











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkLa depredación pre-dispersiva de semillas se refiere al 'ataque' o la muerte que puede experimentar una se milla en la planta madre o en el suelo junto a ella, an tes de ser dispersada (Vander Wall, Forget, Lambert & Hulme, 2005). La remoción a nivel pre-dispersivo por parte de roedores (Denslow & Moermond, 1982; Obeso, 1998; Englund, 1993; Santos & Telleria, 1994; Bas, Gómez & Pons, 2005; Briani & Guimaraes, 2007; Hernández, 2008) y aves (Traveset, 1993; 1994; Jordano, 1995; Bas et al., 2005) en especies de frutos carnosos y no carno sos (Perea, San Miguel & Gil, 2011b) ha sido relativamen te poco estudiada. En el caso de las áreas quemadas la situación es aún más desconocida ya que no hemos encontrado estudios que evalúen el impacto de los ver tebrados supervivientes o colonizadores sobre las diásporas resultantes a partir de la primera fructificación post-incendio. En Europa, la depredación de diásporas de frutos carnosos a nivel pre-dispersivo es realizada bá sicamente por aves de las familias Fringillidae y Paridae (Traveset 1993; 1994; Jordano, 1995) y por roedores de la especie Apodemus sylvaticus, que también podría re mover bellotas de Quercus coccifera, ya que esta planta rebrota abundantemente después del fuego (Gracia & Sabaté, 1996). Apodemus sylvaticus se considera uno de los principales removedores de frutos carnosos y bello tas en áreas templadas y mediterráneas (Hulme, 1997; Torre, Arrizabalaga & Díaz, 2002; Bas et al., 2005; Fedriani & Manzaneda, 2005; Pons & Pausas, 2007; Perea et al., 2011b) y aunque en la mayoría de los estudios no se eva lúa la remoción en la planta madre, existe evidencia de que los roedores son buenos escaladores (Obeso, 1998; Bas et al., 2005; Hernández, 2008) alcanzando alturas de hasta 3,75m en bosques de encinos ( Perea et al., 2011b). La actividad de remoción pre-dispersiva podría estar fuertemente condicionada por la presencia de cobertu ra y refugios (Fedriani & Manzaneda, 2005; Ouden et al., 2005) ya que se infiere un menor riesgo de depredación en ambientes con mayor cobertura debido a una expo sición menos evidente ante los depredadores ((Fedriani & Manzaneda, 2005).
El presente trabajo determina la remoción pre-dis-persiva de semillas por parte de vertebrados sobre tres especies de plantas, dos con frutos carnosos (Smilax aspera y Rubia peregrina) y otra con frutos no carnosos (Quercus coccifera) abundantes en áreas naturales de la Cuenca Mediterránea. Se espera una menor remoción de semillas en el área quemada en relación a un área no quemada (control) debido a la modificación del hábitat que afectaría las densidades y actividad de forrajeo de los roedores (Torre & Díaz, 2004) como principales ver tebrados depredadores de semillas en estos ambientes.
Materiales y métodos
El estudio se desarrolló en el área quemada de Ventalló (3°2' E, 42°8' N) y en un área no quemada cir cundante de la comarca del Alt Empordá, Cataluña, al noreste de la Península Ibérica (Figura 1). El clima de esta zona es de tipo mediterráneo, con un régimen térmico de inviernos moderados y veranos calurosos. Las tem peraturas mínima y máxima absolutas registradas son de -9 y 37°C, la temperatura promedio es de 15,2°C y la precipitación promedio es de 485,0mm anuales (Servei Metereológic de Catalunya, 2011). El estrato superior del bosque está dominado por el pino carrasco Pinus hale-pensis en el dosel y con encinas Quercus ilex dispersas en el subdosel. En el estrato inferior, predominan plantas de porte arbustivo como el romero (Rosmarinus officinalis) y las jaras (Cistus albidus y C. monspeliensis), trepadoras como la zarzaparrilla (Smilax aspera), y rastreras como la rubia brava (Rubia peregrina). Además, entre las plantas arbustivas destacan también el lentisco (Pistacia lentiscus) y la coscoja (Quercus coccifera), así como gramíneas (Brachypodium retusum y B. phoenicoides). La zona de estudio fue impactada por un incendio en el verano del 2006, el cual quemó 1 011ha de las cuales 613 corres pondían a pinares. Posterior al incendio realizaron una cosecha de salvamento ("salvage logging") o tala de los troncos de pino, provocándose una segunda perturba ción que afectó casi la totalidad del área quemada.
El experimento fue acorde con la fenología de fruc tificación de las especies de estudio, iniciándose en los otoños de los años 2008 y 2009, cuando los frutos y las semillas estaban completamente desarrollados (no in maduros). Se seleccionó una zona quemada y otra no quemada aledaña a la anterior. En ambas zonas se selec cionaron plantas individuales o parches de vegetación, según la especie, con una apreciable disponibilidad de frutos. Quercus coccifera, la principal especie produc tora de frutos no carnosos en el área, se distribuye am pliamente, de forma aislada o en parches de mediano a gran tamaño, especialmente en el área quemada. En esta misma área, Smilax aspera se encuentra predominante mente sobre montículos de rocas anteriores a la existen cia del bosque (probablemente vinculadas a actividades agrícolas) y/o en sitios en que la gestión de madera no provocó modificaciones de la cobertura arbustiva. En el área no quemada, las plantas de S. aspera fueron más co munes en áreas próximas a senderos que estaban más expuestos a la radiación solar en comparación a zonas internas del bosque en que el dosel era más cerrado. La selección de plantas de Q. coccifera ocurrió primordial-mente en parches de vegetación tanto en el área quema da como en la no quemada. Los parches con plantas de frutos carnosos seleccionados fueron más dispersos en el área quemada en comparación con el área no quema da. Por tanto, las distancias entre parches de vegetación y las superficies de los mismos fueron variables y condi cionadas a la cantidad de plantas con frutos disponibles en cada sitio. En Q. coccifera se observó que en el área quemada los rebrotes tenían un porte pequeño (<50cm de altura) a diferencia de la robustez de la mayoría de plantas encontradas en el área no quemada (con alturas cercanas o superiores a los 100cm de altura).
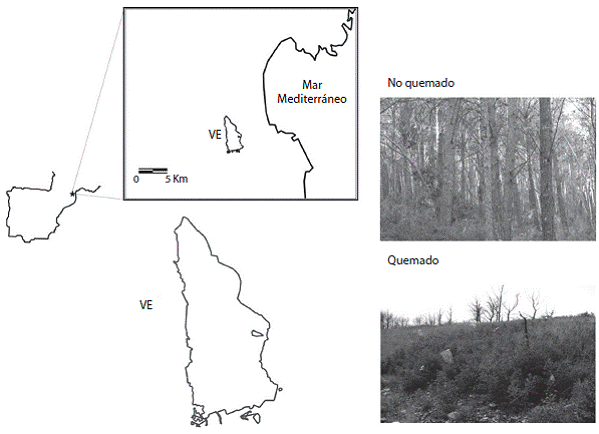
Figura 1: Localización geográfica y perímetro del área quemada de Ventalló (VE) en la cual se estableció el experimento de remoción pre-dispersiva de semillas. En las fotografías se puede constatar la diferencia en la cobertura vegetal de los hábitats quemado y no quemado circundante al perímetro.
Experimentos de remoción pre-dispersiva de se millas: En el muestreo del 2008 se seleccionaron 40 ra mas de Quercus coccifera, 12 racimos de frutos de Smilax aspera y 20 ramas de Rubia peregrina en el hábitat que mado mientras que 24 ramas de Q. coccifera y 8 racimos de S. aspera en el hábitat no quemado. En el muestreo del 2009 se evaluaron 30 ramas de Q. coccifera y 20 raci mos de S. aspera en el hábitat quemado y 20 racimos de Smilax aspera en el hábitat no quemado. En el año 2009 la escasa disponibilidad de bellotas en los bosques no quemados aledaños no permitió trabajar con Quercus coccifera. En el caso de Rubia peregrina se tuvo una si tuación similar por su escasa disponibilidad en ambos hábitats. En términos generales, la diferencia en el núme ro de racimos/ramas muestreados entre años y zonas de muestreo obedeció básicamente a la disponibilidad de frutos de ambas especies en ambos hábitats. Del total de ramas/racimos seleccionados para cada especie, la mitad se excluyeron de la remoción por vertebrados e insectos mediante la envoltura con malla fina de cedazo metáli ca con apertura cuadrada de 2mm de lado (ramas/raci mos excluidos). La otra mitad se dejaron descubiertos (ramas/racimos no excluidos); todos ellos marcados con cinta para su posterior localización (Figura 2). A las ramas/ racimos seleccionadas se les contó la cantidad inicial de frutos verdes y maduros que contenían (producción ini cial). Posteriormente, se realizaron conteos semanales o quincenales sobre las ramas excluidas y no excluidas, en función de su velocidad de maduración, tasa de remo ción o caída, hasta que la cantidad de frutos adheridos a las ramas se aproximara a cero.

Figura 2: Tratamientos de exclusión (izquierda) y no exclusión (derecha) en el experimento de remoción pre-dispersiva en los que se aprecian las tres especies de estudio: Smilax aspera (A) Quercus coccifera (B) y Rubia peregrina (C).
Estimación de la producción de frutos: Para contar con un indicador de la producción de frutos en ambos hábitats se realizó una estimación en las áreas en que se seleccionaron las ramas/racimos. Se realizó un conteo de los frutos carnosos y no carnosos adheridos a las plantas en el punto máximo de fructificación (considerado éste como el momento en que la mayoría de plantas tenían sus frutos desarrollados). Para cada planta con frutos se identificó la especie y se cuantificó el número de frutos adheridos. Dicha estimación se realizó una única vez al año ya avanzado el otoño para cada hábitat median te conteos en los parches de vegetación seleccionados (estimador de la producción del parche) y en sus alrede dores (estimador de la producción del hábitat). La pro ducción del parche se estimó mediante un transecto de 5m de longitud y un metro de anchura, colocado sobre la longitud mayor del parche de vegetación. Se contó la cantidad de frutos de todas las especies de plantas presentes a lo largo y ancho del transecto. En total, se muestreo un área de 5m2 por parche. Por otro lado, la producción de frutos del hábitat se estimó mediante un transecto de 20m, situado a 10m de distancia del parche y orientado de forma aleatoria aunque perpendicular al parche de vegetación. Para el conteo se utilizaron cinco áreas de 1m2 distanciadas entre sí por dos metros sobre la línea imaginaria del transecto. En total se muestreó un área de 5m2 por transecto (ver Figura 3). En el año 2008, se seleccionaron cuatro parches de vegetación en cada hábitat (dos de Q. coccifera, y dos de S. aspera) mientras que en el 2009 se seleccionaron cinco en el área quema da (tres de S. aspera y dos de Q. coccifera) y cinco de S. aspera en el área no quemada.
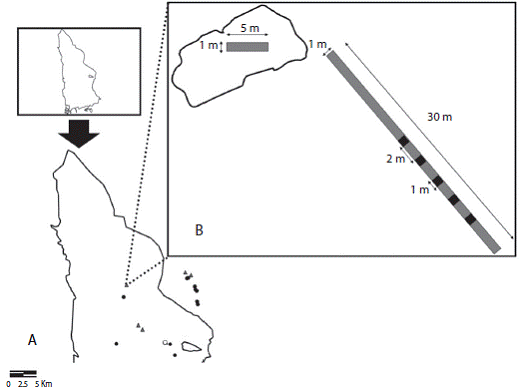
Figigua 3: Ubicación espacial de los parches de Quercus coccifera (triángulo gris), Smilax aspera (círculo negro) y Rubia peregrina (círculo blanco) en el área quemada y no quemada circundante en que se colocaron los tratamientos de exclusión y no exclusión de las ramas/racimos (A) y diseño experimental de la estimación de frutos para cada parche de vegetación (B). 0 2.5 5 Km
Análisis estadístico: En el experimento de remoción predispersiva, la variable respuesta fue la diferencia del número de frutos remanentes entre las ramas excluidas y las no excluidas, a lo largo del tiempo. Esta diferencia representa el nivel de caída y/o remoción de semillas obtenidas al comparar ambos tratamientos de exclusión de vertebrados. Los promedios de remanencia para cada rama se obtuvieron calculando la cantidad de semillas para cada tiempo con base a la producción inicial de fru tos de cada rama. Para el análisis de los datos se orga nizaron los promedios de remanencia de las ramas y se aparearon al azar las ramas excluidas y no excluidas para calcular las diferencias entre las medias obtenidas. Las ramas que no pudieron ser apareadas fueron eliminadas del análisis, así como las ramas que por motivos azarosos (incidencia de nieve, caída de árboles) fueron afectadas y no lograron ser evaluadas hasta el final del experimento. Para el caso de Quercus coccifera, la diferencia de medias se obtuvo desde el día cero (instalación del experimento) hasta los 75 días de muestreo mientras que en el caso de Smilax aspera fue del día cero a los 176 días aproxi madamente. Dado que los porcentajes o proporciones forman una distribución binomial (Zar, 1984) en el caso de Q. coccifera se realizó un Test de Mann-Whitney para evaluar las diferencias de la variable respuesta en fun ción del hábitat (área quemada y no quemada) en el pe riodo de muestreo del 2008. Se pudo disponer de una n = 8 de forma que los tamaños de muestra entre ambos hábitats fueran balanceados. Se suprimieron del análisis los datos de remoción predispersiva Q. coccifera del 2009 por cuanto no se pudieron seleccionar ramas en el área no quemada debido a la poca disponibilidad de frutos. En el caso de los datos de S. aspera se verificó la homoge neidad de varianzas y se realizó una transformación de la raíz cuadrada del arco-seno de los promedios de rema nencia de semillas para mejorar la normalidad de los da tos. Mediante un ANDEVA de dos vías se evaluó el efecto del periodo de muestreo (2008-2009) para cada hábitat (quemado "vs" no quemado) previa eliminación de los datos sobrantes, lo que condujo a un tamaño muestral n=4 para cada nivel del factor, evitándose así la replicación desigual (Zar, 1984). Ambas pruebas se realizaron usando el paquete estadístico STATISTICA 6.0 y se usó un nivel de significancia p<0,05.
Resultados
La remoción de bellotas de Quercus coccifera pre sentó diferencias significativas entre el hábitat quema do y el no quemado (Test de Mann-Whitney, z=2,5205; p<0,05) siendo mayor en el área no quemada que en la quemada. Las curvas de remanencia de frutos excluidos y no excluidos a lo largo del tiempo en el área quema da presentaron un comportamiento similar ambos años. En este hábitat se observó una diferencia casi nula en la caída de frutos entre ambos tratamientos, siendo causa da principalmente por la maduración de las semillas. En contraste, en el hábitat no quemado la diferencia de re moción de frutos entre ambos tratamientos es importan te, observándose que en el tratamiento de no exclusión, a los 40 días de muestreo, quedaban disponibles menos de un 15% de las bellotas. El estado de maduración de las bellotas parece no ser un factor determinante en la remoción de frutos, lo que se evidenció en el comporta miento de remoción de los frutos desde el comienzo del experimento en el área no quemada (Figura 4).
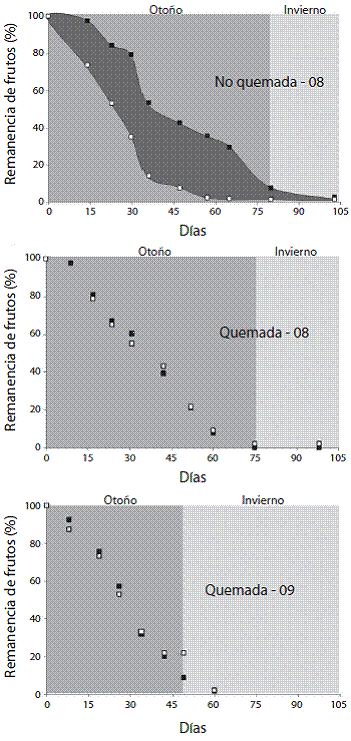
Figura 4: Porcentaje de remanencia de bellotas de Quercus cocci-fera (estimada con base a la producción inicial de frutos) para ramas excluidas (cuadros negros) y no excluidas (cuadros blan cos) a lo largo del 2008 en ambos hábitats y 2009 en la zona quemada. El área sombreada en el gráfico superior representa la tasa de remoción de bellotas por animales.
En el caso de Smilax aspera el comportamiento de re moción y perdida de semillas producto de la senescencia de los frutos fue similar al comparar ambos hábitats pero fue mayor en el año 2008 en ambos hábitats (Análisis de Varianza de dos vías, Hábitat F=0,48;p=0,50, Año F=5,98; p<0,05 HxA F=3,56; p=0,08). En el 2008 a la mitad del periodo de evaluación (90 días de la instalación de los tratamientos) la remanencia de frutos en el tratamiento de no exclusión en ambas áreas de muestreo represen taba valores inferiores al 20% de la cantidad inicial de frutos lo que contrasta con la remanencia de frutos en ramas excluidas (>70%). En el año 2009 se presenta una tendencia hacia la similitud en los tratamientos con una pérdida de frutos más sostenida a lo largo del tiempo y aparentemente más asociada a la caída natural de los mismos (esto es corroborado por el comportamiento de caída de los frutos excluidos) producto de la maduración de los frutos (Figura 5).
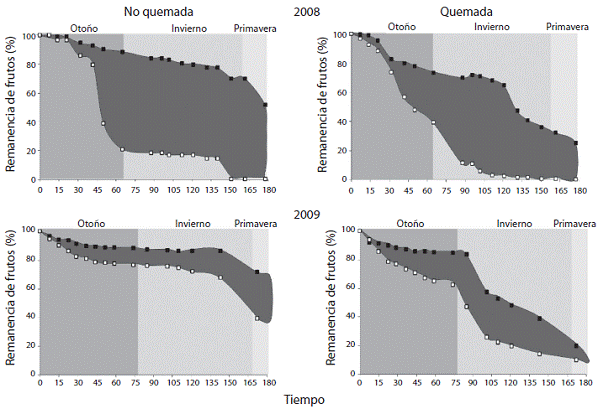
Figura 5: Porcentaje de remanencia de frutos de Smilax aspera (estimada con base en la producción inicial de frutos) para ramas exclui das (cuadros negros) y no excluidas (cuadros blancos) a lo largo del tiempo en 2008 y 2009 en las zonas no quemada y quemada. El área sombreada representa la remoción de frutos por animales.
La remoción de frutos en función de su estado de ma duración difirió entre la especie de frutos no carnosos y las de carnosos. Las bellotas de Quercus coccifera fueron removidas fuertemente desde el inicio del muestreo (partiendo de frutos verdes) mientras que en el caso de Smilax aspera se observó un incremento sustancial en la remoción de frutos conforme avanzaba el tiempo y se daba el proceso de maduración de la diáspora (Fig. 5). Este hecho fue evidenciado también en una evaluación adicional que se realizó en el área quemada para otra es pecie de fruto carnoso (Rubia peregrina) que empezó a experimentar la remoción de semillas a partir de frutos maduros por parte de roedores (Figura 6).
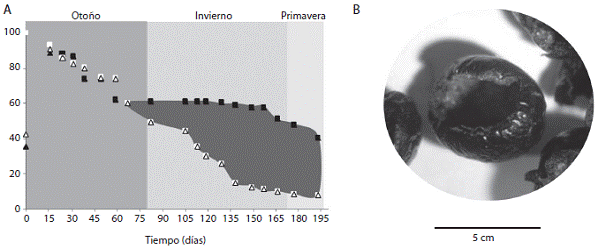
Figura 6: A. Remoción de diásporas de Rubia peregrina a través del tiempo. El área sombreada representa la remoción de frutos por animales.Nótese el inicio e incremento de la remoción de diásporas a partir de los 60 días de iniciado el muestreo cuando empieza a ocurrir la maduración de la diáspora B. frutos maduros con las semillas extraídas provenientes del tratamiento de no exclusión en el área quemada, invierno del 2008-2009.
En cuanto a la estimación de la producción de frutos se observaron variaciones interanuales en la produc tividad de ambos hábitats siendo el hábitat quemado el más productivo en el 2008 y el no quemado en el 2009 (Cuadro 1). La mayor productividad del 2008 está concentrada en la producción de frutos por parte de las dos especies de estudio Quercus coccifera y Smilax aspera mientras que en el 2009 fueron las especies carnosas Pistacia lentiscus y S. aspera las que tuvieron una mayor productividad (Cuadro 2). En condiciones de parche se observa una mayor cantidad de frutos de las dos espe cies de estudio al compararlas con áreas contiguas a los parches, en las cuales se notó una dominancia especial mente de bellotas de Q. coccifera en el hábitat quemado en ambos períodos de muestreo (Cuadro 3).
Cuadro 1: Producción promedio global (frutos/m2) de frutos carnosos y no carnosos disponibles en los sitios del marcaje de ramas para el experimento de remoción de semillas. Muestreos realizados en Otoño del 2008 y 2009
Cuadro 2: Promedios estimados de la producción de frutos (frutos/m2) del total de parches seleccionados para cada especie de estudio en el área quemada y no quemada en los Otoños del 2008 y el 2009
Discusión
De las dos especies principales analizadas en el expe rimento de remoción predispersiva de semillas, Quercus coccifera fue la especie que presentó un comportamien to consistente y claramente diferenciado en el hábitat quemado al compararlo con en el hábitat no quemado, durante ambos años de muestreo. Como se esperaba para el área quemada, la remoción de bellotas por parte de vertebrados fue prácticamente nula probablemente como consecuencia de las características alteradas del hábitat que estaría condicionando el comportamiento de remoción por parte de roedores. La poca disponibi lidad de cobertura a nivel de dosel debido al efecto del fuego y la tala producto de la gestión del área quemada, podría limitar la incidencia de estos granívoros los cua les prefieren actuar en sitios con mayor cobertura para disminuir el riesgo de depredación (Torre & Díaz, 2004; Pons & Pausas, 2007; Perea, San Miguel & Gil, 2011a). En contraste, las mayores tasas de remoción de semillas en el hábitat no quemado sugiere que a pesar del ma yor esfuerzo que podría representar el tener que escalar plantas de mayor tamaño a las encontradas en el área quemada seguramente este el área no quemada ofrece la sensación de un mayor sentido de protección a los roedores. Las buenas habilidades escaladoras de los roe dores removiendo frutos en plantas en la fase pre-disper siva ya ha sido observada también sobre frutos carnosos y bellotas del genero Quercus en sitios con abundante cobertura arbórea y arbustiva (Bas et al., 2005, Perea et al., 2011b).
En el caso de Smilax aspera, el tipo de hábitat (que mado o no quemado), no fue un factor determinante en la remoción de frutos pero si se observaron diferencias marginalmente significativas al considerar el factor in teranual en interacción con el hábitat. En el 2008, la re moción de frutos por animales fue más rápida en el área no quemada que en la quemada. De otro lado, tanto en 2008 como en 2009 la pérdida de frutos por senescencia (ramas excluidas) al final del periodo de estudio fue ma yor el área quemada, posiblemente debido a la mayor in solación que reciben las plantas productoras. Se observó a lo largo del tiempo en el 2009 una remanencia sosteni da de diásporas (especialmente para el caso del área no quemada) en concordancia con la mayor productividad de frutos para la especie y en términos globales durante ese año. La remanencia sostenida de frutos de S. aspera en las ramas este año podría ser consecuencia del fenó meno de saciedad debido a la alta abundancia de frutos carnosos ('masting events") en el ambiente como fue evi denciado en la estimación de producción de frutos es pecialmente al considerar el hábitat no quemado o bien por los patrones de selección de alimento dado que se ha observado como las aves frugívoras tienden a selec cionar frutos de Smilaxsp. en algunos años mientras no los consumen en otros años (Herrera, 1998).
Los patrones de remoción por vertebrados sobre los frutos de ambas especies parecen diferir en relación al estado de maduración solo para el caso de los frutos car nosos. Es así como la remoción de bellotas de Q. coccifera ocurrió continua e indistintamente sobre frutos verdes y maduros mientras que en el caso de los frutos carnosos la mayor remoción de diásporas ocurrió conforme madu raban las diásporas. Esto fue corroborado también para otra especie de fruto carnoso en el área de estudio (Rubia peregrina), en la cual se pudo constatar cuatro eventos de remoción de semillas 105 días después del inicio del experimento cuando ya los frutos estaban maduros. Bajo estas plantas se observaron numerosos restos de pulpa y diásporas enteras maduras cuidadosamente abiertas sin las semillas. Este comportamiento asociado a roedo res denota una clara preferencia de remoción sobre las semillas, y no sobre la pulpa, de los frutos en función del mayor contenido nutricional de la primera estructura (Perea, San Miguel & Gil, 2011c).
Para las dos especies de frutos carnosos se observó una larga permanencia de frutos turgentes en las ramas no excluidas. Esta situación fue más allá del inicio de la primavera y conduce a pensar que la exposición pro longada de las diásporas y la presencia de antocianinas aumentan su probabilidad de detección por diversas especies de roedores y aves granívoras y frugívoras. Las evidencias de remoción de semillas por parte de roedo res y la menor remoción para S. aspera en 2008 en el área quemada (en contraste con la abultada curva de remo ción para el área no quemada) sugieren un aprovecha miento del recurso por parte de roedores especialmente en el área quemada. Esto podría deberse a una menor diversidad y abundancia de aves frugívoras en el área quemada debido a la menor complejidad estructural del hábitat producto de la gestión forestal. Rost, Pons y Bas (2009) y Rost, Clavero, Bas y Pons (2010) encontra ron una mayor actividad de aves frugívoras en sitios en que las actividades de gestión post-incendio generaron estructuras complejas con madera apilada en el suelo. En Ventalló, la gestión post-incendio extrajo los troncos completos para la producción de energía a partir de la biomasa, por tanto la generación de estructuras que pu dieran funcionar como perchas es menor y más bien lo que prevalecía en varias partes del área son numerosos restos de ramas pequeñas desordenados que podrían favorecer la acción de los roedores debido a una mayor cobertura a nivel de suelo. Finalmente se debe resaltar que dada la variación en la permanencia de frutos en las ramas entre las dos especies de estudio parece cla ro observar diferencias en los patrones de remoción de los frutos de especies carnosas y no carnosas. Q. coccifera es una especie de fructificación otoñal y sus bello tas desarrolladas están disponibles muy poco tiempo en las ramas. Esta situación en combinación con la escasa cobertura en el caso del área quemada post-incendio podría suponer más bien un aprovechamiento de las be llotas por parte de roedores pero a nivel post-dispersivo. Contrariamente, las diásporas de las especies carnosas al permanecer más tiempo sobre las ramas tendrían una mayor probabilidad de aprovechamiento a nivel pre-dispersivo pero dicha situación parece estar siendo con dicionada por la comunidad de granívoros y frugívoros existente y por la disponibilidad de recursos alimentarios en el área.

