueba de viabilidad de tetrazolio mostró que el 99,9 % y 100 % de semillas de los frutos y de las aves respectivamente eran inviables al cabo de seis meses.
Discusión
De acuerdo con nuestros resultados P. atalantoides está siendo legítimamente dispersada por las aves en el área de estudio, ya que las semillas estuvieron presentes en un alto porcentaje de las muestras, no sufrieron daños físicos al pasar por el tracto digestivo y el poder germinativo (PG) fue similar al de las semillas provenientes de los frutos. Sin embargo, las semillas que fueron consumidas por las aves registraron un retraso de siete días en el inicio de germinación (GS), además, la velocidad de germinación (TMG) y la tasa de germinación (GR) fueron 2,7 y 4,0 veces menor a la observada para las semillas de los frutos. Este retraso y reducción en la tasa de germinación ha sido observada en otras especies, como Rubus ulmifolius (Rosaceae), Rubia peregrina (Rubiaceae) y Phillyrea sp (Olaceae), pero es poco frecuente y aún se desconoce el mecanismo por lo cual ocurre (Traveset, 1998; Traveset et al., 2001; Traveset & Verdú 2002). Probablemente, los líquidos presentes en el sistema digestivo de las aves provoquen algún tipo de dormancia en las semillas. Sin embargo, aún no existen estudios que corroboren esta hipótesis.
Travestet (1998) sugiere que el paso de las semillas a través del tracto digestivo de los frugívoros afectaría principalmente a la dormancia funcional (ruptura de la latencia en las capas externas de la semilla) pero no a la dormancia fisiológica (dormancia del embrión). Debido a ello, independientemente de la época en la que las semillas son dispersadas, la germinación solo tendrá lugar cuando dichas semillas encuentren las condiciones del medio necesarias (luz, humedad, horas de frío, etc.). Una depreciación en el inicio y tasa de germinación causada por la ingesta de las aves, podría implicar una desventaja para P. atalantoides, ya que pueden encontrar una mayor competencia con las especies que germinan bajo las mismas condiciones y por lo tanto una menor probabilidad de supervivencia. Además, estarían sometidas a un mayor riesgo de depredación, por parte de roedores o insectos, y de ataque por plagas como los hongos (Janzen y Vásquez-Yanes, 1991). Numerosos autores han demostrado las ventajas de una germinación temprana en numerosas especies vegetales y en diferentes ambientes y como un retraso en el inicio de la germinación, aun de unos pocos días, puede afectar negativamente a la supervivencia de las plántulas (Ross & Harper, 1972; Halevy 1974; Weaver & Cavers 1979; Cook, 1980; Garwood, 1983; Zimmerman & Weis 1984; Garwood, 1986; Traveset et al., 2001). Por otro lado, si bien podría pensarse que las semillas en las excretas de las aves pueden verse beneficiadas, en cuanto a protección frente a depredadores y materia orgánica para la germinación, en general las excretas de las aves son pequeñas, se disgregan con facilidad y una simple lluvia o viento las puede dejar rápidamente expuestas. Mayores estudios serían necesarios para corroborar esta hipótesis.
En términos generales de costo - beneficio y teniendo en cuenta la alta movilidad de las aves, la endozoocoria beneficiaría a P. atalantoides ya que las semillas no son dañadas en el tracto digestivo, no se altera su poder germinativo, podrían ser dispersadas lejos de la planta madre y llegar a nuevas áreas abiertas a la colonización, lo cual compensaría el retraso en la germinación y los consecuentes riesgos a factores ambientales negativos.

