











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introduction
The impact of mycorrhizal symbiosis on plant evolution (van der Heidjen et al., 2015) and functioning is undeniable. It enhaces nutrient uptake capacity (Ortaş & Rafique, 2017), improves tolerance to water deficit (Augé et al., 2015), and provides defense against certain phytopathogens (Pascual, 2016). In addition to these benefits, other reported advantages include enhacements in soil aggregates (Lehman et al., 2017), the establishment of cooperative relationships with various rhizospheric microorganisms, and active participation in nitrogen (N) and carbon (C) cycling (Jansa et al., 2019; Jiang et al., 2021; Parihar et al., 2020).
Arbuscular mycorrhizal symbiosis is prevalent in approximately 84 % of plant species (van der Heidjen et al., 2015), encompassing a majority of crops. This widespread occurrence, along with its diverse ecosystem services, underscores the importance of fully leveraging it in agroecosystems (Rillig et al., 2019). In principle, there are two approaches to enhance arbuscular mycorrhizal functioning: by applying effective inoculants of arbuscular mycorrhizal fungi, AMF (Berruti et al., 2016), or by managing resident mycorrhizae (Bender et al., 2016).
While some cautionary notes have been raised (Hart et al., 2018), the number of publications successful inoculation practices is increasing. Comprehensive summaries of this work have been published (Pellegrino et al., 2015; Rivera et al., 2007; Schütz et al., 2017; Zhang et al., 2019). Criteria for comparing the effectiveness of mycorrhizal inoculants have primarily relied on evaluating responses in crop yield or biomass under suboptimal nutrient supply conditions that permit symbiosis establishment. The response is typically assessed as a relative or absolute index, relative to the performance of the non-inoculated counterpart (Janos, 2007).
While a significant relationship between the frequency of mycorrhizal colonization and crop yield or biomass has not always been observed (Koch et al., 2017; Thirkell et al., 2022), other authors have reported such a connection (Chu et al., 2013; Ortas & Bilgili, 2022; Pellegrino et al., 2015; Rivera et al., 2007). Some authors also incorporate assessments of colonization frequency and intensity as criteria for establishing the efficacy of inoculants (González, 2014; Simó-González et al., 2017).
In the Cuban program focused on the use of mycorrhizal inoculants, the efficacy of three inoculants, each formulated with an specific AMF isolate, was assessed through field experiments conducted under non-sterile conditions on a wide range of crops and soils. Positive responses in yields were observed, with distinct effects between noted for each of the three inoculants. These effects were consistent and depended on the pH-H2O of the soil or substrate in which the crops were grown (Rivera Espinosa et al., 2023). Remarkably, under the same soil conditions, the greatest effectiveness for each of crops was achieved with the same corresponding inoculant.
Certain crops demostrated effective compatibility with specific inoculants beyond the soil conditions where these inoculants are generally effective across diverse crops (González et al., 2015; Rivera et al., 2007). This compatibility, evident not only in terms of high yields but also in mycorrhizal functioning indicators, doesn't preclude these crops from also forming an equally effective symbiosis with the inoculant recommended for the prevailing soil condition.
In recent years, the country has witnessed increasingly positive outcomes with the use of various bioproducts, contributing to high yields and reduce fertilizers application (Dibut Alvarez et al., 2011; Rivera Espinosa et al., 2023). Concurrently, due to Cuba's economic challenges, exacerbated by the COVID-19 pandemic, the import or production of fertilizers is constrained, resulting in low agricultural yields, specially in crops like sweet potato, which currently stands at approximately 9 t ha-1 (Oficina Nacional de Estadísticas e Información, 2022). This situation has propelled the production and adoption of bioproducts in agricultural practices, including the use of mycorrhizal inoculants.
Ipomoea batata (sweet potato) holds sixth position globally in terms of annual production (Food and Agriculture Organization of the United Nations, 2019), and is significant in Cuba for both human an animal nutrition. The country has a successful breeding program with at least 9 commercially used cultivars (Ministerio de Agricultura (MINAG), 2012). Sweet potato is tipically cultivated under irrigation in both the rainy growing season, characterized by higher temperatures, and in the low rainy season.
Sweetpotato is a crop reliant on mycorrhization (Mukhongo et al., 2017; Sakha et al., 2019). In the country, a positive response to mycorrhizal inoculation has been observed, with variations in inoculant effectiveness depending on the soil type in which the crop is cultivated (Ruíz et al., 2012). However, this research was conducted using a single cultivar and with inoculants different from those currently employed in the country.
The impact of intraspecific variability on mycorrhization is commonly manifested through varying degrees of mycorrhizal functioning and response to cultivars inoculation. This variability has been identified in crops such as maize (Chu et al., 2013), clover (Ryan et al., 2016), and rice (Toppo & Maiti, 2017). While some authors report a decrease in the mycorrhization response in modern cultivars (Wang et al., 2022), other find that modern cultivars developed for high yields respond positively to inoculation (Abdelhalim et al., 2019; Bonareri Oruru et al., 2018). However, few publications explore the influence of cultivars on the effectiveness of different inoculants (Joao et al., 2016), and none has been conducted on sweet potato.
Despite the influence of planting date on sweet potato yield (Espinosa-Cuéllar et al., 2019) potentially impacting the inoculation response due to differences in plant requirements, there is no published information on how planting date or growing season affect inoculants effectiveness in this crop. Given the diverse criteria for cultivar utilization, the goal is not to determine which inoculated cultivar achieves the highest yields. Instead, this study aims to determine whether all sweet potato cultivars respond to inoculation and to establish if the effectiveness of inoculants varies among cultivars and planting seasons in order to achieve the greatest benefits from the use of inoculants.
Materials and methods
Experimental conditions
Two experiments were conducted in the areas of the Tubers and Tropical Roots Institute Research (INIVIT), situated at 22°35' N, 80°18' W, and an elevation of 40 m above sea level in the municipality of Santo Domingo, Villa Clara province, Cuba. The experiments were carried out over period 2010-2012, corresponding to the two growing seasons specified by the Technical Instructions (MINAG, 2012). One experiment was conducted during the rainy season (planting in March and harvesting in August), and the other during the low rainy season (planting in October and harvesting in March). Both experiments were performed under non-sterilized soil conditions.
The soil was classified as Eutric Cambisol with carbonates soils by World Reference Base for Soil Resources (2014). It exhibited pH-H20 values ranging from 7.2 to 7.5 (slightly alkaline) (Table 1), high calcium (Ca) content, and moderate levels of exchangeable magnesium (Mg) and potassium (K), as well as available phosphorus (P). Organic matter values were low and associated with degradation processes resulting from continuous soil exploitation. The quantity of mycorrhizal spores was low (1.2 spores/g), likely due to continuous cultivation, high fertilization, and soil degradation processes (Guo et al., 2020). These characteristics are commonly observed in this type of soil under continuous cultivation (Ruíz Martínez et al., 2012; Simó González et al., 2020).
Table 1 Principal initial characteristics of the Eutric Cambisol with carbonates soil (0-20 cm depth) in two growing seasons, in Santo Domingo, Villa Clara, Cuba. 2010-2012.
| Growing season | pH H2O1 | OM2 g kg-1 | P3 | Ca4 | Mg4 | Na4 | K4 | AMF5 spores/g |
| mg kg-1 | cmolc kg-1 | |||||||
| Rainy | 7.5 | 21.5 | 9.47 | 33.9 | 4.4 | 0.37 | 0.66 | 1.2 |
| Low rainy | 7.25 | 21.3 | 9.31 | 29.1 | 4.5 | 0.38 | 0.7 | 1.1 |
Each value is the average of 8 composite samples, four from each year. 1) pH-H2O: pH measured in water; 2) OM: soil organic matter; 3) P: available phosphorus in soil; 4) Ca, Mg, Na, K: exchangeable cations; 5) AMF spores: mycorrhizal spores. Chemical determinations in soil according to Paneque et al. (2010). Spore determinations according to Herrera-Peraza et al. (2004). / Cada valor es promedio de ocho muestras compuestas de suelo, cuatro de cada año. 1) pH-H2O: pH medido en agua; 2) OM: materia orgánica del suelo; 3) P: fósforo disponible en el suelo; 4) Ca, Mg, Na, K: cationes intercambiables; AMF spores: esporas micorrízicas. Las determinaciones químicas en el suelo de acuerdo con Paneque et al. (2010). Las determinaciones de esporas de acuerdo con Herrera-Peraza et al. (2004).
Average temperature and accumulated precipitation in the experimental periods
The mean temperature and accumulated precipitation data for each experimental period, as the 25-year average, are presented in Table 2. The data were obtained from Agrometeorological Station #326 (Cuban Institute of Meteorology), located less than 0.5 km from the experimental area. Precipitation measured daily by a rain gauge (± 0.1 mm), and temperature was recorded hourly with a thermometer (± 0.1 ºC). The results are expressed as accumulated precipitation (mm) and mean daily temperatures (ºC) during the experimental periods.
Table 2 Cumulative precipitation and mean daily temperature during the experimental periods (2010-2012) and 25-year averages. Santo Domingo, Villa Clara, Cuba.
| Precipitation (mm) | Temperature ºC | |||||
| Experimental periods | repetition 1 | repetition 2 | 25 years () | repetition 1 | repetition 2 | 25 years () |
| Rainy | 712.7 | 630.7 | 743.9 | 25.9 | 25.8 | 25.5 |
| Low rainy | 198.3 | 155.2 | 270.6 | 20.7 | 22.3 | 22.4 |
Climatological data were sourced from Agrometeorological Station No. 326, affiliated with the Instituto de Meteorología de Cuba (INSMET), and situated at the Instituto de Investigaciones de Viandas Tropicales (INIVIT) / Los datos de las variables climatológicas se obtuvieron de la Estación Agrometeorológica No. 326, adjunta al Instituto de Meteorología de Cuba (INSMET) y ubicada en el propio Instituto de Investigaciones de Viandas Tropicales (INIVIT).
The rainy period is marked by higher rainfall and temperatures compared to those in the low rainfall period. In this phase, which accounts approximately 73.6 % of the total accumulated rainfall, the rainfall was slightly lower in the second year than in the first repetition, ranging in both years between 86 % and 96 % of the 25-year average (743.9 mm). During the low rainy season, values obtained in both repetitions were below the historical average (54 % to 73 %), with the second repetition being drier (Table 3). Temperatures during the rainy period were similar to each other and to the 25-year average, ranging between 3.5 ºC and 5.2 ºC higher than those in the low rainfall period.
Table 3 Cultivars sweet potato (Ipomoea batata) used in the experiments and some of their characteristics. Santo Domingo, Villa Clara, Cuba. 2010-2012.
| Cultivar | Tuberous root shape | Skin color | Mass color | Growing cycle |
| IX-1 | Round | White | White | long |
| IK-37 | Oblong | Yellow | Beige | long |
| IB-71-9 | Elliptical | Pink | Beige | long |
| IFSB | Oblong | Red | Orange | long |
| IX-18 | Round | Yellow | Yellow | long |
| IA-1517 | Round | White | White | long |
| T-65 | Oblong | Red | Orange | long |
| A-4 | Elliptical | Red | Purple | long |
| INIVIT B 240 | Long Elliptical | Pink | White | medium |
| INIVIT BS-16 | Round Elliptical | Red | Orange | medium |
| INIVIT BS-20 | Elongated | White | Beige | long |
| INIVIT B-9 | Oblong | White | Yellow | medium |
| INIVIT BS-12 | Elliptical | White | White | long |
| INIVIT B 98-2 | Round | White | Beige | medium |
| INIVIT B 98-3 | Round | Pink | Beige | medium |
| INIVIT B2-2005 | Round | Red | Yellow | long |
| CEMSA 78-354 | Round elliptical | White | Beige | medium |
Treatments and experimental design
The effectiveness of three AMF inoculants was evaluated in 17 sweet potato cultivars across two growing seasons. Each experiment followed a randomized block design with a split-plot arrangement and four replicates. The main plots (factor inoculants) considered six levels: three inoculants with 50 % of the recommended doses of nitrogen (N), phosphorus (P), and potassium (K) as per Technical Instructions (MINAG, 2012), and three non-inoculated levels, including a control (without fertilizers), 50 % of the NPK dose, and the recommendation 100 % NPK. The 17 cultivars were assigned to the subplots, and both experiments conducted twice.
The evaluated cultivars are outcomes of the breeding program of the Tubers and Tropical Roots Institute Research (INIVIT), showing variations in phenotypic characteristics (Table 3). Long-cycle cultivars have a harvesting period of around 5 months, while medium-cycle cultivars are tipically harvested between 4 to 5 months. The last nine cultivars are commercially planted in the country (MINAG, 2012), whilethe others are promising. In all instances, these are modern cultivars developed for high yields under optimal doses of mineral fertilizers.
The inoculants were each formulated with a specific AMF isolate. The isolates used for preparation were Glomus cubense (Y. Rodr. & Dalpé) isolate INCAM-4/DAOM241198, Rhizoglomus irregulare ((Błaszk., Wubet, Renker & Buscot) Sieverd., G.A. Silva & Oehl) isolate INCAM-11/DAOM711363, and Funneliformis mosseae ((T.H. Nicolson & Gerd.) C. Walker & A. Schüßler) isolate INCAM-2, all belonging to the collection of the National Institute of Agricultural Sciences. These inoculants have been consistently evaluated since 2003 across different crops and soil types (Rivera Espinosa et al., 2023).
A planting frame of 0.90 x 0.25 m, as per MINAG (2012), was employed, and cuttings of 25-30 cm in length served as planting material. In each subplot, 19 sweet potato rows were arranged, with each row corresponding to a different cultivar and two border rows. The central 7 m of the 8 m furrows were assessed, involving 28 plants of each cultivar.
Preparation and inoculants application
Inoculants were prepared by applying each AMF isolate with a high degree of purity to Urochloa decumbens seeds, which were then grown in a specific substrate following the method outlined by Fernández et al. (2000). After four months, the plants underwent water deficit conditions before cutting, and the substrate enriched with mycorrhizal propagules was extracted. This material, containing a mix of substrate material with mycorrhizal propagules, including mycorrhizal roots with a colonization frequency between 61 % to 68 %, was dried at room temperature in the shade. Spore contents ranged between 80 and 120 spores/g, with variations between the formulations. The quantities of mycelia and infective roots were not determined.
To avoid the possible influence of different spore contents on the comparison of inoculant effectiveness, as well as taking into account the procedure followed in the inoculant comparison programme (Rivera Espinosa et al., 2023), the initial contents were adjusted to the range of 25 to 30 spores/g by dilution and homogenization with sterile substrate. Inoculants were named according to the applied AMF.
Inoculation was carried out using a mixture of 0.125 kg of inoculant per 600 mL of water, covering the lower third of the vegetative seed by immersion (Ruíz Martínez et al., 2012). This application method is equivalent to applying 35 kg ha-1 of inoculant. The inoculated cuttings were left to aerate in the shade for two hours before planting.
Fertilization and cultural attentions
The 100 % NPK dose comprises 90 kg ha-1 of nitrogen, 75 kg ha-1 of P2O5, and 150 kg ha-1 of K2O (MINAG, 2012). The 50 % NPK dose used alongside inoculants aligns with previous findings for this specific crop and soil type, representing the optimal fertilizer quantities required for effective inoculation (Ruiz-Martínez et al., 2012). Fertilizers - urea (46-00), simple superphosphate (0-20-0), and potassium chloride (0-0-60) - were applied 25-30 days after palnting as carriers.
During the low rainfall period, irrigation was conducted by applying 300 m3 ha-1 every seven days up to 45 days, followed by watering every 10 days until 15 days before harvest when irrigations was suspended. In the rainy period, irrigation followed similar criteria, applied when rainfall fell below the stupulated standards for each period. All other farming practice were executed in the line with the Technical Instructions (MINAG, 2012).
Soil sampling and chemical determinations
For soil analysis conducted at the commencement of each experimental cycle, two samples, each composed of 10 subsamples from a depth of 0-20 cm, were obtained for every replicate. This resulted in a total of 8 samples for each yearly repetition of each experiment, amounting to 32 samples oveall. The methods employed for various chemical determinations were as follows: Organic matter determination was conducted using the Walkley-Black method. Phosphorus extraction was carried out through the Machiguín method, employed an extractive solution of (NH4)2CO3 at a concentration of 10 g L-1 and with a pH of 9.0, followed by titration with 0.05 M HCl. Exchangeable cations were extracted using NH4Ac at a concentration of 1 M and at soil to solution ratio of 1:5 at pH 7, and their determination was performed via flame photometry. All methodologies were detailed by Paneque et al. (2010).
Frequency of mycorrhizae colonization of roots (%)
The sampling procedure was conducted in both experiments approximately 90 days after the sweet potato planting. Composite samples of fine roots were collected from six plants within each of the 408 subplots in each yearly repetition (17 cultivars x 6 inoculants and fertilizer levels, and four replicates). From the collected roots weighing 200 mg, the samples were oven-dried at 70 °C until a constant mass was achieved. Subsequently, the roots were stained following the protocol outlined by Phillips & Hayman (1970). The evaluation process was performed using a stereo microscope ain line with the method described by Giovanetti & Mosse (1980).
Mycorrhizae spores evaluation
The evaluations were conducted at the onset of the experiments, utilizing the initial soil sampling, and at the harvest stage. During the latter, composite rhizospheric soil samples were gathered from each cultivar, conprising 6 subsamples (0-20 cm), from all 408 subplots in each yearly repetition. Spore extraction was conducted following outlined by Herrera-Peraza et al. (2004). Spores were enumerated on Doncaster plates using a stereo microscope and reported as spores per 50 g of soil.
Commercial yield (t ha-1) determination
All cultivars were harvested 150 days after planting to ensure uniformity in the experimental plots. The plants within the central 7 m of each cultivar were utilized to determine the yield. The yield was estimated in t ha-1 of commercial roots, weighing more than 115 g.
Data analysis
For statistical analysis, ANOVA was conducted on the experimental data. In cases where the interaction between factors was significant (p≤0.05), subsequent partitioning of the higher-order significant interaction involving inoculants and cultivars was performed to mitigate the influence of varying cultivar productivities. Differences among means were assessed using Duncan's test (p≤0.05) or 95 % confidence intervals. Additionally, linear regression and Pearson correlation analyses were conducted to establish relationships between the frequency of colonization versus yields or AMF spores observed during cultivar inoculation in each growing season. The significance of regression and correlation coefficients was determined usinf t-student tests. Data processing was carried out using the statistical package SSPS version 21.0.
Results
Effectiveness of inoculants on the cultivars yield
In the rainy season, each factor and the interactions between inoculants and cultivars were significant (p≤0.001). There was a positive response (p≤0.05) to mineral fertilization across all cultivars, resulting in the following order: 100 % NPK > 50 % NPK > control (Table 4).
Table 4 Effectiveness of inoculants on average two-year yields (t ha-1) of 17 sweet potato cultivars (Ipomoea batata) in rainy season on Eutric Cambisols soils. Santo Domingo, Villa Clara. 2010-2012. Partitioning of inoculants x cultivars interaction.
| Cultivar | Control | 50% NPK | 100 % NPK | 50 % NPK | ||
| INCAM-4 | INCAM-11 | INCAM-2 | ||||
| IX-1 | 25.93 d | 34.0 c | 37.30 ab | 35.12 bc | 40.47 a | 35.24 bc |
| IK-37 | 20.40 d | 24.5 c | 29.80 b | 29.05 b | 33.70 a | 30.39 b |
| IB-71-9 | 21.54 c | 22.2 c | 27.61 b | 26.55 b | 30.56 a | 25.60 b |
| IFSB | 21.62 c | 26.2 b | 27.47 b | 28.63 b | 32.17 a | 26.82 b |
| IX-18 | 20.03 c | 22.6 b | 23.75 b | 24.36 b | 28.98 a | 24.38 b |
| IA-1517 | 24.11 d | 29.05 c | 31.94 b | 32.93 b | 36.27 a | 28.01 c |
| T-65 | 19.08 c | 21.3 c | 23.63 b | 23.87 b | 28.44 a | 23.80 b |
| A-4 | 20.03 e | 26.2 d | 30.48 ab | 29.13 bc | 32.57 a | 26.38 cd |
| I-BS-16 | 23.79 d | 29.7 c | 33.38 ab | 32.85 ab | 35.02 a | 31.64 bc |
| I-BS-20 | 18.60 c | 26.7 b | 27.10 ab | 28.91 a | 31.33 a | 25.68 b |
| I-B-9 | 18.58 d | 21.5 c | 25.12 b | 25.40 ab | 28.13 a | 22.89 bc |
| I-BS-12 | 18.39 d | 23.5 c | 26.72 b | 25.78 bc | 31.61 a | 26.10 bc |
| I-B-240 | 22.95 d | 26.5 c | 30.00 ab | 28.87 bc | 32.10 a | 27.85 c |
| I-B-98-2 | 21.35 c | 26.6 b | 30.86 a | 29.39 a | 31.86 a | 26.50b |
| I-B-98-3 | 20.45 d | 23.4 c | 30.39 a | 26.17 bc | 32.39 a | 26.65 b |
| I-B2-2005 | 23.78 e | 27.0 d | 33.59 ab | 31.13 bc | 34.92 a | 30.34 c |
| C-78-354 | 23.12 d | 29.6 c | 32.64 b | 32.98 b | 36.04 a | 30.85 bc |
| Es A x B | 0.731*** | |||||
Different letters in the same row (cultivar) lead to significant differences at p≤0.05 between the inoculant factor levels by Duncan Test. 100 % NPK means 90, 75 and 150 kg ha-1 of N, P2O5 and K2O respectively. / Letras diferentes en la misma fila (cultivar) conllevan a diferencias significativas a p≤0,05 entre los niveles del factor inoculantes mediante Prueba de Duncan. 100 % NPK significa 90, 75 y 150 kg ha-1 de N, P2O5 y K2O respectivamente.
Differentiated responses to inoculation were observed among cultivars, wherein INCAM-11 consistently yielded the highest results in each of them. In nine cultivars, INCAM-11 led to significantly higher yields (p≤0.05) compared to those obtained with 100 % NPK; for the remaining cultivars, it produced similar results, always surpassing the 50 % NPK treatment.
Moreover, INCAM-11 application resulted in significantly higher yields (p≤0.05) in thirteen cultivars (76.4 %) compared to INCAM-4, and across all cultivars, it consistently outperformed INCAM-2. While cultivars displayed varying yields (Table 4), generally, yields from INCAM-11 inoculation ranged between 28 and 36 t ha-1, averaging 32.7 t ha-1. Notably, only cultivar IX-1 exhibited yields close to 40 t ha-1.
In the low rainy growing season (Table 5), cultivars showed a similar response to inoculation, and significant effects of different factors were observed (p≤0.001), including significant second-order interaction between inoculant and cultivar. The non-inoculated treatments demonstrated an increase response in yield with fertilizer application, with yields ranking (p≤0.05) as follows: 100 % NPK > 50 % NPK > control.
Table 5 Effectiveness of inoculants on two-year average yield (t ha-1) of 17 sweet potato cultivars (Ipomoea batata) grown in the low rainy season on Eutric Cambisol soils of Santo Domingo, Villa Clara. 2010-2012. Partition of inoculants x cultivars interaction.
| Cultivar | Control | 50 % NPK | 100 % NPK | 50 % NPK | ||
| INCAM-4 | INCAM-11 | INCAM-2 | ||||
| IX-1 | 22.83 c | 28.49 b | 32.80 a | 30.93 a | 32.31 a | 28.60 b |
| IK-37 | 20.16 d | 23.15 c | 28.78 ab | 27.24 ab | 29.29 a | 26.78 b |
| IB-71-9 | 18.31 d | 21.25 c | 25.36 a | 23.64 ab | 25.19 a | 22.29 bc |
| IFSB | 21.06 c | 22.30 b | 25.88 a | 23.67ab | 25.24 a | 22.52 b |
| IX-18 | 19.15 b | 19.61 b | 21.54 a | 21.52 a | 22.07 a | 20.85 ab |
| IA-1517 | 23.16 c | 26.16 b | 29.07 a | 27.89 ab | 29.80 a | 26.55 b |
| T-65 | 16.66 c | 19.37 b | 21.86 a | 20.81a b | 21.29 ab | 19.56 b |
| A-4 | 17.25 c | 21.40 b | 23.84 a | 22.45 ab | 23.78 a | 21.13 b |
| I-B-240 | 20.02 c | 22.79 b | 25.90 a | 24.32 ab | 26.14 a | 23.38 b |
| I-BS-16 | 21.71 d | 25.05 c | 29.12 a | 28.06 ab | 29.58 a | 26.68 bc |
| I-BS-20 | 21.09 d | 24.57 c | 28.88 a | 27.10 ab | 29.09 a | 25.84 bc |
| I-B-9 | 17.61 b | 18.25 b | 21.31 a | 20.39 ab | 21.97 a | 19.84 ab |
| I-BS-12 | 19.45 c | 21.71 c | 26.08 a | 24.83 ab | 26.34 a | 22.97 bc |
| I-B-98-2 | 18.33 c | 24.00 b | 27.75 a | 25.62 ab | 27.87 a | 24.16 b |
| I-B-98-3 | 18.12 c | 20.05 b | 24.55 a | 23.72 a | 24.63 a | 22.84 a |
| I-B2-2005 | 22.23 c | 24.09 bc | 27.58 a | 26.06 ab | 27.25 a | 24.69 b |
| C-78-354 | 17.58 d | 22.07 c | 25.91 a | 24.69 ab | 26.29 a | 23.78 bc |
| Es A x B = 0.61*** | ||||||
Different letters in the same row lead to significant differences at p≤0.05 between levels of the inoculant factor for each cultivar by Duncan Test. 100 % NPK means 90, 75 and 150 kg ha-1 of N. P2O5 and K2O, respectively. / Letras diferentes en la misma fila conllevan a diferencias significativas a p≤0,05 entre los niveles del factor inoculante para cada cultivar mediante Prueba de Duncan. 100 % NPK significa 90, 75 y 150 kg ha-1 de N, P2O5 y K2O, respectivamente.
A positive and distinct response to inoculation was observed, consistently resulting in the highest yields for each cultivar with the application of INCAM-11. In all cases, these yields were statistically similar (p≤0.05) to those obtained with the 100 % NPK dose and significantly higher (p≤0.05) than the yields from the non-inoculated treatments. Yields ranged between 22 and 32 t ha-1, with an average of 26 t ha-1, lower than that achieved in the rainy season. While the inoculation of INCAM-11 did not significantly surpass the yield obtained with INCAM-4, it yielded significantly higher results (p≤0.05) than INCAM-2 in 14 out of 17 cultivars.
Effectiveness of inoculants on the cultivars colonization frequencies
In both growing seasons, significant effects of the different factors were observed (p≤0.001), and the interactions between inoculants and cultivars were also significant (p≤0.05). However, in both periods, the cultivars inoculated with INCAM-11 consistently achieved higher colonization frequencies (p≤0.05) compare to the values obtained when applying the other two inoculants (Figure 1 A and B), as well as with the non-inoculated fertilizer levels.
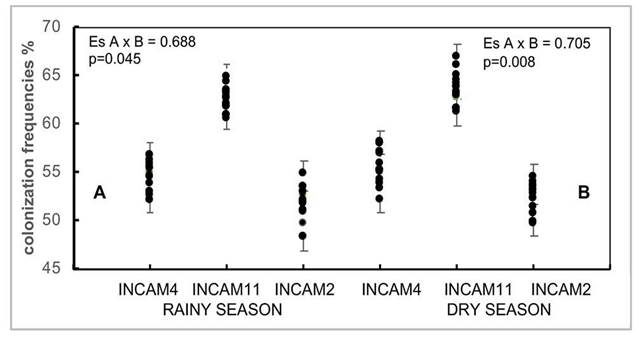
Figure 1 Effectiveness of inoculants on colonization frequency (%) of 17 cultivars of sweet potato (Ipomoea batata) grown on Eutric Cambisol soils. Santo Domingo, Villa Clara (average values for the two years, 2010-2012).
Colonization frequencies when INCAM-11 was applied ranged between 60.8 and 64.2 % in rainy period, and between 61.1 % and 66.9 % in low rainy period, showing differences (p≤0.05) in each period among some cultivars, but not between growing periods. When INCAM-4 and INCAM-2 were inoculated, the colonization frequencies veried between 52.6 % - 56.8 % and 48.3 % - 54.9 % in the rainy period, and between 52.1 % - 57.9 % and 49.6 % - 54.5 % in the low rainy period respectively, showing some differences (p≤0.05) between cultivars in each period, but not between growing periods. Non-inoculated cultivars presented the lowest values of colonization frequency (p≤0.05), ranging from 9 % and 18 %.
Significant correlations (p≤0.001) were observed in both growing seasons between the colonization frequencies of the cultivars inoculated with the three inoculants and their respective yields. The determination coefficients (r2) were 0.49 in the rainy season and 0.36 in the low rainy season (dashed lines in Figure 2 A and B), indicating that higher colonization frequencies were associated with higher yields.
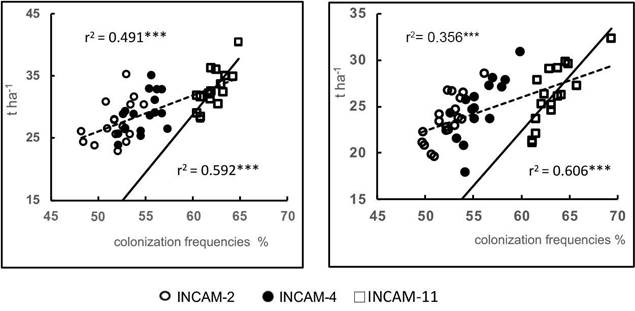
Figure 2 Relationships between colonization frequencies and yields when applying inoculants to sweet potato cultivars (Ipomoea batata) in Eutric Cambisols soils of Santo Domingo, Villa Clara (averaged values for the years 2010-2012).
In both seasons, higher colonization frequencies were consistently observed when inoculating any of the cultivars was inoculated with INCAM-11, compared to those obtained with the other two inoculants. Moreover, when considering only the inoculation with INCAM-11, the determination coefficients (r2) increased to aprproximately 0.6 (p≤ 0.001) in both growth periods (solid lines in Figure 2 A and 2 B). These values were notably higher than the determination coefficients obtained when analyzing the results of each of the other inoculants individually, which ranged from 0.2 to 28 (not significant) in both periods.
Effectiveness of inoculants on the mycorrhizal spores
In both growing season, significant effects were observed for various factors (p≤0.001), including significant interactions between inoculants and cultivars (p≤0.01). However, the inoculation with INCAM-11 consistently led to higher mycorrhizal spore counts in each of the cultivars (p≤0.05), compared to the values obtained with the other inoculants (Figure 3 A and B) and with the non-inoculated fertilizer levels.
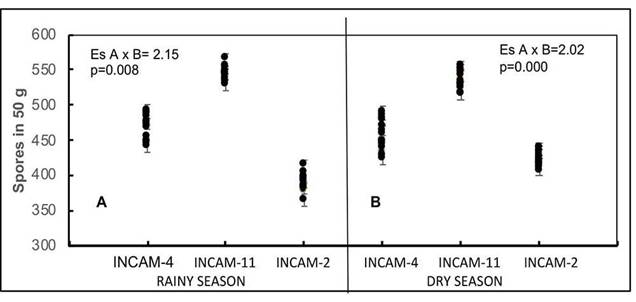
Figure 3 Effectiveness of inoculants on mycorrhizal spores of 17 cultivars of sweet potato (Ipomoea batata) grown on Eutric Cambisol soils of Santo Domingo, Villa Clara. 2010-2012.
The spores obtained with INCAM-11 inoculation ranged between 528 and 566 spores in 50 g during the rainy period and 517 to 551 in the low rainy period, showcasing differences (p≤0.05) among some cultivars in each period, with certain cultivars exhibiting higher values in the rainy period. For INCAM-4 and INCAM-2, mycorrhizal spores varied from 423 to 491 and 365 to 436, respectively. INCAM-4 induced higher spore amounts (p≤0.05) in certain cultivars compare to INCAM-2. Non-inoculated cultivars had the lowest mycorrhizal spore values (p≤0.05), ranging from 68 to 116 spores in 50 g.
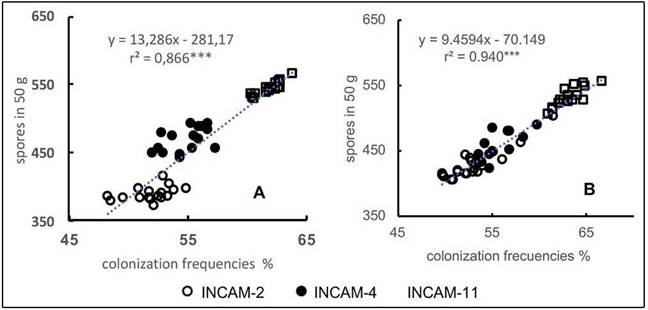
Despite differences in cultivar responses to the three inoculants, a significant direct correlation was noted between colonization frequencies and spores reproduced by inoculated cultivars in both growing seasons (Figure 4). This illustrates that higher colonization frequency corresponded to increased spore reproduction. INCAM-11 consistently resulted in higher values of both fungal variables across all cultivars in both periods under these specific soil conditions.
Discussion
In the rainy season, characterized by higher yields and increased nutritional demands (Espinosa-Cuéllar et al., 2019), the consistently superior response to INCAM-11 inoculation in terms of yield, colonization frequency, and spore counts highlights the effectiveness of this inoculant under these conditions. Moreover, the strong corelation between colonization frequency and yield suggests that the enhaced mycorrhizal performance resulting from INCAM-11 inoculation is directly linked to higher yields, consistent with findings in other crops using similar soil types (González, 2014; Simó González et al., 2017). Overall, the evaluation underlines the superior efficacy of INCAM-11 during this planting period for any of the sweet potato cultivars evaluated.
Differences between the inoculants can be explained in line with Kiers et al. (2011), wherein the higher efficiency of the applied AMF inoculant increased the mutual benefits between the plant and the fungus. Thus, plant growth and yield are increased, associated with increased nutrient and water supply by the mycorrhizae (Augé et al., 2015; Ortas & Rafique, 2017) and in parallel mycorrhizal structures are augmented by the increased supply of carbohydrates and lipids from the plant (Lanfranco et al., 2018). Among the structures that benefit are hyphae, arbuscules and also spores (Willys et al., 2013). The substantial relationship observed between colonization frequency and spore count, with the highest values for both variables with INCAM-11 application, reinforces this underlying mechanism.
Another crucial factor to consider is the plant species itself; sweet potato, being a mycotrophic species with considerable mycorrhizal reliance (Ruiz Martínez et al., 2012), does showcase differences in inoculant effectiveness, albeit less pronounced compares to other crops like Colocasia esculenta and even Manihot esculenta (Rivera et al., 2007). This interplay of factors seems to elucidate than during the season with lower yields, no significant differences were observed among cultivars when inoculated with either INCAM-11 and INCAM-4. In species such as Urochloa, such distinctions in yield effectiveness between these inoculants persist consistently throughout the year, even with more pronounced seasonality in biomass production (González, 2014).
While there were significant differences in colonization frequencies among certain cultivars inoculated with INCAM-11, all instances of colonizations consistently fell within the range of 60 % and 68 %. However, in other crops, studies involving a broader spectrum of cultivars, including modern, traditional, and non-domesticated species, have reported a wider variation in colonization frequencies, ranging from 20 % to 70 % (Chu et al., 2013, Ryan et al., 2016). This disparity may be attributed to several factors, including the homogeneity of the cultivars in the present study obtained under a unified breeding scheme, that uniform application of an effective inoculant across all cultivars, the nutrient supply in these experiments meeting the plants' requirements of the plants, and the inherent mycotrophy of sweet potatoes, which might contribute to this observed behavior.
The significant correlations obtained in both growth periods between colonization frequency and yields when applying the three inoculants not only suggest that yield were dependent on the differentiated colonization caused by the inoculants but also underscore the efficacy of this indicator in evaluating inoculants effectiveness. Similarly, various studies employing these same inoculants have reported significant correlations between mycorrhizal colonization and yield or biomass production in different crops (González, 2014; Simó González et al., 2017). The higher correlation coefficients observed when considering only the application of INCAM-11 indicate that the more effective the inoculant, the stronger the connection between mycorrhizal performance and yield, alingning with the findings of Kiers et al. (2011).
When considering spores, whose reproduction is characteristic of each crop (Willis et al., 2013), comparing quantities across different crop experiments isn't valid. Spore reproduction is a vital part of the fungus's survival mechanism and tipically increases toward the end of the crop cycle or in response to adverse mycorrhizal conditions (van der Heidjen et al., 2015). Regardless of these behaviours, it seems that, at least with these specific inoculants, the effectiveness is also reflected in the quantities of spores reproduced, which correspond to the colonization frequencies obtained.
In line with the findings on this study, Zangaro and Lirio Rondina (2016), studying 27 tropical herbaceous plants and shrubs, also observed strong relationships between colonization and spore numbers, considering both as indicators of mycorrhizal functioning. Similarly, Koch et al. (2017) discovered significant correlation between spore counts and yield while evaluating 56 AMF isolates across three plant species. González (2014), working with similar inoculants in Urochloa fodder areas, reported substantial relationships between colonization and spore production, with the highest values observed when applying the effective inoculant suitable for the soil type. Additionally, higher f spore counts were associated with prolonged persistence of the applied inoculant, delaying the necessity for re-inoculation.
In the present experiments during the high-yield season, only one cultivar (INIVIT B 98-2), across both repetitions, achieved similar yields when inoculated with either INCAM-11 or INCAM-4. However, even in this cultivar, the percentages of colonization and spore counts were consistently higher when inoculated with INCAM11. Moreover, in the low rainy and cold season, while there were no significant differences in yields between the two isolates, the frequencies of colonization and spore counts were notably higher in all cases when INCAM-11 was applied. Therefore, it doesn't seem that the phenomenon of compatibility (Rivera et al., 2007) is evident in any of these cultivars with the studied inoculants.
The results underscore the superior effectiveness of the INCAM-11-based inoculant for sweet potato cultivars in these soils. It's noteworthy that the benefits of inoculation were achieved by reducing NPK fertilizer doses by 50 %, attributable to the increased nutrient uptake in efficiently mycorrhizal crops (Gonzalez et al., 2015; Rivera et al., 2007; Ruiz Martinez et al., 2012;). Additionally, it highligts the potential use of the previous crop inoculated with INCAM-11 (Espinosa-Cuéllar et al., 2019) as a mean to efficiently mycorrhizae any of the studied sweet potato cultivars in succession and reap the benefits of effective mycorrhization in both crops.
Various authors who have compared the efficacy of these same inoculants across different crops such as grass species, fodder legumes, cassava, canavalia, and banana, have consistently observed that in soils with a pH-H2O ranging between 7-8, INCAM-11 exhibits the highest effectiveness, followed by INCAM-4, while INCAM-2 generally displays the lowest efficacy (González et al., 2015; González Cañizares et al., 2016; Joao et al., 2016; Rivera Espinosa et al., 2023; Simó González et al., 2017). Hence, sweet potatoes cultivated in soils at pH-H2O ranging between 7.25 to 7.55 fall within the scope of crops where this pattern holds true.
Conclusion
The sweet potato cultivars developed through the breeding program displayed a positive response to inoculation, even though they were initially selected for high yields with optimal fertilizer supply. Across different cultivars and growing seasons, INCAM-11 consistently showcase the most pronunced effects. This was evident not only in commercial yield but also in terms of colonization frequency and mycorrhizal spore quantity. The varyng levels of colonization frequency and spore production served as a clear indicators of the effectiveness among the different inoculants.
The promising effectiveness of INCAM-11 inoculant across these sweet potato cultivars, particularly in Eutric Cambisol soils with carbonates, offers a potential avenue for simplifying management of AMF inoculants for this crop under similar soil conditions. Further validation at a larger scale in actual production setting is highly recommended.

