










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Mesoamericana
On-line version ISSN 2215-3608Print version ISSN 1659-1321
Agron. Mesoam vol.24 n.2 San Pedro Jul./Dec. 2013
Información Técnica
Síntomas asociados a la deficiencia de boro en la palma aceitera (Elaeis guineensis JACQ.) en Costa Rica
Symptoms associated to boron deficiency in the oil palm (Elaeis guineensis JACQ.) in Costa Rica
Dirección de Correspondencia Symptoms associated to boron deficiency in the oil palm (Elaeis guineensis JACQ.) in Costa Rica
Resumen
El objetivo de este trabajo es brindar información fisiológica relativa a las funciones, la disponibilidad, la economía y la deficiencia de boro en las plantas, con énfasis en la palma aceitera cultivada en Coto, Costa Rica. Se exponen las generalidades sobre la química del boro en el suelo y en las plantas, y se discuten sus posibles funciones estructurales, metabólicas y morfogenéticas. Se resalta su papel en la formación y la estabilidad de la pared celular, el transporte de azúcares, el control de los niveles de auxinas y en el desarrollo reproductivo. Se compara la susceptibilidad relativa de las hojas y las raíces a la deficiencia de boro. Se describen los síntomas de la deficiencia de boro observados en la palma aceitera, los posibles mecanismos causales y los métodos apropiados para su diagnóstico. Se discuten además las terapias, formulaciones químicas y métodos agronómicos disponibles para su corrección.
Palabras clave: absorción y transporte de iones, diagnóstico deficiencia de boro, corrección deficiencia de boro.
Abstract
The objective of this work is to summarize the physiological and eco-physiological information relative to the functions, availability, economy and deficiency of boron in plants, with emphasis on the oil palm growing in Coto, Costa Rica. General aspects of boron chemistry in soils and plants are outlined, and its possible structural, metabolic and morphogenetic roles are discussed. The roles of boron in plant cell wall formation and stability, sugar transport, control of auxin levels, and reproductive development are discussed. The relative susceptibility of leaves and roots to boron deficiency are also compared. Boron deficiency symptoms observed in the oil palm are discussed, the possible causal mechanisms are outlined, and appropriate methods for deficiency corrections are mentioned.Therapies and chemical formulations and agronomic practices available for correcting boron deficiency and grant proper boron nutrition are presented.
Keywords: ion uptake and transport, boron defficiency diagnostic, boron defficiency correction.
Introducción
La esencialidad del boro en la nutrición mineral de las plantas superiores, fue establecida mediante métodos fisiológicos por Warington (1923) utilizando Vicia faba, cebada, frijol común y centeno, y Sommer y Lipman (1926) en girasol, algodón, higuerilla y mostaza, entre otras especies. Esto se dio gracias a avances técnicos como la producción de frascos de borosilicato para el cultivo de plantas en soluciones nutritivas asépticas (Sommer y Lipman 1926). En la actualidad, se dispone de métodos fisicoquímicos basados en la combinación de la ionización con plasma y la espectrometría de masas (ICP-MS) para determinar la concentración de boro en los tejidos de las plantas y en diversos sustratos agrícolas (Sah y Brown 1997); la determinación del contenido de boro del suelo es aún problemática, por lo que a menudo es excluido del análisis convencional.
El boro es un mineral necesario para que las plantas completen su ciclo de vida, participa en funciones metabólicas y estructurales en las cuales no puede ser sustituido, las plantas severamente privadas de boro exhiben anormalidades en su crecimiento, desarrollo y reproducción, y su deficiencia se asocia a síntomas específicos que sólo pueden ser corregidos con la aplicación de este elemento (Rains 1976, Frieden 1984, Welch 1995). La concentración total de boro en el suelo es de 20-200 mg/kg, la mayoría del cual es inaccesible para las plantas. Su disponibilidad está controlada mayormente por el pH, la textura y la materia orgánica del sustrato (Daroub y Snyder 2007).
La concentración de boro en las hojas de las plantas superiores fluctúa dentro de un reducido rango alrededor de los 20 mg/kg (2 umol/g peso seco), lo que sugiere un delicado balance entre la suficiencia y la toxicidad (Kirkby 2012). El boro produce numerosos efectos benéficos en la salud humana y podría constituir un nutriente esencial de nuestra dieta (Tinker 1981, Nielsen 1997). En las plantas, la distribución del boro entre diferentes compartimentos celulares depende de la especie, el genotipo, el suministro del elemento y el estatus nutricional, pero se encuentra en todos los componentes celulares principales. El mayor contenido de boro parece localizarse en las paredes celulares e intracelularmente, la fracción citoplasmática parece ser mayor que la vacuolar (Dannel et al. 2002). Tanto la deficiencia como la toxicidad del boro causan grandes pérdidas en los cultivos alrededor del mundo (Robinson 1993, Nable et al. 1997, Lӓuchli 2002), y numerosos trastornos fisiológicos como el corazón negro de las brassicas (coliflor, brócoli, Shelp et al. 1992), las hojas en palmilla del café (Carvajal 1972), deformaciones de frutos como la manzana (Wӧjcik et al. 1999) y la papaya (Wang y Ko 1974), y la necrosis de los bordes de algunas hojas. La gutación ha sido implicada en los mecanismos por los que las plantas regulan la concentración de boro en las hojas, en este caso excretándolo en la savia exudada (Shapira et al. 2012).
Aspectos fisiológicos de la nutrición por boro
El boro se absorbe del suelo en la forma de ácido bórico (B(OH)3) o borato (B(OH)4-) a pH neutro y alcalino (Raven 1980, Lӓuchli 2002), lo que lo hace uno de los dos nutrientes esenciales que existen en la solución del suelo como una molécula no disociada. Las plantas responden directamente al boro soluble (no al adsorbido, Goldberg 1997), cuya disponibilidad en la solución del suelo es mayor a pH 5-7, decrece a pH 7,5-8,5, y aumenta de nuevo a pH superior a 8,75. A bajas concentraciones en la solución nutritiva (<1 uM), el boro parece ser absorbido por transporte activo, pero a concentraciones mayores se mueve por difusión (pasiva) a través de la bicapa lipídica (Dannel et al. 2002); se ha reportado también el transporte a través de los canales de las acuaporinas de las membranas (Hu y Brown 1997) y otros canales específicos para la absorción de boro (Takano et al. 2008). Una vez absorbido, no sufre cambios de valencia en el interior de la planta (Bennett 1996), y su distribución depende de procesos de difusión pasiva, transporte activo, y del acomplejamiento con dioles cis de los carbohidratos. Se reportan importantes interacciones entre el boro, el calcio y el fósforo. Las plantas dicotiledóneas parecen contener mayor concentración de boro que las monocotiledóneas (Bennett 1996), como la palma aceitera y el cocotero. No parece ser un elemento esencial para algunas plantas acuáticas (hidrófitas), que no requieren de la lignina para el soporte mecánico de sus órganos flotantes (Lewis 1980b).
El boro ha sido implicado en importantes procesos estructurales, metabólicos y morfogenéticos como el control del tamaño de los poros de las paredes celulares, el transporte por el floema, la formación de complejos con los azúcares y algunos carbohidratos de la pared celular (Matoh 1997, Blevins y Lukaszewski 1998, Lӓuchli 2002), y la germinación del polen (Lewis 1980a). Desde el punto de vista estructural, parece ser un constituyente esencial tanto de las membranas como de las paredes celulares de las plantas. En las membranas, podría formar complejos con varios constituyentes (fosfolípidos, glicoproteínas, azúcares), y en las células de las raíces causa híper-polarización, lo cual antecede los aumentos de la respiración y la síntesis de ATP, requeridos para la absorción y el transporte de iones (Lӓuchli 2002).
También se ha implicado al boro en el control de la síntesis de lignina y de la diferenciación del xilema, a través de sus funciones en el metabolismo de los fenoles y en el control de los niveles endógenos de auxinas (AIA), por lo que afecta varios eventos celulares (elongación, desarrollo), así como la fortaleza estructural de los tejidos (Dutta y McIlrath 1964). Los perfiles de los aminoácidos se alteran como consecuencia de la deficiencia de boro, especialmente el de aquellos involucrados en las respuestas al estrés y en la señalización, en patrones específicos para cada órgano (Alves et al. 2011). Parece antagonizar la acción de la catecol oxidasa, la polifenol oxidasa, la tirosinasa, las peroxidasas y la AIA oxidasa. Muchos otros sistemas enzimáticos están regulados por los niveles internos de boro (Rains 1976, Frieden 1984, Welch 1995).
El boro está involucrado en el transporte de azúcares a través de las membranas, controla el balance entre estos y la formación de almidón, y afecta la transpiración en las hojas (Bennett 1996). Forma complejos estables con los oxígenos libres y con los grupos cis-hidroxilo de los polisacáridos y los azúcares simples, reduce su polaridad, y facilita su traslado a través de las membranas durante la carga, la translocación y la descarga del floema, lo que le confiere alta movilidad en especies con alto contenido de polyoles en el floema (manitol, sorbitol), como cebolla, apio, culantro, espárragos, brassicas, manzana, melocotón, café, zanahoria, frijol común, aguacate, arveja y olivo, entre otros (Campbell et al. 1975, Brown y Shelp 1997). Interviene además en el metabolismo de los ácidos nucleicos (especialmente del ARN), la síntesis de giberelinas, en la actividad de la alfa amilasa durante la germinación de algunas semillas (avena), y en las síntesis de las bases pirimídicas y de ciertos flavonoides (otra clase de fenoles).
Eco-fisiología de la nutrición por boro
El transporte de boro en el xilema es proporcional al flujo de savia en la corriente de transpiración (Raven 1980). Esto es una consecuencia del funcionamiento del sistema continuo suelo-planta-atmósfera, mediado por la arquitectura hidráulica y el área foliar de las plantas. Una vez alcanzadas las terminaciones de las venas, su redistribución puede ser muy limitada. Sin embargo, como se indicó antes, su concentración en la planta completa depende también de su translocación en el floema y de su demanda por los diferentes sumideros (Brown y Shelp 1997). Esto requiere de un “hábil manejo” de la economía del boro por parte de las plantas, pues a pesar de la gran demanda de boro y nutrientes del floema impuesta por los sumideros, el peligro de la toxicidad debe ser neutralizado manteniendo su concentración por debajo de los requerimientos potenciales (Raven 1980). Esto puede ser especialmente importante en la fertilización precisa de los cultivares compactos y las variedades modernas, precoces y altamente productivas de palma aceitera, que mantienen abundantes sumideros durante todo el año. El boro es de relevancia en el control de las relaciones planta-patógeno debido a su importancia en la formación de la pared celular, en la economía y la partición de los azúcares y otros nutrientes por el floema, en la producción de fenoles y otros metabolitos secundarios como las fitoalexinas (Lewis 1980a), y el control de la relación raíz:tallo (Stangoulis y Graham 2007).
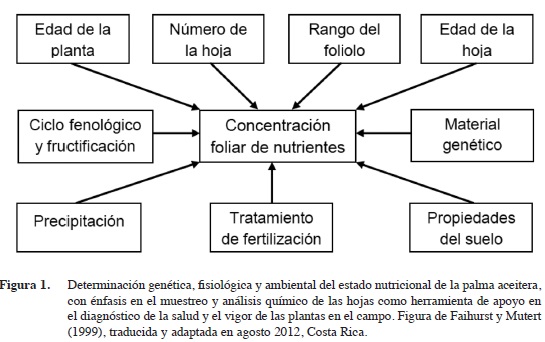
En la palma aceitera y otras especies cultivadas, el estado nutricional se puede analizar como el resultado de una compleja interacción de factores genéticos, fisiológicos y agro-ecológicos, que operan a varias escalas (suelo, plantas, atmósfera), y que tiene importantes componentes temporales (la edad de las hojas, el ciclo fenológico de las plantas, la variabilidad climática) (Figura 1) (Chapman y Gray 1949, Smith 1962, Faihurst y Mutert 1999).
La concentración de boro en las hojas de las plantas superiores fluctúa alrededor de 20 mg/kg (2 umol/g peso seco), y en la palma aceitera, concentraciones foliares entre 15 y 25 mg/kg se consideran adecuadas (Fairhurst y Härdter 2003). Ramírez y Rodríguez4 señalan que en un clon compacto y otro vigoroso muestreados en Coto y en Quepos, Costa Rica, concentraciones foliares de boro de 21-22 mg/kg son consideradas óptimas. Debido a su baja movilidad del boro en muchas especies (Smith 1962), su concentración aumenta con la edad de la hoja, junto con la de otros elementos poco móviles como el calcio y el hierro.
En la palma aceitera, la concentración de boro aumenta con la edad de las hojas, y las puntas y los márgenes de los foliolos contienen más boro. Dentro de una hoja, las hojuelas superiores contienen más boro que las inferiores, y las que apuntan hacia arriba contienen más que las que apuntan hacia abajo (Figura 1). La concentración de boro en las hojas de la palma aceitera varía durante el día, en función de la fotosíntesis y transpiración, y a lo largo de los ciclos climáticos y fenológicos de las plantas, y su acumulación puede estar punteada por periodos de aparente pérdida del elemento (Rajaratnam 1972). Esto implica que el diagnóstico del estado nutricional de una palma aceitera a través del análisis mineral de los tejidos, debe ser realizado preferiblemente mediante el muestreo de hojas jóvenes.
En la palma aceitera y otras plantas monopodiales con arquitectura relativamente simple, y un solo meristemo apical que genera todas las hojas nuevas, es recomendable muestrear las hojas más jóvenes completamente expandidas (5 a 11) (Bell 1997). En contraste con la hoja 17, estas son las mejor ilumina- das, poseen la mayor cantidad de nitrógeno (elemento móvil muy dinámico), y realizan las mayores tasas de intercambio gaseoso y de ganancia fotosintética diaria. El contenido de boro de una palma aceitera adulta es de aproximadamente 7-8,5 mg B/kg de materia seca. En una hectárea de palmas aceiteras adultas, 0,5 kg de boro están contenidos en la biomasa aérea, y 0,07 kg son removidos por los racimos cosechados (Fairhurst y Härdter 2003).
Determinación del cojntenido de boro en hojas con síntomas de deficiencia
El trabajo de campo y la documentación foto gráfica se realizaron en fincas de palma aceitera de ASD-Costa Rica, en Coto, Puntarenas, Costa Rica. Se obtuvo evidencia fotográfica de los síntomas asociados a la deficiencia de boro en palmas productivas, cuyos registros de análisis foliares indicaron la presencia del elemento en concentraciones deficientes. Se muestrea- ron y se analizó el contenido de boro de las hojas 3, 9 y 17 de palmas aceiteras adultas y productivas, en los laboratorios del Centro de Investigaciones Agronómicas de la Universidad de Costa Rica (UCR). Las observaciones de campo y el muestreo foliar fueron realizados el 28 de julio de 2012, durante la transición de la estación seca a la lluviosa en Coto 49, y en Coto 54, Costa Rica, en junio del 2012 y febrero del 2013. La literatura señala que la concentración óptima en palmas adultas debe ser de 15-25 umol/g peso seco (Fairhurst y Härdter 2003).
Deficiencia de boro en la palma aceitera
Las palmas aceiteras son muy sensibles a las bajas concentraciones de boro, por lo que rápidamente muestran síntomas de deficiencia en las hojas, las raíces y las plantas completas (Figuras 2 y 3); estos síntomas han sido descritos parcialmente para otras especies de palmas grandes como la palmera real y el cocotero (Broschat 2005, Patnude y Nelson 2012).
La deficiencia de boro es el desorden nutricional más común en las plantaciones de palma aceitera alrededor del mundo (Fairhurst y Härdter 2003), y se agrava en regiones con suelos alcalinos, arenosos y sujetos a alta precipitación, y en materiales genéticos altamente productivos bien abastecidos con otros nutrientes (N, P, K), cuyo crecimiento vegetativo y el desarrollo continuo de múltiples sumideros imponen altas demandas de boro a las plantas y al suelo de manera permanente. La deficiencia severa inhibe completamente el desarrollo de las hojas nuevas y culmina con la formación de un cráter alrededor del meristemo apical, debido a la desintegración de los primordios foliares no expandidos (Figura 2) (Fairhurst y Härdter 1972).
Las hojas muestran expansión irregular y malformaciones especialmente en la sección distal. Debido a la poca movilidad del boro en la palma aceitera, los síntomas aparecen primero en las zonas en crecimiento, y los foliolos en la sección distal de las hojas son los más afectados. Con frecuencia se observa coloración verde más intensa en las hojas sintomáticas, que se han descrito como acartuchadas, acordeonadas, arrugadas, cola de pescado, hojas ciegas, y hoja chongas, entre otros términos. Además de las malformaciones, las pinnas son tiesas, frágiles y quebradizas (Figura 2).
La deficiencia de boro parece afectar más severamente el crecimiento reproductivo que el vegetativo, tal vez porque se reprimen la síntesis de citoquininas y la germinación del polen, cuyo tubo polínico emerge con malformaciones que le impiden alcanzar el saco embrionario (Lewis 1980a). El rendimiento también puede disminuir debido a la fecundación insuficiente, causada por la pobre germinación del polen y las malformaciones de los tubos polínicos. Los efectos post-fertilización pueden incluir embriogénesis anormal, aborto de semillas y frutos y producción de embriones y frutos deformes. La variedad y severidad de las consecuencias difiere entre especies y dentro de una misma especie, con la localidad, las condiciones agro-ecológicas y a través de los años (Dell y Huang 1997).
En las raíces, la deficiencia de boro retarda el crecimiento meristemático (Figura 3) y causa niveles supra-óptimos de auxinas, que pueden inhibir la división celular e inducir aumentos de la AIA-oxidasa, y la producción de raíces con anormalidades morfológicas, apariencia achatada (Bohnsack y Albert 1977, Hirsch y Torrey 1980), corchosis y niveles reducidos de antioxidantes como el ácido ascórbico (Lukas-zewski y Blevins 1996). La pérdida de la dominancia apical causa la diferenciación de raíces laterales que no llegan a desarrollarse completamente, formándose aglomeraciones de raíces cortas y gruesas cerca de las puntas (Figura 3A). La temprana inhibición del crecimiento de las raíces en comparación con la aparición de los efectos en el tallo y las hojas causa aumentos de la relación tallo:raíz, lo cual puede predisponer a las plantas a experimentar volcamiento, deficiencias de agua y de nutrientes del suelo (Bennett 1996). Las raíces finas de las palmas aceiteras deficientes en boro son menos abundantes, adquieren apariencia enmarañada, lucen secas y más oscuras que las raíces de las palmas sanas (Figura 3).
Los síntomas descritos son la consecuencia de las funciones estructurales y metabólicas desempeñadas por el boro en las plantas terrestres en general. En la palma aceitera, su deficiencia causa aumentos de la concentración de ácidos fenólicos, los cuales inhiben la oxidación del AIA, con aumentos sustanciales en los niveles de auxinas (Rajaratnam 1974). Esto podría relacionarse con los síntomas semejantes a la etiolación observados en palmas adultas deficientes en boro (Figura 2D). Adicionalmente, el tamaño de los poros de las paredes celulares se incrementa conforme la disponibilidad disminuye. Debido a que el crecimiento depende de la estructura, la organización y la lignificación de los polisacáridos pécticos de la pared celular (Lӓuchli 2002), la deficiencia de boro causa inhibición de las expansión de las hojas y resulta en el debilitamiento y el doblamiento del cogollo.
Corrección de la deficiencia de boro
El diagnóstico científico completo de la presencia de deficiencias de boro en los cultivos y la caracterización de los síntomas asociados a su deficiencia, debe hacerse de manera complementaria en instalaciones hidropónicas “asépticas”, que garanticen la exclusión del elemento, empleando descriptores adecuados que permitan asociarlos con los síntomas expresados en el campo cuando las raíces crecen en el suelo. El análisis de la composición mineral de los tejidos y la observación periódica de los sistemas radicales deben acompañar la verificación y el monitoreo de campo basado en el reconocimiento de la sintomatología.
Debido a la naturaleza poco móvil del boro en la palma aceitera, y en gran medida debido a su función estructural, es difícil corregir sus deficiencias, que usualmente implica la aparición simultánea de otros desórdenes nutricionales como la deficiencia de calcio, debido parcialmente a la alteración de las membranas y a la disfunción de los transportadores involucrados en la absorción de otros elementos. A esto se suma el hecho de que transcurren varios meses antes de que una hoja nueva muestre síntomas y esté lo suficientemente desarrollada para ser muestreada (Shorrocks 1997, Broschat 2005).
El boro es usualmente aplicado en la forma de borato de sodio al suelo, en tasas de 0,01 a 0,035 kg B/ planta, en palmas jóvenes y adultas, respectivamente. Las formulaciones comerciales más conocidas son el Borax (Na2B4O2 • 10H2O, 11% B) y el Borato (Na2B4O2 • 5H2O, 14% B), ambas altamente solubles en agua y de rápida acción. La corrección inmediata puede lograrse mediante la aplicación de varias formulaciones químicas al suelo, a las hojas, y a otras partes de la planta como el cogollo o las hojas (Shorrocks 1997). Puede practicarse un “drench” o colocarse fuentes de boro de lenta liberación, sujetas a la lixiviación por la lluvia, en la forma de envoltorios de papel en las axilas de algunas hojas jóvenes (Rajaratnam 1976). Aunque la efectividad de este método ha sido verificada en el campo, la concentración idónea para la palma aceitera no ha sido determinada y el método puede causar distribución irregular dentro de la planta y toxicidad severa en casos de solubilización excesiva debida, por ejemplo, a periodos de alta precipitación.
Estos métodos tienen la desventaja común de que sus efectos no son permanentes, por lo que corrigen los problemas en los tejidos y órganos existentes al momento de la aplicación, pero no previenen los problemas nutricionales de las hojas y los frutos producidos subsecuentemente. Se recomienda también la remoción de sumideros fuertes de boro como las inflorescencias y los racimos (ablación) para reducir la demanda del elemento y acelerar la recuperación de las palmas deficientes (Patnude y Nelson 2012).
La corrección mediante la inyección de sustancias (Roach 1939, Boyle et al. 1991), un método fisiológico ampliamente documentado y utilizado en diferentes especies y situaciones, puede hacer una contribución significativa a la corrección duradera de la deficiencia de boro, porque introduce el nutriente directamente en los tejidos vasculares y facilita su transporte hacia los sumideros con poca transpiración y poco poder de atracción de minerales, como las hojas en expansión y los racimos jóvenes sujetos a la sombra del dosel. La efectividad de la inyección para introducir sustancias orgánicas e inorgánicas y aliviar los síntomas de diferentes tipos de estrés (Boyle et al. 1991), también ha sido bien documentada. La inyección como práctica hortícola con fines fitosanitarios es una práctica común en lugares donde las palmas son componentes importantes del paisaje urbano, como Hawaii y Florida. En la palma aceitera, la deficiencia de otros minerales de baja movilidad, como el hierro, ha sido corregida efectivamente mediante la inyección de sustancias férricas (FeSO4 • 7H2O) a través de las raíces gruesas y superficiales (Wanasuria et al. 1999).
Agradecimientos
Agradecemos a ASD-Costa Rica y Palma Tica, Coto 47, Costa Rica, por permitir el uso de sus plantaciones para realizar el trabajo de campo que sustenta esta publicación, y financiar los análisis nutricionales reportados en esta nota técnica.
Citas
4 Ramírez, F; Rodríguez, L. 2012. Estimación de requisitos nutricionales en dos clones de palma aceitera (correo electrónico). Coto 47, Corredores, Puntarenas, Costa Rica.
Literatura citada
Alves, M; Chicau, P; Matias, H; Passarinho, J; Pinheiro, C; Pinto-Ricardo, C. 2011. Metabolic analysis revealed altered amino acid profiles in Lupinus albus organs as a result of boron deficiency. Physiologia Plantarum 142:224-232. [ Links ]
Bell, RW. 1997. Diagnosis and prediction of boron deficiency for plant production. Plant and Soil 193(10):149-168. [ Links ]
Blevins, DG; Lukaszewski, KM. 1998. Boron in plant structure and function. Ann. Rev. Plant Physiol. Plant Molec. Biol. 49:481-500. [ Links ]
Bennett, WF. 1996. Plant nutrient utilization and diagnostic plant symptoms. In Bennett, WF. ed. Nutrient deficiencies and toxicities in plants. EEUU, Minnesota, APS Press. p. 1-7. [ Links ]
Bohnsack, CW; Albert, LS. 1977. Early effect of boron deficiency on indoleacetic acid oxidase levels of squash root tips. Plant Physiol. 59:1047-1050. [ Links ]
Boyle, MG; Boyer, JS; Morgan, PW. 1991. Stem infusion of liquid culture medium prevents reproductive failure of maize at low water potential. Crop Science 31:1246-1251. [ Links ]
Brown, PH; Shelp, BJ. 1997. Boron mobility in plants. Plant and Soil 193(7):85-101. [ Links ]
Broschat, TK. 2005. Boron deficiency in palms. ENH1012. Univ. Florida Extension Service. 6 p. [ Links ]
Campbell, LC; Miller, MH; Loneragan, JF. 1975. Translocation of boron to plant fruits. Aust. J. Plant Physiol. 2:481-487. [ Links ]
Carvajal, JF. 1972. Cafeto-cultivo y fertilización. Suiza, Basel, International Potash Institute. 139 p. [ Links ]
Chapman, GW; Gray, HM. 1949. Leaf analysis and the nutrition of the oil palm. Ann. Bot. 13(52):415-443. [ Links ]
Dannel, F; Pfeffer, H; Römheld, V. 2002. Update on boron in higher plants uptake, primary translocation and compartmentation. Plant Biol. 4:193-204. [ Links ]
Daroub SH; Snyder, GH. 2007. The chemistry of plant nutrients in soil. In Datnoff, LE; Elmer, WH; Huber, DM. eds. Mineral nutrition and plant disease. EEUU, St. Paul, APS Press. p. 1-7. [ Links ]
Dell, B; Huang, L. 1997. Physiological response of plants to low boron. Plant and Soil 193(8):103-120. [ Links ]
Dutta, TR; McIlrath, WJ. 1964. Effects of boron on growth and lignifications in sunflower tissue and organ cultures. Botan. Gaz. 125(2):86-96. [ Links ]
Fairhurst, TH; Härdter, R. 2003. Oil palm: management for large and sustainable yields. Basel, Suiza, International Potash Institute. 382 p. [ Links ]
Fairhurst, TH; Mutert, E. 1999. Interpretation and management of oil palm leaf analysis data. Better Crops International 13(1):48-51. [ Links ]
Frieden, E. 1984. Biochemistry of the essential ultra-trace elements. California, US, Plenum Publishing Corp. 421 p. [ Links ]
Goldberg, S. 1997. Reactions of boron with soils. Plant and Soil 193(3):35-48. [ Links ]
Hirsch, AM; Torrey, JG. 1980. Ultrastructural changes in sunflower root cells in relation to boron deficiency and added auxin. Can. J. Bot. 50:856-866. [ Links ]
Hu, H; Brown, PH. 1997. Absorption of boron by plants roots. Plant and Soil 193(4):49-58. [ Links ]
Kirkby, E. 2012. Introduction, definition and classification of nutrients. In Marschner, P. ed. Mineral nutrition of higher plants. San Diego, EEUU, Academic Press. p. 3-5. [ Links ]
Läuchli, A. 2002. Functions of boron in higher plants: recent advances and open questions. Plant Biol. 4:190-192. [ Links ]
Lewis, DH. 1980a. Are there interrelations between the metabolic role of boron, synthesis of phenolic phytoalexins and the germination of pollen? New Phytol. 84:261-270. [ Links ]
Lewis, DH. 1980b. Boron, lignifications and the origin of vascular plants, a unified hypothesis. New Phytologist 84:209-229. [ Links ]
Lukaszewski, KM; Blevins, DG. 1996. Root growth inhibition in boron-deficient or aluminum-stressed squash may be a result of impaired ascorbate metabolism. Plant Physiol. 112:1135-1140. [ Links ]
Matoh, T. 1997. Boron in plant cell walls. Plant and Soil 193(5):59-70. [ Links ]
Nable, RO; Bañuelos, GS; Paull, JG. 1997. Boron toxicity. Plant and Soil 193(12):181-198. [ Links ]
Nielsen, FH. 1997. Boron in human and animal nutrition. Plant and Soil 193(13):199-208. [ Links ]
Patnude, E; Nelson, S. 2012. Boron deficiency of palms in Hawaii. Plant disease PD-83. Col. Trop. Agric. Hum. Res. 6 p. [ Links ]
Rains, DW. 1976. Mineral metabolism. In Bonner, J; Varner, JE. eds. Plant biochemistry. 3 ed. EEUU, New York, Academic Press, p. 561-597. [ Links ]
Rajaratnam, JA. 1974. The distribution and mobility of boron within the oil palm (Elaeis guineensis). Ann. Bot. 36:289-297. [ Links ]
Rajaratnam, JA; Lowry, JB. 1974. The role of boron in the oil-palm (Elaeis guineensis). Ann. Bot. 38:193-200. [ Links ]
Raven, JA. 1980. Short and long distance transport of boric acid in plants. New Phytol. 84:231-249. [ Links ]
Roach, WA. 1939. Plant injection as a physiological technique. Annals of Botany 3(9):155-225. [ Links ]
Robinson JBD. 1993. Nutrient deficiencies and toxicities in crop plants. St. Paul, MN, EEUU, APS Press. 202 p. [ Links ]
Sah, RN; Brown, PH. 1997. Techniques for boron determination and their application to the analysis of plant and soil samples. Plant and Soil 193(2):15-33. [ Links ]
Shapira, O; Israeli, Y; Shani, U; Schwartz, A. 2012. Salt stress aggravates boron toxicity symptoms in banana leaves by impairing guttation. Plant Cell and Environment (en línea). Consultado 13 nov. 2012. Disponible en http://onlinelibrary.wiley.com/store/10.1111/j.1365-3040.2012.02572.x/asset/ pce2572.pdf?v=1&t=hdnfpjt3&s=73db0acf5451cd0b8fdd5b076f0a46757eb48776 [ Links ]
Shelp, BJ; Penner, R; Zhu, Z. 1992. Broccoli (Brassica oleracea var. italica) cultivar response to boron deficiency. Can. J. Plant. Sci. 72:883-888. [ Links ]
Shorrocks, VM. 1997. The occurrence and correction of boron deficiency. Plant and Soil 193(9):121-148. [ Links ]
Smith, PF. 1962. Mineral analysis of plant tissues. Ann. Rev. Plant Physiol. 13:81-108. [ Links ]
Sommer, AL; Lipman, CB. 1926. Evidence of the indispensable nature of zinc and boron for higher green plants. Plant Physiology 1:231-249. [ Links ]
Stangoulis, JCR; Graham, RD. 2007. Boron and plant disease. In Datnoff, LE; Elmer, WH; Huber, DM. eds. Mineral nutrition and plant disease. St. Paul, EEUU, APS Press. p. 207-214. [ Links ]
Takano, J; Miwa, K; Fujiwara, T. 2008. Boron transport mechanisms: collaboration of channels and transporters. Trends in Plant Science 13(8):451-457. [ Links ]
Tinker, PB. 1981. Levels, distribution and chemical forms of trace elements in food plants. Phil. Trans. R. Soc. Lond. B. 294:41-55 [ Links ]
Warington, K. 1923. The effect of boric acid and borax on the broad bean and certain other plants. Ann. Bot. 37(4):629-672. [ Links ]
Wanasuria, S; Setyobudi, H; Mayun, IB; Suprihatno, B. 1999. Iron deficiency of oil palm in Sumatra. Better Crops International 13(1):33-35. [ Links ]
Wang, D-N; Ko, WH. 1974. Relationship between deformed-fruit disease of papaya and boron deficiency. Phytopathology 65:445-447. [ Links ]
Welch, RM. 1995. Micronutrient nutrition of plants. Crit. Rev. Plant Sci. 14:49-82. [ Links ]
Wӧjcik, P; Cieslinski, G; Mika, A. 1999. Apple yield and fruit quality as influenced by boron application. Journal of Plant Nutrition 22(9):1365-1377. [ Links ]
Correspondencia a:
2Marco V. Gutiérrez-Soto: Universidad de Costa Rica, Estación Experimental Fabio Baudrit M., Apdo. 183-4050 Alajuela, Costa Rica. surdo26@racsa.co.cr
3Joaquín Torres-Acuña: ASD de Costa Rica, Departamento de Agronomía de Variedades y Clones. Coto 49, Corredores, Puntarenas, Costa Rica. jtorresa@numar.net.
Parte de proyecto de investigación con Agricultural Services & Development (ASD) - Costa Rica.
Recibido: 13 de noviembre, 2012. Aceptado: 28 de octubre, 2013.


{kind=link}
{kind=link}
{kind=link}