











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Los sistemas ganaderos proporcionan múltiples beneficios a los seres humanos, ofrecen productos de origen animal ricos en proteína que contribuyen a la seguridad alimentaria, fomentan el empleo y dinamizan la economía rural, además se convierten en capital disponible y fuerza de trabajo en muchos países en desarrollo (Sekaran et al., 2021). En Colombia, la ganadería es una de las actividades productivas que más aporta al sector rural del país, con una participación del 53 % del producto interno bruto (PIB) pecuario, 19,5 % del agropecuario y 1,3 % del PIB nacional (Enciso et al., 2018).
En el piedemonte amazónico colombiano, la ganadería bovina es la principal actividad productiva y se posiciona en el quinto lugar en el inventario bovino nacional (Enciso et al., 2018). Sin embargo, esta actividad pecuaria presenta baja productividad y rentabilidad, con un impacto ambiental negativo y uno de los núcleos de deforestación de mayor persistencia a nivel nacional (Enciso et al., 2018; Pardo Rozo et al., 2020).
La baja productividad zootécnica de los sistemas pecuarios del piedemonte amazónico y la alta tasa de deforestación (34 % de la deforestación nacional) (Forero et al., 2016), puede comprometer el cumplimiento de las metas ambientales nacionales de los sistemas pecuarios locales.
Para mantener el incremento anual de temperatura ambiente por debajo de 2 ºC a través de baja emisión de gases de efecto invernadero (GEI) por parte de la ganadería (Convención Marco de las Naciones Unidas sobre el Cambio Climático, 2015), se requiere una mayor eficiencia en la producción ganadera a través de prácticas sostenibles. Por ello, se ha planteado el uso de estrategias para mejorar el rendimiento productivo animal, disminuir la deforestación y el impacto ambiental generado por la ganadería bovina extensiva, al considerar como eje principal al árbol (Valencia-Salazar et al., 2018).
Los árboles y arbustos nativos son fundamentales para la rehabilitación productiva y la conservación de la biodiversidad en los paisajes ganaderos (Murgueitio et al., 2011). La presencia de especies leñosas en los potreros puede contribuir a la disminución de la deforestación y degradación de los suelos, la cual se ha incrementado de manera acelerada en los últimos años (Cabrera-Núñez et al., 2019).
La mayoría de las especies arbóreas y arbustivas poseen abundante forraje, algunos contienen frutos y vainas que pueden ser utilizados también en la alimentación animal (Pinto-Ruiz et al., 2010), se ha reportado que las hojas de Albizia niopoides (Spruce ex Benth.), Gliricidia sepium (Jacq.) Kunth ex Walp., Leucaena leucocephala (Lam.) de Wit, Samanea saman (Jacq.), Moringa oleifera Lam., Cordia dentata Poir. y Guazuma ulmifolia Lam. poseen un alto contenido de proteína comparados con las gramíneas tropicales, considerándose como una alternativa promisoria, debido a su gran potencial como forrajeras (Pérez-Almario et al., 2013).
En el piedemonte amazónico colombiano se han caracterizado especies como Clitoria fairchildiana R.A. Howard y Piptocoma discolor (Kunth) Pruski, los resultados indican aceptables contenidos nutricionales que posibilitan su uso en alimentación animal (Riascos-Vallejos et al., 2020; Suárez et al., 2008). Sin embargo, aún se hace necesario llevar a cabo investigaciones relacionadas con el reconocimiento, evaluación y selección de especies promisorias, debido a la gran diversidad florística presente en la zona.
El objetivo de la presente revisión fue recopilar y analizar información producto de investigaciones sobre las especies forrajeras Erythrina poeppigiana (Walp.) O.F. Cook, C. fairchildiana, P. discolor y G. ulmifolia.
Influencia de la ganadería bovina en el piedemonte amazónico colombiano
La Amazonía, al ser la mayor selva tropical del mundo, desempeña un papel fundamental para la tierra, influye en los flujos globales de carbono, el ciclo del agua y la biodiversidad (Cherubin et al., 2018). En el piedemonte amazónico colombiano, la ganadería continúa siendo una de las principales actividades económicas (Pardo Rozo et al., 2020). Sin embargo, esta amplia región se colonizó por parte del hombre de manera desproporcionada, sin medir las implicaciones ocasionadas por la intervención de un ecosistema frágil, con manejo inapropiado del recurso suelo, especies vegetales no adaptadas con deficiente manejo de praderas y animales (Cuesta Muñoz & Gómez Mesa, 2002).
Se ha documentado que un sistema ganadero de doble propósito en el piedemonte amazónico se caracteriza por una baja carga animal (0,35-1,71 UGG/ha), ganancias de peso de 321,5 g/día y entre 4,3 y 8 L de producción de leche por vaca día (Enciso et al., 2018; Torrijos et al., 2017), con emisiones de GEI de 19,6 kg CO2 eq/kg de carne y 1,63 kg de CO2 eq/L de leche, lo que permite inferir que el 85 % de estas emisiones, provienen de la fermentación entérica (Enciso et al., 2018).
El establecimiento de pasturas para ganadería bovina se ha realizado a partir de la tala y quema de bosques de manera acelerada en los últimos cincuenta años, con posibles cambios irreversibles en la biodiversidad y el funcionamiento del ecosistema (Brando et al., 2020). En la actualidad, los bosques amazónicos son transformados en pastizales para la implementación de sistemas ganaderos extensivos, caracterizados por su baja inversión tecnológica y bajo rendimiento productivo (Mora-Marín et al., 2017; Murcia García et al., 2007).
Los cambios de cobertura natural implican procesos de tala y quema de bosque, los reportes de deforestación en la región amazónica colombiana para el periodo comprendido entre los años 2017 - 2018 fueron de 138 176 ha, comparados con 56 962 ha deforestadas en los años 2014 - 2015 (Olaya López, 2022), donde los departamentos con mayores pérdidas de cobertura boscosa fueron Caquetá (43,7 %), Meta (16,3 %), Putumayo (15,9 %) y Guaviare (12,2 %) (Guarnizo et al., 2020).
La producción ganadera en el piedemonte amazónico colombiano se basa en el uso de gramas naturales y pastos introducidos, entre los que predominan diferentes especies del género Urochloas sp. (syn. Brachiarias) (Rivera et al., 2015). Este tipo de gramíneas presentan bajos niveles de proteína cruda (6,03 %), digestibilidad (48,1 - 55,1 %) y altos niveles de fibra (FDN: 67,3 - 80,8 % y FDA: 39,5 - 55,8 %), situación que se acrecienta durante los meses de menores precipitaciones donde se disminuye la oferta forrajera y se afecta la productividad animal (Rivera et al., 2015; Suárez Salazar et al., 2008; Velásquez & Muñoz Ramos, 2006).
Los sistemas ganaderos de esta región requieren una reconversión productiva donde se reemplacen los monocultivos de gramíneas por la combinación de pastos naturalizados y especies vegetales perennes, con mayor tolerancia y adaptación a las particularidades de los suelos amazónicos (Landínez-Torres, 2017), una alternativa es la implementación de sistemas silvopastoriles (SSP) (Gómez Fuentes Galindo et al., 2017).
Se ha demostrado que los SSP permiten la interacción de una o más especies vegetales en diferentes estratos, en un mismo espacio y periodo de tiempo (Murgueitio et al., 2016). Además, de considerarse como un factor determinante en la recuperación de suelos y pasturas degradadas, mitigación de las emisiones de GEI, la generación de servicios ecosistémicos, reforestación y conservación de la biodiversidad, la reducción de la huella de carbono y adaptación al cambio climático, frente a la ganadería extensiva tradicional (Sotelo et al., 2017).
Características edafoclimáticas y usos de especies arbóreas y arbustivas del piedemonte amazónico
Factores como las variaciones en la intensidad y frecuencia de las precipitaciones, los fenómenos del Niño y la Niña, así como las altas temperaturas, generan efectos importantes en la producción de alimentos (Riascos-Vallejos et al., 2021), sumado a esto, las pasturas tropicales por la calidad de los suelos en la amazonia se caracterizan por presentar baja producción de biomasa y baja calidad nutricional (Rivera et al., 2015), razones por las que los productores ven la necesidad de encontrar nuevos materiales que ayuden a mejorar la oferta alimenticia para sus animales (Gallego-Castro et al., 2017).
Las condiciones edafoclimáticas de la región amazónica, condicionan la actividad ganadera, se han observado precipitaciones de hasta los 3500 mm anuales, temperaturas superiores a los 30 °C y humedad relativa del 86 %(Instituto de Hidrología, Meteorología y Estudios Ambientales ((IDEAM), 2018), no obstante, las plantas arbustivas y arbóreas surgen como una alternativa de producción con aceptables características de adaptación (Riascos-Vallejos et al., 2020). Es el caso de las especies forrajeras E. poeppigiana, C. fairchildiana, P. discolor y G. ulmifolia, las cuales se encuentran presentes en bosques de regeneración y secundarios en el piedemonte amazónico (Cuadro 1).
Cuadro 1 Características edafoclimáticas y usos de especies leñosas del piedemonte amazónico colombiano reportadas por varios autores. Colombia. 2022.
| Especie forrajera | Altitud msnm | Precipitación mm año | Usos | Condiciones suelos | Referencia |
| Erythrina poeppigiana | 600 - 1300 | 1000 - 2500 | Bancos forrajeros Sombra Restaurador de suelos Cercas vivas Fijación de N | Tolera suelos de alto nivel freático e incluso inundaciones Textura arcillosa a franca | Botero & Russo (1999) |
| Clitoria fairchildiana | 0 - 1200 | 2000 - 3600 | Bancos forrajeros Cercas vivas Fijación de N Restaurador de suelos | Tolera suelos con bajo nivel de fertilidad, alto grado de acidez y de saturación de aluminio | Jaquetti et al. (2021) |
| Piptocoma discolor | 0 - 1000 | 2500 - 3700 | Regenerador de suelos Bancos forrajeros Madera Dendroenergético | Tolera suelos arcillosos, ácidos, bajos en fósforo, con altos contenidos de aluminio | Hurtado Ducuara & Guayara Suárez (2013); Riascos-Vallejos et al. (2020); Saltos et al. (2015) |
| Guazuma ulmifolia | 0 - 1600 | 700 - 1500 | Bancos de proteína Cercas vivas Medicinal (antioxidante, antimicrobiano) | Tolera suelos con alto contenido de aluminio, arcillosos y de alta compactación | Araujo Pereira et al. (2019); Rojas-Rodríguez & Torres-Córdoba (2019) |
E. poeppigiana es una leguminosa perteneciente a la familia Fabaceae (Espeche et al., 2020) al igual que C. fairchildiana (Vale-Montilla, 2019). P. discolor pertenece a la familia Asteraceae (Hurtado Ducuara & Guayara Suárez, 2013) y G. ulmifolia, es una Esterculiaceae (Matulevich Pelaez & Garcia Rodríguez, 2016). Estas especies son propias del neotrópico, presentan varias potencialidades como alternativa de alimento para rumiantes durante la época de mínima precipitación, con escases forrajera, están adaptadas a los suelos degradados de baja fertilidad y a las condiciones climáticas cambiantes del trópico, lo que garantiza su supervivencia, persistencia y crecimiento (Rodríguez Fernández & Fandiño, 2013).
Por las características de ubicación del piedemonte amazónico, las condiciones de humedad, precipitación y temperatura que propician la abundancia de flora y fauna (Pardo Rozo et al., 2020), es factible la incorporación en sistemas ganaderos de especies arbustivas como E. poeppigiana, C. fairchildiana, P. discolor y G. ulmifolia, las cuales toleran altos niveles de precipitación (entre 1500 y 3 600 mm/año), propios de la región amazónica donde prevalece el bosque húmedo tropical (Padilla-Gil, 2019).
Estudios reportan que la utilización de árboles en sistemas ganaderos tiene efectos positivos en la recuperación y conservación de suelos. La presencia de M. oleifera contribuye, en el corto plazo, a recuperar las características biológicas del suelo en condiciones de bosque húmedo tropical (Navas-Panadero & Ríos de Álvarez, 2019), al igual se considera que E. poeppigiana y C. fairchildiana por ser leguminosas tienen la capacidad de establecer relaciones simbióticas con microorganismos fijadores de nitrógeno (bacterias del genero Rhizobium), considerándose importantes en la recuperación de suelos degradados (Bianco & Cenzano, 2018; Pedrozo et al., 2018).
Composición química y producción de biomasa de especies arbóreas y arbustivas del piedemonte amazónico
En regiones tropicales como el piedemonte amazónico, el recurso básico para la obtención de la energía alimentaria para los rumiantes son los forrajes herbáceos, gramíneas, que por lo general no cubren los requerimientos nutricionales de los animales (Barahona & Sanchez, 2005). La incorporación de árboles y arbustos por medio de los SSP se considera una opción para obtener forraje de alto valor nutricional y a la vez aprovechar los múltiples servicios ambientales derivados de su implementación (Murgueitio et al., 2011).
Varios árboles y arbustos nativos de Mesoamérica como las Erythrinas spp., G. sepium, L. leucocephala, G. ulmifolia y Prosopis juliflora, entre otros, presentan altos contenidos de proteína cruda (14 a 32 %) (Cuadro 2) y aceptable producción de biomasa forrajera con valores que oscilan entre 7 y 14 t/ha/año MS, cuando se siembran a una densidad de 20 000 a 40 000 plantas por hectárea y con una frecuencia de corte entre 3 y 6 meses (Ibrahim et al., 2004).
Cuadro 2 Valores promedio de composición nutricional de las especies seleccionadas reportada por varios autores para condiciones tropicales. Colombia. 2022.
MS%: materia seca/dry matter. PC%: proteína cruda/crude protein. FDN%: fibra en detergente neutro/neutral fibre detergent. FDA%: fibra en detergente acido/acid fibre detergent. ES: época seca/dry season. EL: época de lluvias/rainy season.
Las especies analizadas se pueden considerar de elevado potencial forrajero. Estudios reportan una producción de biomasa promedio de C. fairchildiana, de 861,5 g/planta de MS, en dos tipos de suelos del piedemonte amazónico colombiano (Suárez et al., 2006). En la actualidad la búsqueda de alternativas de alimentación ha permitido evaluar varias especies promisorias. Al comparar el rendimiento productivo de Morus alba L., Hibiscus rosa-sinensis L. y E. poeppigiana, se encontraron diferencias significativas a favor de esta última, relacionadas con una mayor relación de hojas por tallo y producción de forraje (683,5 g/planta de MS), lo que demuestra su elevada capacidad de producción de biomasa forrajera (Vargas Burgos et al., 2015).
P. discolor, es una especie de fácil adaptación y con gran capacidad de producción en zonas de regeneración natural, reportes indican un rendimiento de 180,56 g de MS/planta a los 60 días post corte en el piedemonte amazónico del departamento del Caquetá (Hurtado Ducuara & Guayara Suárez 2013). Sobresale por su rápido crecimiento, en comparación con otras forrajeras tropicales (Hernández Benalcázar et al., 2015). En cuanto a la producción de forraje de G. ulmifolia, existen reportes de 171 g/planta de MS a los 45 días (Vargas & Quintero, 2017) y 390 g/planta de MS con 120 días de recuperación (Rodríguez Fernández & Fandiño, 2013).
La inclusión de especies nativas ofrece la posibilidad de mejorar el conocimiento del potencial productivo de los recursos forrajeros que forman parte del patrimonio genético y que puede contribuir con las soluciones tecnológicas demandadas por los productores en la actualidad (Rodríguez Fernández & Fandiño, 2013).
Potencial para la disminución de CH4 entérico de plantas arbustivas y arbóreas del piedemonte amazónico
Las regiones tropicales de todo el mundo presentan una alta variedad de árboles, arbustos y plantas herbáceas de elevada composición nutricional (14 a 32 % de proteína cruda) y con potencial para ser utilizadas como alternativa alimenticia en los sistemas de producción ganaderos (Valencia-Salazar et al., 2021). Esta diversidad de especies favorece procesos de recuperación de suelos degradados, fijación de nitrógeno y la disminución de las emisiones de CH4 de origen ruminal, debido al contenido de metabolitos secundarios en el tejido vegetal (Arango et al., 2020).
La pérdida de energía en forma de CH4 tiene una importancia significativa, debido a que las dietas de los bovinos en el trópico se basan en el pastoreo gramíneas nativas y pastos naturalizados con bajo contenido de proteína cruda (<7 %), digestibilidad (<50 %) y alto contenido de carbohidratos estructurales (60-80 % FDN) (Benitez et al., 2017; Ku-Vera et al., 2013; Piñeiro-Vázquez et al., 2017). Este tipo de dietas, generan una alta proporción de ácido acético, baja producción de propionato y un aumento en la emisión de CH4 entérico (Archimède et al., 2014).
Lograr una reducción en la producción de CH4 en los rumiantes es uno de los principales objetivos para reducir las emisiones de GEI y mejorar el rendimiento productivo animal (Hristov et al., 2013).
La mayoría de plantas producen diversos compuestos biológicos que son clasificados como metabolitos primarios y secundarios (Cardona-Iglesias et al., 2016). Las moléculas activas que participan en procesos como el crecimiento, el desarrollo y la reproducción de las plantas, son considerados metabolitos primarios (Bodas et al., 2012). Los metabolitos secundarios actúan en la planta como mecanismos de defensa contra agentes externos, ataques de insectos o adaptación a condiciones climáticas adversas y su presencia puede variar entre especies por el estado fenológico de la planta y por factores ambientales (Patra & Saxena, 2011; Ramakrishna & Ravishankar, 2011; Wallace, 2004).
En términos de acción metabólica, el efecto de los metabolitos secundarios en el rumen se fundamenta en la inhibición/control del crecimiento/desarrollo de protozoos y arqueas metanogénicas (Patra & Saxena, 2009a). La acción de los taninos depende de su estructura química, así como de la especie bacteriana (Vasta et al., 2019), estos provocan una alteración en la permeabilidad y cambios morfológicos en las membranas de los microorganismos ruminales (Scalbert, 1991; Smith et al., 2005) (Figura 1). Sin embargo, la respuesta sobre el rendimiento animal y la disminución de metano, dependerá de las dosis, estructura química, peso molecular y composición de la dieta (Ortiz et al., 2014).
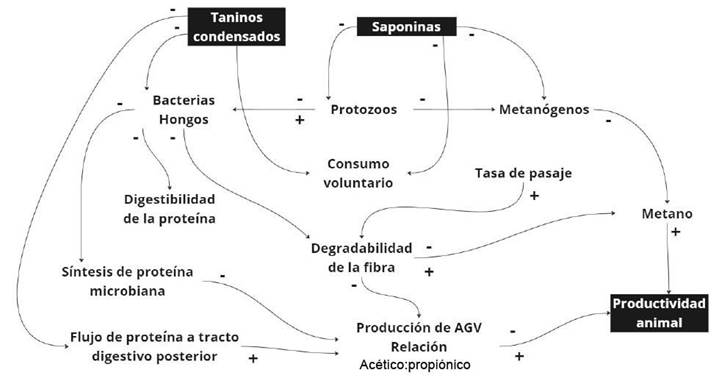
Figure 1 Representation of the mechanism of action of condensed tannins and saponins on rumen microbiota and methane production. Adapted from Patra and Saxena (2009b).
Se han llevado a cabo varios estudios para evaluar las alternativas de reducción del CH4 entérico por acción de los metabolitos secundarios de las plantas. Por lo anterior, se ha documentado que la inclusión de taninos de Acacia mearnsii De Wild. disminuye en un 43 % la producción de metano in vitro (Durmic et al., 2014). De igual manera, Pérez-Can et al. (2020) reportaron que la inclusión del forraje de especies arbóreas y arbustivas como L. leucocephala (14,47 mL CH4/g/MS), G. sepium (40,49 mL CH4/g/MS), Tithonia diversifolia (Hemsl.) A. Gray (20,17 mL CH4/g/MS) y G. ulmifolia (34,06 mL CH4/g/MS) tienden a disminuir la producción de metano in vitro (Cuadro 3).
Cuadro 3 Producción de metano in vitro y perfil fitoquímico de E. poeppigiana (Walp.) O.F. Cook, C. fairchildiana R.A. Howard, P. discolor (Kunth) Pruski y G. ulmifolia Lam, reportada por varios autores para condiciones tropicales. Colombia. 2022.
| Especie forrajera | mL CH4 g MS fermentada | % de MSP | Presencia de MSP | Referencia | ||
| TC | SP | TC | SP | |||
| Erythrina poeppigiana (Walp.) O.F. Cook | 22,1 | 0,4 | ND | +++ | +++ | Aragadvay-Yungán et al. (2021) Rodríguez & Ledesma (2014) |
| Clitoria fairchildiana R.A. Howard | ND | 0,39 | ND | ++ | - | Aragadvay-Yungán et al. (2021) Riascos-Vallejos et al. (2020) |
| Piptocoma discolor (Kunth) Pruski | ND | 0,27 | ND | ++ | + | Riascos-Vallejos et al. (2020) |
| Guazuma ulmifolia Lam | 37,98 ND 34,06 | ND 0,91 0,47 | ND ND 0,91 | ND ++ +++ | + - + | Galindo et al. (2014) Ojeda et al. (2018) Pérez-Can et al. (2020) |
+++ presencia abundante/high presence ++ presencia moderada/moderate presence + presencia baja - ausente/low presence - abscence. MSP: metabolitos secundarios de las plantas/plant secondary metabolites. TC: taninos condensados/condensed tannins. SP: saponinas/saponins. ND: no determinado/undetermined.
En C. arborea Benth., se ha identificado la presencia de taninos (1,3 % taninos condensados y 1,9 % fenoles totales) que demuestran una posible reducción en la producción de metano in vitro, atribuida a estos metabolitos (1,23 mL CH4 g MS 24 h) (Aragadvay-Yungán et al., 2021). Para P. discolor no se encontraron datos en la literatura sobre su efecto en la reducción de CH4; no obstante, se ha demostrado que posee un contenido de proteína adecuado para la nutrición de rumiantes (Riascos-Vallejos et al., 2020).
La composición nutricional es un criterio útil para la selección de especies con potencial de alimentación y mitigación del CH4 en rumiantes (Albores-Moreno et al., 2018), se puede considerar a esta especie como promisoria para el piedemonte amazónico colombiano.
Estudios in vivo realizados por Molina-Botero et al. (2019), demostraron que la inclusión de vainas de Enterolobium cyclocarpum (Jacq.) Griseb. y forraje de G. sepium en un 15 y 30 % de la materia seca de la ración, disminuyó las emisiones anuales de metano por unidad de producto, sin afectar la ingesta diaria de materia seca. Al incluir el 4 % de taninos hidrolizables y taninos condensados en la dieta de corderos, no se afectó la población de bacterias y hongos ruminales (Salami et al., 2018). Sin embargo, los mismos autores reportaron que los taninos condensados redujeron de manera significativa la abundancia de microorganismos metanogénicos.
La mayoría de los compuestos fitoquímicos presentes en las hojas de los árboles y arbustos tropicales, modulan la fermentación ruminal e inhiben la producción de metano, por lo tanto, representan una alternativa viable de mitigación de las emisiones de GEI (Barahona et al., 2003; Patra & Saxena, 2011).
El contenido de metabolitos secundarios en la ración debe contribuir a disminuir la metanogénesis, sin afectar la digestibilidad, el perfil de fermentación o la tasa de sobrepaso de materia seca en el rumen (Cardona-Iglesias et al., 2016). Además, se ha demostrado que el follaje de E. americana Mill. con una edad de rebrote de 60 a 120 días, presentó un contenido de taninos condensados entre 0,6 a 1,4 % que no afectó el consumo voluntario en pequeños rumiantes (Hernández-Espinoza et al., 2020).
Algunos autores aseguran que la concentración de taninos en la dieta de los rumiantes debe variar del 2 al 4 % de la MS (Otero & Hidalgo, 2004; Waghorn et al., 1999). Al incluir niveles bajos de taninos en la dieta de vacas lecheras (0,2 %) se demostró la inhibición directa de los protozoos y una consecuente disminución en la producción de CH4 en el rumen (Cieslak et al., 2012). Se indica que una concentración de taninos condensados de 0,5 % a 3 %, tiene la capacidad de mitigar la producción de CH4 sin afectar la digestibilidad de la MS (Saminathan et al., 2015). Concentraciones por encima del 5 % de la MS pueden afectar la palatabilidad, el consumo y reducir la eficiencia en la utilización del alimento (Mueller-Harvey, 2006; Waghorn, 2008).
Sistemas silvopastoriles con arbóreas y arbustivas del piedemonte amazónico como estrategia de adaptación al cambio climático: bienestar animal y captura de carbono
En los últimos años, la sostenibilidad de los sistemas de producción bovina ha sido cuestionada por los impactos ambientales y sociales que se le atribuyen (generación y emisión de GEI, pérdida de biodiversidad, deterioro de las fuentes de agua y degradación de los suelos). Sin embargo, la producción ganadera es una actividad indispensable e insustituible para el desarrollo de la humanidad (Naranjo-Ramírez & Ruiz-Buitrago, 2020). El manejo extensivo de los sistemas ganaderos ha contribuido a que el sector sea relacionado con el calentamiento global. No obstante, existen alternativas de manejo que permiten mitigar las emisiones y crear modelos que se adapten a los efectos adversos del cambio climático.
El silvopastoreo es un modelo de producción pecuaria en donde se asocia un componente arbóreo/arbustivo con uno herbáceo (pasturas naturales o mejoradas) y otro pecuario (ganado) en un mismo sitio, de manera que existan interacciones biológicas entre ambos con el objetivo de maximizar el uso de la tierra (Russo, 2015). De igual manera se ha demostrado que brindan otra serie de servicios como la producción de madera, frutos, sombra, regulación hídrica y térmica, hábitat de la fauna silvestre y embellecimiento del paisaje (Buitrago-Guillen et al., 2018).
Las especies forrajeras arbóreas y arbustivas presentes en los SSP brindan una serie de interacciones que permiten obtener beneficios adicionales como la regulación térmica y el mejoramiento de las variables fisiológicas-metabólicas relacionadas con el estrés calórico (Barragán-Hernández et al., 2015). Los animales manejados bajo SSP han presentado comportamientos que indican un entorno térmico confortable (Vieira et al., 2021). Además, de una interacción positiva entre los árboles y las pasturas, donde la protección de la radiación solar y del viento, reduce la evapotranspiración y en consecuencia mejora la disponibilidad de agua en el suelo (Bosi et al., 2020).
Estudios con SSP en el piedemonte amazónico, han reportado un incremento hasta del 35 % en la productividad animal por unidad de área (Rivera et al., 2015). Beneficios adicionales radican en la regulación de la temperatura, la cual puede ser de 2 a 3 °C (Murgueitio et al., 2014) inferior en comparación con las praderas sin cobertura, lo que demuestra un efecto positivo sobre el comportamiento ingestivo, el confort térmico y el bienestar animal (Parra-Herrera et al., 2017).
En condiciones tropicales, animales en pastoreo bajo un SSP compuesto por Cynodon nlemfuensis Vanderyst, Megathyrsus maximus (Jacq.) B.K. Simon & S.W.L. Jacobs y arbustos de L. leucocephala en alta densidad, tuvieron una mayor disponibilidad de forraje y confort térmico, con tiempos de rumia y descanso más prolongados, mientras que en un monocultivo de C. nlemfuensis Vanderyst, los animales dedicaron más tiempo a buscar alimento, aún en las horas de mayor temperatura, lo que puede afectar el bienestar y el rendimiento productivo animal en un monocultivo sin presencia de árboles (Améndola et al., 2019).
Los SSP pueden ser una buena alternativa para la producción ganadera sostenible, proporcionan alimento, brindan servicios ecosistémicos y tienden a mejorar el bienestar de los animales (Améndola et al., 2015). Sin embargo, para potencializar las acciones de mitigación del cambio climático por medio de los SSP, es necesario considerar que estos son unos importantes sumideros de carbono (Contreras-Santos et al., 2020), que incrementan su absorción a través de la materia orgánica del suelo y se almacena en los componentes de la biomasa aérea (tronco, ramas, hojas) y raíces de la vegetación asociada (Blanco-Libreros et al., 2015; Hoosbeek et al., 2016).
La acumulación de carbono en la biomasa aérea de un SSP puede llegar a las 8,83 t ha-1 de C, en comparación con un sistema tradicional, que logra acumular 1,88 t ha-1 de C, lo cual demuestra un almacenamiento 4,6 veces superior respecto a los sistemas de manejo convencionales en el piedemonte amazónico colombiano (Villegas et al., 2021).
Estudios realizados en Nicaragua por Hoosbeek et al. (2016), para determinar la absorción de carbono en SSP con G. ulmifolia, demostraron que se pueden almacenar hasta 70 t ha-1 de C en comparación con praderas sin árboles, donde se alcanza un stock de 45 t ha-1 de C. Además, los mismos autores informaron que se mejora la disponibilidad de elementos indispensables como N y P en el suelo, gracias al aporte de hojarasca.
E. poeppigiana se ha utilizado como cultivo de sombra en asocio con producciones de café y cacao (Viguera et al., 2019). Se ha demostrado que en una densidad de 111 árboles por ha, bajo un manejo asociado al cultivo de cacao, puede capturar hasta 12,91 t ha-1 de C (Vera et al., 2019).
Los SSP compuestos por C. plectostachyus (K. Schum.) Pilg., cercas vivas de G. sepium y árboles dispersos de Diphysa robinioides Benth., en el sureste de México, demostraron una acumulación total de 23,38 t ha-1 de C (Nahed-Toral et al., 2013). En contraste, sistemas más estructurados y arreglos multiestratos con especies arbustivas, arbóreas y maderables han registrado un almacén de carbono con valores entre 60,6 y 65,1 t ha-1, lo cual superó entre 58 % y 69 % el stock de carbono (38,3 t ha-1) registrado en la pradera de monocultivo (Contreras-Santos et al., 2020), lo que evidencia el potencial de los SSP en la captura de carbono y mitigación de los GEI.
La implementación de SSP, en suelos poco productivos de la región amazónica puede constituirse en una alternativa para aumentar las reservas de carbono orgánico a una profundidad de 20-30 cm (Olaya-Montes et al., 2021). Este efecto estaría mediado por la inclusión de especies arbóreas y arbustivas que puede ser una alternativa prometedora para convertir el suelo en un importante sumidero de carbono en las regiones tropicales. Sin embargo, hace falta información de especies promisorias, con potencial nutricional como P. discolor y C. fairchildiana como estrategia para reducir las emisiones de CH4 entérico e incrementar la absorción de carbono y de esta manera, facilitar la adaptación de los sistemas ganaderos a los efectos del cambio climático.
Conclusiones
Especies forrajeras como E. poeppigiana, C. fairchildiana, P. discolor y G. ulmifolia, tienen la capacidad de adaptarse a suelos de baja fertilidad y zonas con alta variación climática. Estas especies podrían representar una alternativa factible de uso como fuente de alimento o para aprovechar los beneficios ecosistémicos de su incorporación en las praderas de monocultivos. La valoración agronómica puede servir como base para emitir recomendaciones de implementación e inclusión en arreglos silvopastoriles, que permitan aprovechar la biomasa forrajera para consumo directo, en sistemas de corte y acarreo o como suplemento en ensilajes, sin dejar a un lado el potencial de absorción de carbono que han demostrado estas especies y así convertir los sistemas ganaderos del piedemonte amazónico en sumideros que mitiguen y se adapten de manera eficiente a los efectos del cambio climático.
Se considera pertinente realizar estudios in vitro - in vivo de la producción de metano a fin de analizar las interacciones entre el contenido de metabolitos secundarios y la composición nutricional, ya que en los reportes consultados se evidencia un elevado aporte de nutrientes y aceptable perfil fitoquímico.

