










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgronomía Mesoamericana
versão On-line ISSN 2215-3608versão impressa ISSN 1659-1321
Agron. Mesoam vol.24 no.2 San Pedro Jul./Dez. 2013
Identificación morfológica, morfométrica y molecular de meloidogyne incognita en higuera (ficus carica l.) en Costa Rica
Morphological, morphometrical and molecular identification of Meloidogyne incognita in fig (Ficus carica L.) in Costa Rica
Morphological, morphometrical and molecular identification of Meloidogyne incognita in fig (Ficus carica L.) in Costa Rica
Walter Peraza-Padilla2*, Johaner Rosales-Flores3*, Alejandro Esquivel-Hernández2, Irena Hilje-Rodríguez3, Ramón Molina-Bravo3, Pablo Castillo-Castillo4*
*Dirección de correspondencia
Resumen
El objetivo de este estudio fue identificar la especie de Meloidogyne asociada a dos plantaciones de higuera en Costa Rica. En marzo de 2012, en el distrito de Pacayas, provincia de Cartago, se detectaron agallas en los sistemas radicales de plantas de higuera (Ficus carica L.) de dos fincas. De las raíces agalladas se extrajeron hembras, masas de huevos y juveniles (J2) de Meloidogyne sp. Se examinaron los patrones perineales de las hembras y los segundos estadios infectivos se analizaron morfológica y molecularmente mediante PCR-RFLP. Se amplificó la región intergénica (IGS) del genoma mitocondrial, delimitada por el gen de la subunidad II de la citocromo oxidasa (COII) y el gen ribosomal 16S. La población de nematodos se identificó como M. incognita. El tamaño de los productos de PCR generados con los imprimadores C2F3 y 1108 fue de 1,7 kb.
Al tratar los productos de PCR con enzimas de restricción, se generaron cuatro fragmentos de 850, 450, 250 y 150 pb con la enzima AluI y dos fragmentos de 1300 y 400 pb con la enzima HinfI.
Palabras clave: nematodo agallador, patrones perineales, taxonomía de nematodo.
Abstract
The objective of this study was to identify species of Meloidogyne associated with two fig plantations in Costa Rica. On March 2012 in Pacayas, Cartago province, root-galls were found in two fig plantations of Ficus carica L. Females, egg-masses and juveniles (J2) of Meloidogyne sp. were extracted from the galled roots. Female perineal patterns were examined and second infective stages were analyzed morphometrically and molecularly by PCR-RFLP. The mitochondrial intergenic region (IGS) flanked by the cytochrome oxidase subunit II gene (COII) and the 16S ribosomal gene was amplified. The population was identified morphologically, morphometrically and molecularly as M. incognita. The PCR product obtained with primers C2F3 and 1108 were 1.7 kb in size. When PCR products were treated with restriction enzymes they generated four fragments of 850, 450, 250 and 150 bp with AluI and two fragments of 1300 and 400 bp with HinfI.
Keywords: root-knot nematode, perineal patterns, taxonomy nematode.
Introducción
La higuera (Ficus carica L.) y su fruto comestible, el higo, es una de las frutas más antiguas conocidas por el hombre. Es originario de la región mediterránea y se consume como fruto seco en un 90% y enlatado o como conserva en un 10% (Bolin y King 1980).
Actualmente, se cultiva en más de 20 países siendo Egipto, Turquía y Argelia los principales productores en toneladas a nivel mundial con 304 000, 205 000 y 79 000 respectivamente. En América sobresale Brasil con una producción de 25 727 toneladas al año (FAOSTAT 2010). Según datos del 2009, Costa Rica produjo más de 7000 kg de higo, de los cuales el 94% se comercializó como fruto fresco. Se estima que en la zona de Cartago (Tierra Blanca, Llano Grande) existen alrededor de 47 productores (Miranda 2009); sin embargo, esta cifra podría ser mayor ya que en la zona de Prusia, Tobosi de Cartago y Pérez Zeledón existen productores que iniciaron recientemente la producción de higo (Barboza 2007).
Los nematodos del género Meloidogyne (Nemata:Heteroderidae) constituyen una de las principales plagas agrícolas en una gran variedad de cultivos anuales y perennes (Goeldi 1892). Se estima que estos patógenos pueden causar pérdidas económicas de hasta un 20% en una amplia gama de plantas hospederas (Padgham et al. 2004). Sherb (1993), menciona que dentro de los problemas fitosanitarios de la higuera se encuentra Meloidogyne, considerado una plaga limitante del rendimiento y la producción junto con la roya (Cerotelium fici). Se han documentado numerosos casos de daños por nematodos agalladores en la cuenca Mediterránea, en el norte y sur de América, y en el sur de África. En Costa Rica, López (1980) reportó por primera vez este nematodo asociado a este cultivo en fincas localizadas en Llano Grande de Cartago. En un estudio posterior, se confirmó la presencia en plantaciones de higuera en San Rafael de Escazú, San José (López et al. 1991).
Entre las especies asociadas a este cultivo en el mundo se encuentran M. arenaria, M. javanica y M. incognita (McSorley 1981), esta última en mayor frecuencia. La infección de plantaciones comerciales de higuera por M. incognita fue previamente registrada en Colombia (Volcy 1986, Tamayo y Becerra 2002), Japón (Hosomi y Uchiyama 1998) y Brasil (Rossi 2002, Medina et al. 2006, Dias-Ariera et al. 2008, Bauer et al. 2009, Formentini, 2009). La presencia de este nematodo en el cultivo afectó en el pasado la producción comercial en Estados Unidos (Knight Jr 1980), Francia (Scotto La Massèse et al. 1984) y Brasil (Ferraz et al. 1982, Campos 1997), países con las mayores producciones por hectárea del mundo. En Costa Rica, este nematodo está asociado a una gran variedad de cultivos de importancia agrícola y ornamental, pero no existe en la mayoría de los casos, datos que relacionen daños a las plantas con la presencia de Meloidogyne sp. El cultivo de la higuera en Costa Rica ha crecido en área cultivada y recientes prospecciones detectaron plantaciones con síntomas de este nematodo fitopatógeno.
En la actualidad, el género Meloidogyne incluye más de 90 especies (Siddiqi 2000, de Waele y Elsen 2007). De acuerdo con Vovlas et al. (2005), la identificación correcta de este nematodo facilita la implementación de medidas de combate, como el uso de cultivares resistentes y de medidas cuarentenarias.
Comúnmente, la identificación a nivel de especie se realiza mediante la observación de caracteres morfológicos y morfométricos (Taylor y Netscher 1974, Esser et al. 1976, Taylor y Sasser 1978); sin embargo, estos procedimientos requieren mucho tiempo y un amplio conocimiento sobre taxonomía de nematodos.
Además, la plasticidad fenotípica, variabilidad intraespecífica y similitudes morfológicas entre especies dificulta su identificación (Poinar 1990, Pamjav et al. 1999). Usualmente la identificación morfológica se realiza mediante el estudio del patrón perineal, región que se encuentra en la parte posterior de la hembra y consiste en líneas laterales o pliegues cuticulares que son propios para cada especie (Taylor y Netscher 1974). No obstante, las frecuentes variaciones intraespecíficas de los patrones perineales los hace poco confiables para la identificación a nivel de especie (Xu et al. 2004).
Otros métodos de identificación complementan los procedimientos tradicionales y permiten una caracterización rápida y precisa. El empleo de la reacción en cadena de la polimerasa (PCR por sus siglas en inglés) (Mullis et al. 1986) y la técnica RFLP (Polimorfismo en la Longitud de los Fragmentos de Restricción) facilitan el análisis de regiones del genoma de los organismos (Louws et al. 1999) y permiten la identificación molecular de los nematodos mediante el análisis de su ADN (Hussey 1990, Powers 2004). El método de PCR-RFLP primero genera un producto de PCR específico a varias especies y posteriormente, las con un tamaño de producto similarson diferenciadas por los patrones o fragmentos que se generan con enzimas de restricción. La combinación de estos dos métodos le facilita a los científicos distinguir entre las diferentes especies de Meloidogyne de manera rápida y a bajo costo. Una de las regiones que se utiliza con mayor frecuencia para diferenciar especies es la región intergénica (IGS) entre los genes mitocondriales, específicamente entre la subunidad II de la citocromo oxidasa (COII) y la subunidad grande (16S) del ADN ribosomal (Powers y Harris 1993). Esta región mitocondrial contiene secuencias altamente variables entre especies y está delimitada por genes muy conservados, lo cual crea una situación ideal para análisis y diagnóstico molecular. En el presente estudio se integraron métodos morfológicos, morfométricos y moleculares para determinar la especie de Meloidogyne responsable de producir agallas en el sistema radical de plantas de higuera provenientes de dos fincas productoras de la localidad de Pacayas, Cartago, Costa Rica.
Materiales y Métodos
Recolecta de material vegetal. Se recolectaron raíces de higuera (F. carica L.) durante los meses de febrero y marzo del 2012, en dos fincas productoras de la localidad de Pacayas, Cartago. El primer sistema de producción se localizó a los 09º 55’41,50’’N y 083º48’33,01’’O y el segundo a los 09º 55’47,49’’N y 083º48’44,09’’O. Ambas plantaciones se establecieron hace aproximadamente tres años. En cada una de las fincas, se tomaron cinco muestras de raíces noduladas o agalladas. Dichas raíces se llevaron al Laboratorio de Nematología y los nematodos al Laboratorio de Biología Molecular de la Escuela de Ciencias Agrarias de la Universidad Nacional de Costa Rica para su análisis.
Extracción e identificación morfológica y morfométrica. Se extrajeron hembras y masas de huevos de Meloidogyne sp. a partir de raíces con agallas con ayuda de una aguja de disección y bajo un estereoscopio a 45X. Se prepararon patrones perineales de 10 a 15 hembras de acuerdo con el método descrito por Hartman y Sasser (1985). La caracterización morfométrica de juveniles de segundo estadio (J2) se llevó a cabo mediante la observación de doce especímenes.
Las observaciones y mediciones se realizaron con ayuda de un microscopio compuesto a 100X. Se comparó la morfología de los diseños con descripciones e ilustraciones de varios autores (Esser et al. 1976, López y Dickson 1977, Taylor y Sasser 1978, Whitehead 1968). Se tomaron fotografías bajo el microscopio con una cámara digital integrada a 100X.
Identificación molecular
Extracción de ADN. Se extrajo el ADN de juveniles infectivos (J2) mediante la técnica de maceración directa del nematodo (Powers y Harris 1993). Se colocó un nematodo en 15 μl de agua ultrapura sobre un cubre objetos y se maceró a presión con una punta de micropipeta estéril bajo un microscopio. Posteriormente, dicha solución con el ADN se transfirió a un tubo de microcentrifuga y se conservó a -20°C para su posterior procesamiento y análisis.
Amplificación de ADN (PCR). Para el análisis molecular se utilizaron los imprimadores diseñados por Powers y Harris (1993), C2F3 (5’-GGTCAATGTTCAGAAATTTGTGG-3’) y 1108 (5’-TACCTTTGACCAATCACGCT- 3’), los cuales amplifican una región del ADN mitocondrial entre los genes COII y 16S. La reacción se colocó en un tubo de 0,2 ml para PCR a un volumen total de 25 μl que incluyó buffer de la Taq polimerasa 1X, dNTPs 0,08 mM, MgCl2 1,0 mM, imprimadores a 0,8 μM cada uno, 0,8 U de Taq polimerasa y 15 μl del ADN. El perfil de temperaturas consistió en un primer paso de desnaturalización a 94°C durante 2 minutos, seguido de 35 ciclos de 10 segundos de desnaturalización a 94°C, 30 segundos de alineamiento a 48°C, 2 minutos de extensión a 68°C y una fase de extensión final a 72°C durante 2 minutos. La reacción se llevó a cabo en un termociclador de 96 pozos.
Tratamiento enzimático de los fragmentos amplificados (RFLP). Los productos de PCR se trataron posteriormente con las enzimas de restricción AluI y HinfI en forma independiente, según las indicaciones del fabricante. Las reacciones se realizaron en un volumen total de 15 μl (5 μl de producto de PCR, 1 μl de enzima, 1,5 μl del buffer de reacción de la enzima a 1X, 7,5 μl de agua ultrapura) y se incubaron durante una hora a 37°C en un termociclador de 96 pozos.
Electroforesis. Los fragmentos de PCR y RFLP se separaron por medio de electroforesis en geles de agarosa de 1,5 y 2% respectivamente y preparados con TBE 1X. Los geles se corrieron dentro de una cámara de electroforesis a 90V durante 1 hora y 30 minutos.
Los fragmentos de ADN se visualizaron y se digitalizaron en un foto-documentador con un transiluminador de luz UV después de teñir los geles con GelRed® (Biotium) a 3X. El tamaño de las bandas se calculó con un programa de análisis de datos electroforéticos.
Resultados y Discusión
Las raíces recolectadas en ambas parcelas mostraron síntomas severos de nodulación radical, tanto en zonas apicales como en zonas intermedias (Figura 1).
En el interior de dichas agallas se encontraron varias hembras de Meloidogyne que se extrajeron mediante disección directa para su identificación.
Identificación morfológica
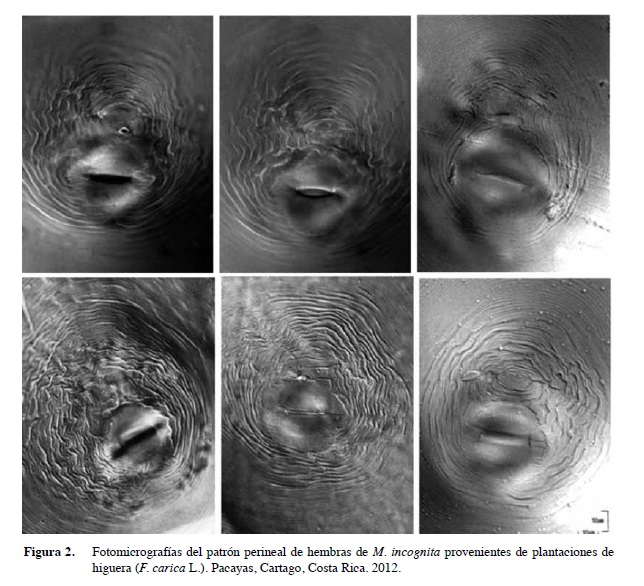
Patrones perineales. Se estudiaron y analizaron al menos diez cortes perineales de hembras ovígeras.
Los patrones perineales se caracterizaron por presentar en su mayoría, un arco dorsal alto y ligeramente achatado; sin embargo, se observaron algunas variaciones intraespecíficas en las poblaciones estudiadas. La posición del poro excretor de las hembras coincidió con el de M. incognita (Figura 2).
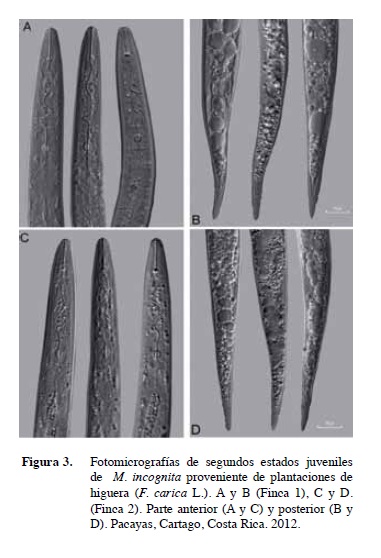
La morfología de los juveniles aislados en el cultivo de higuera fue muy similar entre sí pero con algunas diferencias con respecto a la descripción original.
Algunos de los caracteres morfológicos de importancia diagnóstica (Esser et al. 1976, Jepson 1987, Taylor 1978), tales como longitud del cuerpo, longitud del estilete, longitud de la cola y porción hialina de la cola mostraron en ambas poblaciones estudiadas, rangos de valores superiores que los observados en la descripción realizada por Kofoid y White (1919), Chitwood (1949), Orton-Williams (1973) y Jepson (1987) de M. incognita (Figura 3).
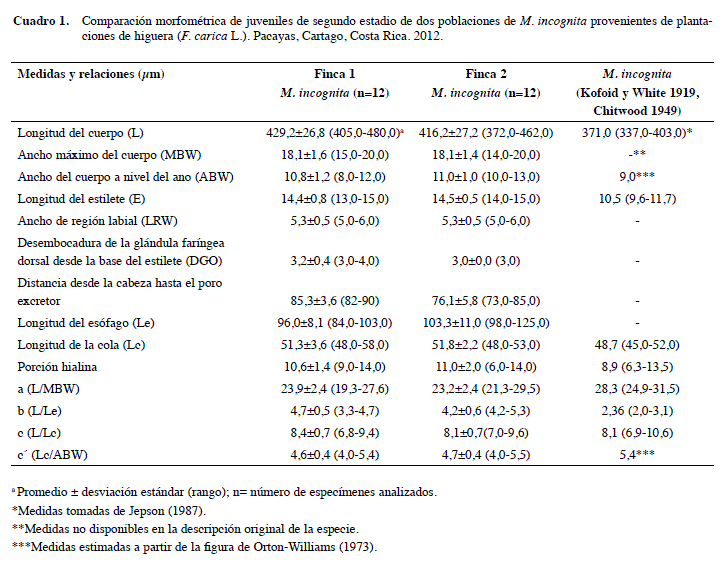
El análisis morfométrico de doce juveniles o J2 de M. incognita junto con la descripción original de M. incognita realizada por Orton-Williams (1973) se describe en el Cuadro 1. Los valores promedio de la longitud del cuerpo de las dos poblaciones (429,2 μm para la Finca 1 y 416,2 μm para la Finca 2) fueron mayores que el promedio reportado (Cuadro 1) el cual fue de 371,0 μm. Por otra parte, los valores medios de la longitud de la cola (48,7 μm) y la porción hialina de la cola (8,9 μm) descritos por Jepson (1987) para M. incognita son menores a los encontrados en este estudio (51,3 y 10,6 μm para la Finca 1) y (51,8 μm y 2,2 μm para la Finca 2). Existe la posibilidad de encontrar variabilidad en los parámetros morfométricos dentro de una misma especie, es decir, superposición en algunas medidas. Los promedios de las medidas obtenidas en este estudio están dentro del rango establecido para la especie (Kofoid y White 1919, Chitwood 1949). Estas diferencias en los caracteres morfológicos de J2 entre poblaciones de M. incognita de Costa Rica, pueden estar ligadas a factores ambientales que influyen directamente con el desarrollo de los estados juveniles.
Identificación molecular
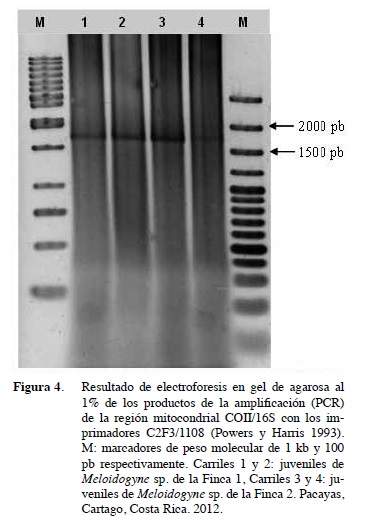
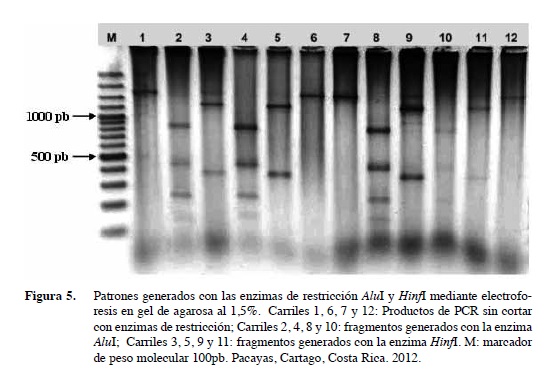
La amplificación (PCR) de la región mitocondrial COII/16S con los imprimadores C2F3/1108 generó un fragmento de 1,7 kb (Figura 4) en todas las muestras analizadas lo que concuerda con otros estudios de M. incognita (Powers y Harris 1993, Williamson et al. 1994, Han et al. 2004). Los fragmentos obtenidos con las enzimas de restricción fueron de 850, 450, 250 y 150 pb con la enzima AluI y de 1300 kb y 400 pb con la enzima HinfI (Figura 5). Estos fragmentos coinciden con los obtenidos por Han et al. (2004) y Orui (1998) en sus investigaciones al cortar con ambas enzimas de restricción. Sin embargo, Powers y Harris (1993), reportaron que el producto de PCR de M. incognita tratado con la enzima de restricción HinfI generó tresfragmentos de 1000, 400 y 300 pb, mientras que Williamson et al. (1994) en Estados Unidos, Oriu (1998) en Japón, Han et al. (2004) en Corea y Maleita et al. 2012 en Portugal al igual que en este estudio en Costa Rica, mostraron sólo dos bandas de 1300 y 400 pb con dicha enzima de restricción. Aunque los nematodos presentaron una morfología muy similar, esas variaciones en el tamaño de los fragmentos que se generan con las enzimas de restricción supone variantes genéticas o polimorfismos dentro de una misma especie o población que puede deberse a mutaciones en el sitio de corte de las enzimas de restricción.
Tradicionalmente la identificación de especies de Meloidogyne se fundamenta en la morfología de hembras adultas o en la obtención y análisis de ADN a partir de un pool de hembras (Flores 2008, Humphreys et al. 2012). Este es el primer estudio que logra la amplificación y corte de ADN a partir de un solo juvenil de Meloidogyne. Además es el primer estudio en donde se identifica molecularmente a la especie M. incognita infectando plantas de higuera en Costa Rica. El diagnóstico molecular de las especies a partir de un solo juvenil ofrece ventajas comparativas con los métodos antes descritos, por la facilidad de aislar de muestras de suelo o raíz los estados infectivos (J2), lo que efectivamente simplifica y agiliza el proceso de reconocimiento. La identificación correcta de las especies de Meloidogyne asociadas a cultivos en nuestro país es esencial para conocer su distribución y propagación e implementar estrategias de manejo. Así mismo, existe hoy día la apertura de mercados internacionales para el comercio de cultivos y ornamentales, que puede favorecer la introducción y diseminación de especies de Meloidogyne no autóctonas. Se desconoce la extensión del problema de Meloidogyne en el cultivo de higuera en Costa Rica, por lo que es necesario realizar prospecciones fitonematológicas en otras áreas productoras para determinar la magnitud del daño y si la especie de Meloidogyne asociada es la misma en todas las zonas. Finalmente, es necesario estar alerta a la infección de este nematodo para evitar su diseminación en otros sitios donde se produce este fruto.
Agradecimientos
Los autores desean expresar su agradecimiento a la Fundación CRUSA-CSIC por el financiamiento y apoyo al proyecto “Diagnóstico y caracterización bioquímica y molecular de nematodos noduladores (Meloidogyne spp.) en cultivos tropicales y hortícolas de Costa Rica”. Al proyecto “Aplicación de tecnologías innovadoras para contribuir con el desarrollo sostenible del cultivo del higo y mora en Costa Rica” financiado por la Comisión Nacional de Rectores (CONARE).
Literatura citada
Bauer, C; Somavilla, L; Carneiro, RMDG; Desiré, A; da Costa, Fábia; Lima, I. 2009. Monitoramento do nematóide das galhas (Meloidogyne spp.) em figueira (Ficus carica L.) no Rio Grande do Sul, Brasil. Embrapa Clima Temperado. 18 p. (Boletim de pesquisa e desenvolvimento 86). [ Links ]
Barboza, S. 2007. Informe final: Desarrollo del cultivo de higo (Ficus carica) para consumo fresco y procesado, como una alternativa de diversificación para el sector agrícola. Cartago, Costa Rica, Instituto Tecnológico de Costa Rica. 30 p. [ Links ]
Bolin, HR; King, AD. 1980. Figs. In Nagy, S; Shaw, PE. (eds) Tropical and Sub-tropical fruits. Composition, properties and uses. Westport, Connecticut, AVI Publishing, Inc. p. 492-505. [ Links ]
Campos, VP. 1997. Nematodes of the fig crop. Informe Agropecuario Belo Horizonte 18188. Brasil. p. 33-38. [ Links ]
Chitwood, BG. 1949. Root-know nematodes, part I. A revision of the genus Meloidogyne Goeldi, 1887. Proceedings of the Helminthological Society of Washington 16:90-104. [ Links ]
De Waele, D; Elsen, A. 2007. Challenges in tropical plant nematology. Annual Review of Phytopathology 45:457-485. [ Links ]
Dias-Ariera, CR; Molina, RO; Costa, AT. 2008. Nematóides causadores de doenças em frutíferas. Boa Vista, Brasil (en línea). Consultado 15 may. 2012. Disponible en http://revista.ufrr.br/index.php/agroambiente/article/download/230/169 [ Links ]
Esser, RP; Perry, VG; Taylor, AL. 1976. A diagnostic compendium of the genus Meloidogyne Nematoda: Heteroderidae. Proceedings of the Helminthological Society of Washington 43:138-150. [ Links ]
FAOSTAT (Food and Agriculture Organization Corporate Statistical Database). 2010. Crops production (en línea). Consultado 21 abr. 2012. Disponible en http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor [ Links ]
Ferraz, LCCB; Pereira, FM; Valente, J. 1982. Considerações sobre o use de nematicidas em viveiro visando a recuperacao demudas de Ficus carica ev. roxo de valinhos infestadas por Meloidogyne incognita. In Lordello, LGE. ed. Trabalhos apresentados a VI reunias Brasileira de nematologia. Publicacao No. 6. Sociedade Brasileira de Nematologia, Piracicaba, São Paulo, Brasil. p. 13-20. [ Links ]
Flores, L. 2008. Identificación de especies de Meloidogyne descritas en Costa Rica con base en la variabilidad del ADN mitocondrial. Tesis de Maestría. Costa Rica, Universidad de Costa Rica. 48 p. [ Links ]
Formentini, E. 2009. Manipueira no control de Meloidogyne incognita e no rendimento da figueira (Ficus carica L.) cv. Roxo de Valinhos no oeste Paranaense. Dissertação de Mestrado. BR, Univerdidade Estadual do Oeste do Paraná. 60 p. [ Links ]
Goeldi, EA. 1892. Relatorio sobre a molestia do cafeeiro na provincia do Rio de Janeiro. Arquivos do Museu Nacional. Rio de Janeiro, BR. 8:7-121. [ Links ]
Han, H; Cho, MR; Jeon, ChY; Lim, ChK; Jang, HL. 2004. PCR-RFLP identification of three major Meloidogyne species in Korea. Journal of Asia-Pacific Entomology 7(2):171-175. [ Links ]
Hartman, KM; Sasser, JN. 1985. Identification of Meloidogyne species on the basis of differential host test and perineal patterns morphology. In Barker, KR; Carter, CC; Sasser, JN. eds. An advanced treatise on Meloiodyne. v2. Methodology. Raligh, USA, North Carolina State University Graphics. p. 69-77. [ Links ]
Hosomi, A; Uchiyama, T. 1998. Growth inhibiting factors in sick soil of fig orchards. Journal of the Japanese Society for Horticultural Science 67:44-50. [ Links ]
Humphreys, DA; Williamson, V; Salazar, L; Flores-Cháves, L; Gómez-Salazar, L. 2012. Presence of Meloidogyne enterolobii Yang and Eisenback (=M. mayaguensis) in guava and acerola from Costa Rica. Nematology 14:199-207. [ Links ]
Hussey, RS. 1990. Biochemical and molecular methods of identifying Meloidogyne species: symposium introduction. Journal of Nematology 22:8-9. [ Links ]
Knight, RJ. 1980. Origin and world importance of tropical and sub-tropical fruit crops. In Nagy, S; Shaw, PE. eds. Tropical and sub-tropical fruits. Composition, poperties and uses. Westport, Connecticut, USA, AVI Publishing, Inc. p. 1-120. [ Links ]
Kofoid, CA; White, WA. 1919. A new nematode infection of man. Journal of the American medical Association 72:567-569. [ Links ]
Jepson, SB. 1987. Identification of root-knot nematode (Meloidogyne species). Wallingford, UK, CAB International. 265 p. [ Links ]
López R; Dickson, DW. 1977. Morfometría y respuesta de hospedantes diferenciales a tres poblaciones de Meloidogyne incognita y una de Meloidogyne javanica. Agronomía Costarricense 1:119-127. [ Links ]
López, R. 1980. Cambios estacionales de la distribución espacial de namatodos de un plantío de higo. Nematropica 10(1):2. [ Links ]
López, R; Salazar, L; Azofeifa, J. 1991. Observaciones sobre la morfología de Meloidogyne incognita con el microscopio electrónico de rastreo. Agronomía Costarricense 15:105-112. [ Links ]
Louws, FJ; Rademaker, KLW; de Bruijin, FJ. 1999. The three Ds of PCR-Based Genomic Analysis of Phytobacteria: Diversity, Detection, Disease Diagnose. Annual Review Phytopathology 37:81-125. [ Links ]
Maleita, CM; Simões, MJ; Egas, C; Curtis, RHC; Abrantes, IM. de O. 2012. Biometrical, biochemical, and molecular diagnosis of Portuguese Meloidogyne hispanica isolates. Plant Disease 96:865-874. [ Links ]
McSorley, R. 1981. Plant parasitic nematodes associated with tropical and sub-tropical fruits. Bulletin 823 of Agricultural Experiment Station Institute of Food and Agricultural Science. Gainesville, Florida, USA, University of Florida. 51 p. [ Links ]
Medina, IL; Bauer, GC; Rossi, C; Carneiro, RM DG. 2006. Caracterização e identifação de polulações de namatóides de galhas provenientes de figueiras (Ficuas carica L.) do Rio Grande do Sul e de São Paulo. Nematologia Brasileira 30(2):179-187. [ Links ]
Miranda, D. 2009. Evaluación de alternativas frutícolas amigables con el ambiente para contribuir al desarrollo de la zona norte de Cartago. Centro Internacional de Política Económica (CINPE) de la Universidad Nacional. Costa Rica. 18 p. [ Links ]
Mullis, K; Faloona, F; Scharf, S; Saiki, R; Horn, G; Erlich, H. 1986. Specific enzymatic amplification of DNA in vitro: the polymerase chain reaction. Cold Spring Harbor Symposium in Quantitative Biology 51:263-73. [ Links ]
Orui, Y. 1998. Identification of Japanese species of the genus Meloidogyne (Nematoda: Meloidogynidae) by PCR-RFLP analysis. Applied Entomology and Zoology 33:43-51. [ Links ]
Orton-Williams, KJ. 1973. Meloidogyne incognita. C.I.H. descriptions of plant parasitic nematodes. Commonwealth Institute of Helminthology, St. Albans, UK. Set 2, No. 18. 4 p. [ Links ]
Padgham, JL; Mazid, MA; Duxbury, JM; Abawi, GS; Hossain, M. 2004. Yield loss caused by Meloidogyne graminicola on lowland rainfed rice in Bangladesh. Journal of Nematology 36:42-48. [ Links ]
Pamjav, H; Triga, D; Buzas, ZT; Vellai, A; Lucska, I; Adams, A; Reid, AP; Burnell, A; Griffin, C; Glazer, I; Klein, MG; Fodor, A. 1999. Noval application of PhastSystem polyacrylamide gel electrophoresis using restriction fragment length polymorphism - internal transcribed spacer patterns of individuals for molecular identification of entomopathogenic nematodes. Electrophoresis 20:1266-1273. [ Links ]
Poinar, GO. 1990. Taxonomy and biology of Steinernematidae and Heterorhabditidae. In Gaugler, R; Kaya, HK. eds. Entomopathogenic nematodes in biological control. Boca Raton, USA, CRC Press. p. 23-60. [ Links ]
Powers, TO. 2004. Nematode molecular diagnostics: from bands to bar-codes. Annual Review of Phytopathology 42:367-383. [ Links ]
Powers, TO; Harris, TS. 1993. A polymerase chain reaction method for identification of five major Meloidogyne species. Journal of Nematology 25:1-6. [ Links ]
Rossi, C. 2002. Nematóides em fruteiras. VII Reunião Itinerante de Fitossanidade do Instituto Biológico. Centro Experimental Central do Instituto Biológico. Laboratório de Nematologia. Brasil. 130 p. [ Links ]
Sherb, CT. 1993. Flutuação populacional de Meloidogyne incognita (Kofoid y White, 1919) Chitwood, 1949 em figueira (Ficus carica L.) inoculadas no campo. Tese Mestrado. ESALQ. Lavras, MG, Brasil. 150 p. [ Links ]
Scotto La Massèse, C; Deportes, L; Mercier, S; Roger, J. 1984. Les principaux ennemis du figuier: les nematodes et maladies. Phytoma – Défense des Cultures 354:39-41. [ Links ]
Sidiqqui, M. 2000. Tylenchida: parasites of plants and insects. 2 ed. London, UK, Biddles. 833 p. [ Links ]
Tamayo, PJ; Becerra. 2002. Otra enfermedad del brevo (Ficus carica L.) en el Departamento de Antioquía. Informa 28. Paipa, Boyacá, Colombia, ASCOLFI. p. 46-47. [ Links ]
Taylor, DP; Netscher, C. 1974. An improved technique for preparing perineal patterns of Meloidogyne spp. Nematologica 20:268-269. [ Links ]
Taylor, AL; Sasser, JM. 1978. Biology, identification and control of root-knot nematodes (Meloidogyne species). Raleigh, North Carolina, USA, North Carolina State University Graphics. 111 p. [ Links ]
Volcy, Ch. 1986. Registros de nematodos fitoparásitos para Colombia. In Resúmenes VII Congreso de la Asociación Colombiana de Fitopatología y Ciencias Afines. Paipa, Boyacá, Colombia, ASCOLFI. 39 p. [ Links ]
Vovlas, N; Mifsud, D; Landa, BB; Castillo, P. 2005. Pathogenicity of the root-knot nematode Meloidogyne javanica on potato. Plant Pathology 54:657-664. [ Links ]
Whitehead, AG. 1968. Taxonomy of Meloidogyne (Nematodea: Heteroderidae) with descriptions of four new species. Transactions of the Zoological Society of London 31:263-401. [ Links ]
Williamson, VM; Caswell-Chen, EP; Wu, FF; Hanson, D. 1994. PCR for nematode identification. In Lamberti, F; De Giorgi, C; Bird, DM. eds. Advances in molecular plant Nematology. New York, USA, Plenum Press. p. 119-127. [ Links ]
Xu, J; Liu, P; Meng, Q; Long, H. 2004. Characterisation of Meloidogyne species from China using isozyme phenotypes and amplified mitochondrial DNA restriction fragment length polymorphism. European Journal of Plant Pathology 110:309-315. [ Links ]
*Correspondencia a:
2Walter Peraza-Padilla. Laboratorio de Nematología, Escuela de Ciencias Agrarias. Universidad Nacional. Heredia, Costa Rica. 86-3000. wperaza@una.ac.cr
3Johaner Rosales-Flores. Laboratorio de Cultivo de Tejidos y Células Vegetales, y Laboratorio de Biología Molecular. Universidad Nacional. Heredia, Costa Rica. 86-3000. johngarden@gmail.com
2Alejandro Esquivel-Hernández. Laboratorio de Nematología, Escuela de Ciencias Agrarias. Universidad Nacional. Heredia, Costa Rica. 86-3000.aesquive@una.ac.cr
3Irena Hilje-Rodríguez. Laboratorio de Cultivo de Tejidos y Células Vegetales, y Laboratorio de Biología Molecular. Universidad Nacional. Heredia, Costa Rica. 86-3000. ihilje@una.ac.cr
3Ramón Molina-Bravo. Laboratorio de Cultivo de Tejidos y Células Vegetales, y Laboratorio de Biología Molecular. Universidad Nacional. Heredia, Costa Rica. 86-3000. rmolin@una.ac.cr
4Pablo Castillo-Castillo.Instituto de Agricultura Sostenible (IAS), Consejo Superior de Investigaciones Científicas (CSIC). Apdo. 4084, 14080 Córdoba, España. pcastillo@ias.csic.es
Esta investigación forma parte del proyecto “Diagnóstico y caracterización bioquímica y molecular de nematodos noduladores (Meloidogyne spp.) en cultivos tropicales y hortícolas de Costa Rica” financiado por la Fundación CRUSA-CSIC.
Recibido: 8 de junio, 2012. Aceptado: 28 de octubre, 2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}