










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Mesoamericana
On-line version ISSN 2215-3608Print version ISSN 1659-1321
Agron. Mesoam vol.23 n.1 San Pedro Jun. 2012
Fenología, producción y contenido de almidón en árboles de mango en Guanacaste, Costa Rica
*Dirección para correspondencia
Resumen
Fenología, producción y contenido de almidón en árboles de mango en Guanacaste, Costa Rica. El objetivo de este trabajo fue determinar la variación estacional del contenido de almidón en árboles de mango y relacionarla con sus patrones fenológicos y de producción. Se estudiaron árboles de las variedades Tommy Atkins (TA) y Keitt (K), en Guanacaste, Costa Rica. Los estados fenológicos evaluados fueron: (1) aparición de brotes vegetativos, (2) brotes florales y (3) producción de frutos. Esta última se registró como peso de frutos por árbol. El almidón se determinó mediante hi- drólisis enzimática de madera de raíces, troncos y ramas en cuatro estados fenológicos durante 2004 y 2005. Además se registró la temperatura y la precipitación. Los contenidos de almidón fluctuaron a través del período analizado. Se obser- vó que estuvieron influenciados por las etapas fenológicas de producción de flores y frutos. El contenido de almidón pre- sentó concentraciones más altas (TA: 11,7%, K: 14,4%) en la etapa de reposo y prefloración, entre agosto y noviembre. Los menores niveles (6,1%) se observaron en poscosecha en mayo para las dos variedades. La variedad Tommy Atkins presentó la mayor producción en el 2006 con 208,5 kg/árbol y la menor producción en 2005 con 34,9 kg, mientras que en la variedad Keitt, la mayor producción fue en el 2005 con 212,1 kg y la menor se registró en el 2004 con 36,7 kg/árbol. Los años de mayor producción en las dos variedades pueden corresponder a años “on” (media o alta producción), los de menor producción a años “off” (baja o mala producción) y el 2004 en la variedad Tommy Atkins presentó una cosecha intermedia de 86,7 kg/árbol.
Palabras claves: Mangifera indica L, crecimiento vegetativo, floración, variedad Tommy Atkins.
Abstract
Phenology, production and starch content of mango trees in Guanacaste, Costa Rica. The objective of this work was to determine the seasonal variation of the starch content of mango trees and its relationships to phenology and fruit production. Trees of Tommy Atkins (TA) and Keitt (K) varieties were studied in Guanacaste, Costa Rica. The phenological stages recorded were: (1) appearance of vegetative buds, (2) flowering, and (3) fruit production. The latter represented the total weight of fruits per tree. Starch content was determined through enzymatic hydrolysis of wood from roots, trunks and branches for four phenological stages during 2004 and 2005. Temperature and precipitation was also recorded. Starch content varied throughout the study, and was influenced by flower and fruit production, with the highest concentration (TA: 11,7%, K: 14.4%) just before flowering (between August and November). The lowest concentration of starch (6.1%) was recorded at postharvest for both varieties (May). The Tommy Atkins variety had the highest production in 2006 with 208,5 kg/tree and the lowest production in 2005 with 34.9 kg. In contrast, peak production occurred in the Keitt variety in 2005 with 212.1 kg, and the lowest in 2004 with 36.7 kg/tree. The highest harvest in both varieties could be assigned to “on” years and the lowest to “off” years, while 2004 in Tommy Atkins could be considered an intermediate harvest of 86.7 kg/tree.
Key words: Mangifera indica L, vegetative growth, flowering, Tomy Atkins variety.
Introducción
El árbol de mango presenta flujos o episodios de crecimiento más numerosos en condiciones tropicales comparado con el subtrópico. En las regiones tropicales el crecimiento se rige principalmente por las variaciones en la disponibilidad de agua en el suelo, mientras que en el subtrópico es regido básicamente por cambios en la temperatura. En general, en cada ciclo anual del cultivo se dan varios episodios de crecimiento vegetativo y uno de crecimiento reproductivo. La época de floración está influenciada por temperatura, precipitación, variedad utilizada, manejo que reciba la planta, madurez de las hojas y las yemas previo a la diferenciación (Schaffer et al. 1994, Avilán 1999, Galán 1999, Monselise y Goldschmidt 1982, Gamboa y Mora 2010). La fenología del mango se caracteriza por presentar fases bien marcadas en climas subtropicales. En los trópicos este no es el caso, ya que se produce simultaneidad de varias fases fenológicas en un mismo período (Galán 1999).
La producción de mango tiende a alternar un año de abundante cosecha (año “on”), seguido de uno o varios de menor producción (años “off”). Las plantas que presentan alternancia producen de manera no uniforme en períodos o años consecutivos, situación que parece estar relacionada con la cantidad de sustancias de reserva que se acumulen en la planta (Monselise y Goldschmidt 1982). La producción alternante puede deberse también a factores como cultivares, magnitud de la producción anterior, crecimientos vegetativos posteriores a la cosecha, estado de madurez de los brotes terminales durante la diferenciación floral, relación C/N de los ápices terminales en el momento en que se producen las condiciones ambientales conducentes a la inducción floral, estado nutricional de las plantas, y a los factores climáticos actuales y pasados a los que han estado expuestas las plantas (Schaffer et al. 1994).
El estudio del metabolismo de carbohidratos es esencial para comprender eventos como la producción de follaje, ramas, raíces, flores, y frutas, y relacionarlos con la formación, acumulación y utilización de carbohidratos de reserva. El contenido de almidón se considera el mejor indicador del nivel de carbohidratos, del estado fisiológico de los árboles y de su potencial productivo (Monselier y Goldschmidt 1982, Rasmunssen y Henry 1990, Ponchner 1992).
Las reservas almacenadas en las plantas pueden ser utilizadas en crecimiento y mantenimiento. En árboles, las raíces parecen ser el órgano donde se acumula la mayor cantidad de carbohidratos de reserva. (Wood y McMeans 1989, Loescher et al. 1990, Pochner 1992). Las plantas leñosas acumulan reservas en el período previo a la floración, que se consumen en el desarrollo de flores y frutos, y durante nuevos episodios de crecimiento vegetativo (Kozlowsky et al. 1997). Estas reservas cambian constantemente dependiendo de la redistribución de los carbohidratos, en apoyo del crecimiento (Keller y Loescher 1989, Kozlowsky 1992, Pochner 1992). La fruta es un fuerte demandante de carbohidratos y domina las otras partes de las plantas durante su desarrollo y hasta la cosecha (Stasssen y Janse van Vuuren 1997).
En ciertas regiones luego del crecimiento inicial de los frutos puede darse además un brote y crecimiento de yemas vegetativas. (Davie y Janse van Vuuren 1998). Los árboles con gran número de frutos experimentan disminución de carbohidratos de reserva al inicio de su desarrollo, antes de alcanzar su tamaño final. Tras la recolección, se empiezan a recobrar las reservas de almidón en raíces y madera lo que puede variar un poco dependiendo de la fenología particular de cada árbol (Davie et al. 1995).
Durante el cuajado y la fase de desarrollo de la semilla se consume la mayoría de los carbohidratos disponibles, lo que resulta en la inhibición temporal de los crecimientos vegetativos (Kozlowsky 1997). La renovación de las reservas es esencial si se desea una producción adecuada en el siguiente período (Davie et al. 2000). Luego de la cosecha el uso de carbohidratos de reserva es bajo, con la excepción de lo utilizado en el crecimiento de raíces (Davie y Janse van Vuuren 1998).
En estudios realizados en Australia, Rasmunssen y Henry (1990) reportaron concentraciones de almidón en árboles de mango con valores que fluctúan entre el 0,6 y 22,3% en tallos, y entre 2,9 y 33,3% en raíces de árboles de mango. Estudios realizados en Sudáfrica en mango han contribuido al diseño de un modelo del ciclo de carbohidratos y valores críticos de almidón para el mango en los subtrópicos (Robert y Wolstenholme 1993). Según este modelo se produce una acumulación de reservas en las raíces y madera del árbol en los meses previos a la floración. Posterior a la floración y cuajado de frutos, se produce una disminución de las reservas (Galán 1999). Según Davie y Stassen. 1997a, b y Davie et al. 2000, las reservas de almidón de las raíces son las más móviles, pues se consumen más rápidamente que las de la madera hasta alcanzar valores mínimos. Otro estudio en Sudáfrica registró una mayor acumulación de almidón en la zona del árbol comprendida entre las raíces y las ramas primarias (Robert y Wolstenholme 1992). Por otra parte Schroeder (1992) en una investigación en la variedad Sensation menciona que las reservas de carbohidratos aparentemente son almacenadas mayormente en la madera del tronco.
El fenómeno de alternancia o bianualidad en la producción en mango puede deberse principalmente por el agotamiento de las reservas de almidón durante la producción de fruta y el desarrollo vegetativo, ya que las hojas existentes no pueden reponer rápidamente las reservas para suplir las demandas de un nuevo ciclo de floración y desarrollo de los frutos (Davie y Stassen 1997a). Esto puede ocasionar que luego de la cosecha de un año dado, el árbol no disponga de suficientes reservas para una buena producción el año siguiente. Estos bajos niveles en las reservas de carbohidratos pueden conducir a la instauración del ciclo “on-off” de producción, con un año (o varios) de muy baja o nula producción (año “off”) seguido de otro de cosecha media a alta con muchos frutos generalmente de menor tamaño (año “on”) (Singh 1968, Davie y Janse van Vuuren 1998).
Se han reportado diferencias en rendimientos del mango entre 3 a 45 toneladas por hectárea para diferentes áreas geográficas (Moncur et al. 1985, Robert y Wolstenholme 1992, Galán 1999, Yahía et al. 2006). Los bajos rendimientos de 5 toneladas por hectárea reportados por Gillot (1997) en Costa Rica, justifican la necesidad de una mayor investigación para aumentar esos rendimientos.
El objetivo del presente trabajo fue determinar la variación estacional de los contenidos de almidón y su relación con patrones fenológicos y de producción en árboles de mango de las variedades Tommy Atkins y Keitt en condiciones tropicales.
Materiales y Métodos
Este estudio se realizó en una finca del Instituto Nacional de Aprendizaje (INA) de Costa Rica, ubicada en Liberia, Guanacaste a 100 msnm, a 10o 36’ de latitud norte y 85o 32’ longitud oeste. La zona presenta una temperatura media de 27°C, una precipitación promedio anual de 1507,1 mm, y una humedad relativa promedio anual de 75,1% (Instituto Meteorológico Nacional 2002).
Se utilizaron las variedades comerciales Tommy Atkins y Keitt. Ambas se injertaron sobre el patrón “Jamaica” o “mango mecha” (Araya 2002). Los árboles se escogieron siguiendo criterios de homogeneidad y sanidad. Se utilizaron para el estudio tres árboles por cada variedad. Las plantas se sembraron en 1990, por lo que al inicio tenían aproximadamente trece años, y se encontraban en el período de plena producción (Araya 2002, Avilán 1999, Gamboa y Mora 2010). Los árboles recibieron podas de mantenimiento poscosecha, fertilización, control de malezas, combate de enfermedades y de plagas. No se realizaron aplicaciones de productos con efecto hormonal que interfieran con el desarrollo fenológico normal, excepto la inducción floral con nitrato de potasio (KNO3) (Gamboa 2008).
En las plantaciones se utilizó nitrato de potasio como un inductor de la floración (Nuñez-Elisea 1986, Guzmán 1996, Gamboa y Mora 2010), con un contenido del 13% de nitrógeno y 44% de potasio. El momento de la aplicación del producto varió según las condiciones de clima, de la madurez del tejido foliar y yemas de los árboles. Se aplicó el nitrato de potasio al follaje en solución, a una concentración de 4,2%. En promedio se realizaron de dos a tres aplicaciones de diciembre a enero, en los ciclos productivos del 2004, 2005 y 2006.
Fenología y clima
Los árboles se evaluaron durante dos años y medio y se les realizó una estimación cualitativa del porcentaje de cada estado fenológico por cuadrante por árbol, en un ámbito de 0 a 100%, basada en las metodologías de Fournier (1974) y Villalpando y Ruiz (1993). Los estados fenológicos evaluados fueron: brotación de yemas vegetativas, brotación de flores y la producción de frutos. Se realizaron determinaciones quincenales de febrero del 2004 a julio del 2006, que se utilizaron para obtener el porcentaje de cada estado fenológico. Es importante señalar que las observaciones se efectuaron a partir de febrero del 2004, por lo que posiblemente la floración de ese año fue subestimada debido a que la época de mayor floración por lo general se presenta en los meses inmediatamente anteriores (enero y diciembre). La medición de la cosecha se obtuvo con base en el número y peso de las frutas, cosechadas en su punto de madurez fisiológica. Se contabilizó la producción por planta y por variedad del 2004 al 2006 (tres cosechas).
La información de las variables climáticas se obtuvo de la Estación Meteorológica del Aeropuerto Daniel Oduber de Liberia, ubicada a unos 10 km del área del experimento. Las variables climáticas registradas fueron temperatura y precipitación. Además se utilizaron datos promedios mensuales de brillo solar y radiación (Instituto Meteorológico Nacional 2006).
Determinación de los contenidos de almidón
Se tomaron muestras de aproximadamente 100 g de peso fresco de raíces, tronco y ramas de árboles de cada una de las variedades. Las muestras de las diferentes partes del árbol se obtuvieron mediante cortes de aproximadamente 1 cm de profundidad de la madera. Las muestras de raíces se tomaron de raíces secundarias con un diámetro mayor a 2 cm. Para el tronco se tomó tejido de la parte superior del injerto y para las ramas se utilizaron ramas secundarias y terciarias mayores de 10 cm de diámetro. La colecta de las muestras de madera se efectuó en febrero, mayo, agosto y noviembre del 2004 y 2005. Se escogieron esos meses en función de los estados fenológicos esperados (Gamboa 2008). Las muestras se secaron en una estufa a 60-65°C durante 3-6 días; luego se pulverizaron en un molino de mallas de 1mm. Posteriormente se determinó la concentración de almidón en el laboratorio del Centro de Investigación en Tecnología de Alimentos de la Universidad de Costa Rica. El análisis se realizó mediante hidrólisis enzimática con α-amiloglucosidasa y oxidación de la glucosa resultante (E.C. 3.2.1.3 y EC 1.1.3.4) (Southgate 1976, Bruce y Dickson 1979, Rasmunssen y Henry 1990). La glucosa se determinó con un espectrofotómetro de doble rayo a 505 nm. También se realizó un análisis de la muestra sin la enzima donde se determinó la glucosa de fondo que no provino de la hidrólisis del almidón. Se obtuvo un promedio del contenido de almidón por árbol para cada variedad a partir de las muestras de cada parte del árbol.
Resultados y Discusión
Clima
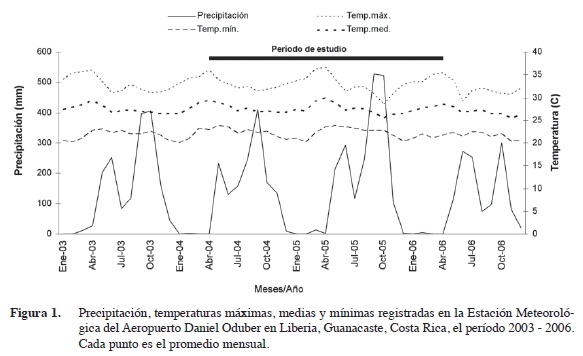
La zona donde se realizó el estudio tiene una época seca y una lluviosa. Sin embargo esta última se interrumpe entre julio y agosto cuando ocurre una disminución de la precipitación (Figura 1).
Las temperaturas máximas ocurrieron en marzo y abril con valores entre 30 y 35°C. Los valores mínimos estuvieron entre los 20 y 25°C. La temperatura media fue cercana a los 27°C durante todo el periodo de estudio. La media acumulada de precipitación de los registros de 44 años (período 1957-2001) es de 1507,1 mm. En cada año alterno del período de estudio, la precipitación fue superior o inferior a la media acumulada de los registros. En el 2003 y 2005 llovió más que el promedio acumulado (1703 mm y 2049mm respectivamente), mientras que, en el 2004 y 2006 la precipitación fue menor (1500 mm y 1217 mm respectivamente). Durante el año se presentaron dos picos de precipitación, uno entre mayo-junio y otro entre setiembre y octubre, con valores de 232 mm y 411 mm en mayo y octubre del 2004 respectivamente; 294, 528 y 524 mm en junio, setiembre y octubre del 2005; 272 y 254 mm en junio y julio y 299 mm en octubre del 2006 (Figura 1).
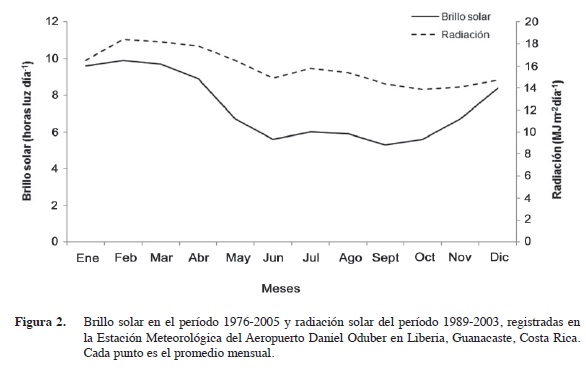
Los datos diarios de brillo y radiación solar corresponden a los promedios de los períodos 1976-2005 y 1989-2003 respectivamente. La zona de Liberia presenta un mayor número de horas luz de diciembre a abril de cada año, con un promedio de nueve horas por día. Por otra parte, el menor número de horas luz se presenta entre junio y setiembre con seis horas (Figura 2). En esta zona se obtiene mayor radiación entre febrero y abril con valores de 18 MJ m-2día-1. Durante la época lluviosa de setiembre a noviembre esta radiación disminuye a valores cercanos a 14 MJ m-2día-1. La radiación promedio anual es de 15,9 MJ m-2día-1.
El clima registrado en el período del estudio fue en términos generales adecuado para el desarrollo del árbol de mango (Moncur et al. 1985). Dadas las condiciones de estacionalidad en la zona donde se hizo la investigación, se presenta un déficit hídrico de diciembre a mayo y esta época coincide con la fase de floración y desarrollo de frutos (Araya 2002). En climas tropicales la temperatura presenta poca variación durante el año. Bajo estas condiciones la cantidad de agua del suelo es uno de los factores más influyentes en la fenología. (Schaffer et al. 1994, Galán 1999).
Fenología
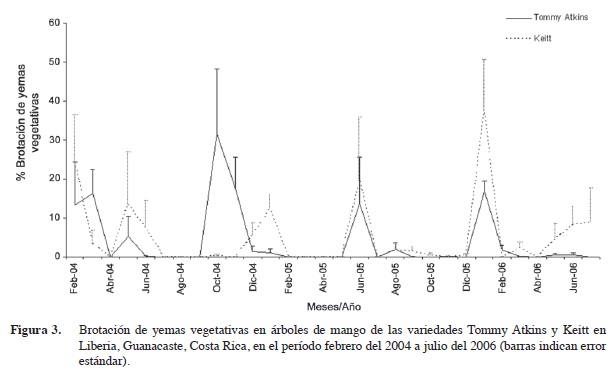
Las observaciones fenológicas se realizaron durante 29 meses para contemplar al menos dos ciclos anuales. Los árboles de la variedad Tommy Atkins presentaron los mayores porcentajes de yemas vegetativas brotadas en marzo y octubre del 2004 (17% y 32% respectivamente); en junio del 2005 (14%); y en enero del 2006 (17%). Por otra parte los árboles de la variedad Keitt presentaron máximos eventos de crecimiento vegetativo en febrero y mayo del 2004 (24% y 14% respectivamente); en enero y junio del 2005 (12 y 20% respectivamente), y en enero del 2006 (37%). En las dos variedades se observaron flujos de crecimiento vegetativo en momentos similares, excepto en el 2004, año en el que el crecimiento fue mayor en la variedad Tommy Atkins (Figura 3). En 2005 y 2006 los flujos de crecimiento ocurrieron en épocas similares en las dos variedades, pero fueron de mayor intensidad en la variedad Keitt.
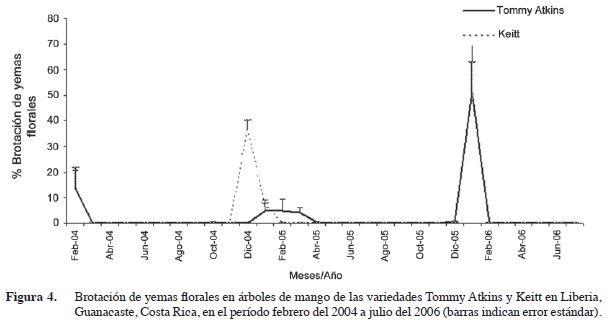
Los picos de brotación de yemas florales en la variedad Tommy Atkins se observaron en febrero del 2004 (14%) y en enero del 2006 (51%). En el 2005 se observó una floración baja entre enero y marzo con promedios mensuales de 4 a 5%. Los picos de brotación de la variedad Keitt se observaron en febrero (14%) y diciembre del 2004 (36%) y en enero del 2006 (49%) (Figura 4). Estos eventos se presentaron con más regularidad que los vegetativos, generalmente en diciembre y enero de cada año. La intensidad de los flujos de crecimiento reproductivo de las dos variedades fueron similares a inicios del 2004. Sin embargo a fines de ese año, la variedad Keitt registró una floración mayor que la Tommy Atkins. Durante el 2005 no se observó desarrollo floral significativo en ninguna de las dos variedades. En enero del 2006 la intensidad de la floración fue similar en ambas variedades y además se observó una alta brotación de yemas vegetativas y florales en las dos variedades en ese momento.
Producción
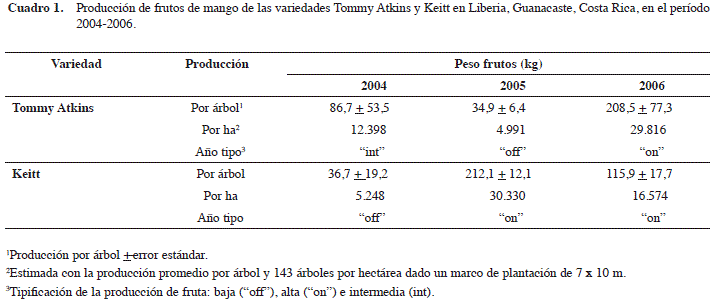
En la variedad Tommy Atkins la cosecha en el 2005 fue la más baja del período estudiado, con 74 frutos y 34,9 kg por árbol; mientras que en el 2006 se registró la mayor producción con 414 frutos y 208,5 kg. Por otra parte la variedad Keitt presentó la cosecha más baja en el 2004 con 87 frutos y 36,7 kg por árbol. La cosecha del 2005 fue la mejor con 462 frutos y 212,1 kg. A partir de los datos de cosecha por árbol y densidad de plantación, se estimó la producción por unidad de área (Cuadro 1). Se tipificó la producción de cada año como baja (“off”), alta (“on”) e intermedia (int), al compararlas con lo reportado por la literatura.
Los resultados del estudio fenológico mostraron una fuerte floración en la variedad Keitt en diciembre 2004 que se reflejó en una alta producción de frutos en el 2005, a esto se suma el hecho de que esta variedad venía de un año (2004) de producción baja. Por otra parte la ausencia de crecimientos vegetativos en la segunda mitad del año fue un factor adicional que pudo contribuir a la alta producción en el 2005 en esta variedad. De esta manera se dieron condiciones para una adecuada maduración de tejidos y diferenciación floral generando la producción más alta del período estudiado.
En contraste, en la variedad Tommy Atkins no se observó floración en diciembre 2004 y posteriormente registró valores muy bajos de enero a marzo de 2005, lo que podría explicar parcialmente la baja producción de frutos en esta variedad en ese año. Es importante además notar que la variedad Tommy Atkins, presentó eventos de crecimiento vegetativos con valores altos en los meses inmediatamente anteriores a la floración (octubre y noviembre 2004), lo que probablemente contribuyó a la baja producción de frutos en el 2005. El tejido vegetativo que brotó entre octubre y noviembre probablemente no alcanzó la suficiente madurez para una óptima diferenciación floral. Esta condición la señala Monselise y Goldschmidt (1982) y por Schaffer et al. (1994) como requisito para alcanzar la diferenciación floral.
Otro factor asociado a una baja producción de la variedad Tommy Atkins en el 2005 pudo ser el hecho de que el año anterior (2004) la zona en estudio registró una precipitación de 1500 mm, la cual puede considerarse relativamente baja para las necesidades de los árboles de esta variedad. También hay que considerar que venía de un año (2004) de producción relativamente alta, lo que podría contribuir a la baja producción en el 2005 dada la condición aparentemente alternante de esta variedad (Cuadro 1)
En enero del 2006 las dos variedades mostraron altas floraciones, la variedad Keitt registró una buena producción en ese año, pero no muy alta. Esto podría explicarse por diversos factores: en primer lugar esta variedad venía de un año de máxima producción en 2005, año que también registró una buena precipitación lo que le permitió una recuperación a los árboles de esta variedad. Por otra parte se presentó un fuerte crecimiento vegetativo en enero del 2006, simultáneo con la floración. Esta sincronización de eventos pudo afectar en algún grado el desarrollo de frutos a pesar de la relativa buena producción dada la alta floración de ese año.
Por otra parte la variedad Tommy Atkins en el 2006 presentó la máxima producción registrada del período de estudio. Esta alta producción pudo deberse a varios factores: en primer lugar esta variedad venía de un año de baja producción en 2005 (la mínima del período de estudio), sumado a una precipitación adecuada en ese año y un crecimiento vegetativo moderado y simultáneo con la floración en enero 2006.
En el año 2006 la variedad Tommy Atkins presentó la mayor producción, mientras que la Keitt fue en el 2005. En contraste, la menor producción para estas variedades se registró en el 2005 y el 2004 respectivamente. Los años de mayor producción en las dos variedades pueden corresponder a años “on”, los de menor producción a años “off” y el 2004 en la variedad Tommy Atkins puede catalogarse como intermedio.
La alternancia en la producción puede deberse a factores endógenos de la planta. Entre ellos la inhibición de la iniciación floral causada por problemas hormonales y nutritivos; por problemas de polinización; la competencia entre tejidos vegetativos y reproductivos; y el agotamiento de reservas producto de cosechas altas de los años “on”, la que es considerada como una de las principales causas de alternancia (Monselise y Goldschmidt 1982).
Araya (2002) en un estudio que llevó a cabo en el período 1995-2000 en la misma plantación donde se realizó el presente trabajo, obtuvo en el año 2000 rendimientos de 15,4 y 26,1 t/ha de fruta, en las variedades Tommy Atkins y Keitt respectivamente, que fueron los valores más altos obtenidos en ese estudio. Los árboles en ese momento tenían ocho años de edad y venían de un período de recuperación de los efectos del evento del Niño ocurrido en el periodo 1997-1998. Según nuestros resultados esos valores corresponden a niveles de producción intermedia y “on” respectivamente.
Contenido de almidón
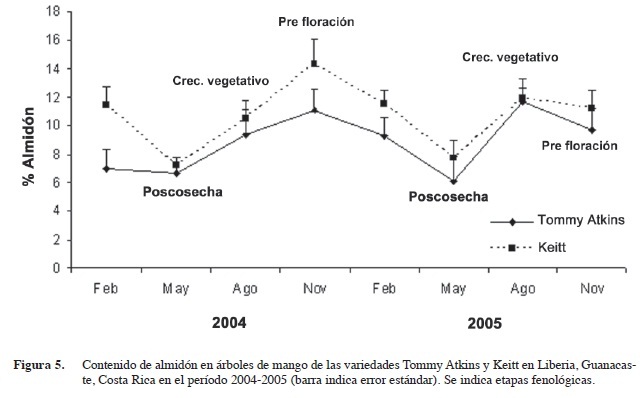
El promedio del contenido de almidón por variedad obtenido en las diferentes épocas mostró mínimos en mayo del 2004 y 2005, con valores de 6,7% y 6,1% en Tommy Atkins y de 6,1% y 7,7% en la variedad Keitt. Por otra parte se registraron valores máximos en noviembre del 2004 con 11,1% en Tommy Atkins y 14,4% en Keitt; mientras que en agosto del 2005 Tommy Atkins presentó 11,7% y Keitt 11,9% (Figura 5). Ambas variedades presentaron la misma tendencia, pero con niveles superiores de almidón en la variedad Keitt en todos los muestreos. Las concentraciones más bajas de almidón ocurrieron en los períodos poscosecha (mayo 2004 y mayo 2005), y los máximos en prefloración (noviembre 2004), y en un período de reducido crecimiento vegetativo (agosto 2005).
En condiciones tropicales, y en particular para el caso de Costa Rica no se dispone de información del manejo de reservas de carbohidratos en árboles de mango. El conocimiento de cómo funcionan los árboles de mango es indispensable para obtener una óptima calidad y cantidad de fruta (Davie y Janse van Vuuren 1998). Esta información es muy importante para determinar mejoras a las condiciones de manejo de los árboles con prácticas agronómicas adecuadas (Robert y Wolstenholme 1992, Davie y Janse van Vuuren 1998).
En este estudio, el contenido de almidón presentó valores que se ubican dentro del ámbito reportado en la literatura para este cultivo (Rasmunssen y Henry 1990 y Galán en 1999). En términos generales el contenido de almidón alcanza niveles máximos en noviembre de cada año, lo que corresponde al estado fenológico de prefloración. Mientras que los niveles de almidón más bajos se registraron en mayo de cada año correspondiente a la fase poscosecha. Estos resultados están de acuerdo a lo reportado por la literatura, en el sentido de que las plantas leñosas acumulan reservas en prefloración y que posteriormente las consumen en el desarrollo de flores, frutos y nuevos crecimientos vegetativos (Davie y Janse van Vuuren 1998). La demanda de carbohidratos es muy alta en el desarrollo de los frutos por lo que probablemente los árboles en producción, tuvieron un acelerado consumo de las reservas de almidón en esos períodos (Kozlowsky 1992, Koslowsky et al. 1997, Davie et al. 1995, Stasssen y Janse van Vuuren 1997). Una vez finalizada la fructificación (mayo de cada año) los niveles de almidón llegan a sus valores mínimos y luego empiezan a aumentar. Esto concuerda con lo reportado por Davie et al. 1995, Robert y Wolstenholme 1992, Robert y Wolstenholme 1993, Davie et al. 2000. En los trópicos el consumo de las reservas se debe a los gastos generados por crecimientos vegetativos aéreos y radicales (Kozlowsky et al. 1997), pero muy probablemente el mayor consumo se deba a la producción de flores y frutos.
Con relación al uso de las reservas, es importante señalar que el crecimiento de la raíz no se evaluó en el presente estudio. La raíz utiliza una gran cantidad de carbohidratos de reserva (Kozlowsky 1992 y Kozlowsky et al. 1997). El crecimiento podría ser promovido al inicio del período lluvioso en mayo-junio, en las zonas donde se cultiva mango sin riego. Además ese momento corresponde con los niveles bajos de reservas de almidón por el gasto realizado previamente en el crecimiento de los frutos, lo que podría afectar el desarrollo de las raíces.
La mayoría de los estudios consultados sobre niveles de almidón en árboles de mango consideran datos de solo un ciclo anual y en zonas subtropicales. Esto representa una limitante al considerar la alternancia en la producción que presenta el mango, ya que es recomendable disponer de datos de al menos dos ciclos de producción. El período poscosecha parece ser el momento más adecuado para determinar los niveles de almidón; ya que este momento podría indicar el nivel de consumo de estas reservas para la producción de fruta. La literatura no menciona cual es el nivel adecuado de reservas previo a la floración, ni tampoco indica cual sería un nivel de consumo alto o moderado de las reservas en el período poscosecha. Estos podrían ser algunos de los aspectos a considerar para la evaluación de tendencias de producción.
De acuerdo a los resultados de esta investigación, una buena producción (año “on”) es precedida por altos niveles de almidón en la fase de prefloración de esa cosecha. Esta tendencia la muestran las dos variedades estudiadas, sin embargo la variedad Keitt en todos los períodos mostró niveles más altos de almidón lo que podría estar relacionado con un mayor potencial productivo de esta variedad. Esto concuerda con lo expresado por Ryugo et al. 1977, Monselise y Goldschmidt 1982, y Tromp 1983. Por otra parte Davie y Janse van Vuuren (1998) reportaron que el contenido de almidón en árboles de mango con alternancia, mostraron un pico alto cerca del momento de la formación del botón floral, para luego decrecer gradualmente. Además encontraron que esta alta concentración fue mayor en los años de fuerte producción, comparado con un período de menor producción. Sin embargo bajo las condiciones tropicales en que se realizó la presente investigación no se encontró una relación clara entre niveles de almidón previos a la floración y el nivel de producción obtenido en ambas variedades. Por otro lado, el bajo número de árboles utilizados en los muestreos, debido al costo del análisis de almidón, puede considerarse una limitante de esta investigación.
Los resultados sugieren, que la variedad Keitt posee mayor potencial productivo, comparado con la Tommy Atkins, lo que concuerda con lo reportado por Araya (2002) en la misma zona de estudio. Sin embargo el uso de una variedad particular en una zona determinada está definido no sólo por su potencial productivo o genético, sino que también intervienen factores de clima, manejo agronómico, mercado y costos que hay que considerar. Los patrones fenológicos observados en este estudio en ambas variedades podrían interpretarse como no óptimos desde el punto de vista productivo, ya que en algunos de los años se presenta poco crecimiento vegetativo al inicio de la época lluviosa y en otros abundantes crecimientos en la segunda mitad del año. Esto puede deberse entre otros factores, que en la zona de estudio se presenta un período seco de extensión variable a mitad del año, que puede inducir cierto grado de maduración de las yemas vegetativas presentes. El reinicio del período lluvioso en setiembre y octubre afecta las yemas mencionadas, generando un descontrol en el patrón de desarrollo y crecimiento de los árboles de mango. Esto puede hacer que se produzca poco tejido vegetativo de adecuada madurez a fines de año y el gasto de reservas en crecimiento en la segunda mitad de año, que puede resultar en una baja producción de frutos. El manejo de las condiciones de la plantación, tales como poda, fertilización, anillado, riego, y la aplicación de reguladores del crecimiento vegetal, pueden contribuir a mejorar el potencial productivo de las variedades. Asimismo la selección de patrones que controlen el vigor vegetativo de los árboles y el uso de materiales más adaptados a las condiciones tropicales pueden también contribuir a controlar excesivos flujos de crecimiento vegetativo en momentos inadecuados y a promover una mayor floración y producción de frutos.
Agradecimientos
Los autores agradecen al Instituto Nacional de Innovación y Transferencia en Tecnología Agropecuaria (INTA) de Costa Rica por el apoyo financiero y logístico para la realización de este trabajo. Al Instituto Nacional de Aprendizaje (INA) por la colaboración brindada en el uso de la plantación de mango. Al Dr. José Fermín Subirós R. por sus valiosas sugerencias al manuscrito y apoyo en esta investigación.
Literatura Citada
Araya, A. 2002. Estudio integral de cuatro variedades de mango (Mangifera indica L.) con dos densidades de plantación en Liberia Guanacaste. Práctica dirigida Lic. Ing. Agr, Heredia, CR, Universidad Nacional, 101 p. [ Links ]
Avilán, L. 1999. Fertilización del mango en el Trópico. Informaciones agronómicas N.34. Instituto de la potasa y el fósforo. Canadá. 6 p. [ Links ]
Bruce H; Dickson, R. 1979. Starch measurement in plant tissue using enzimatic hydrolysis. Physiol. Plant. 47:151-157. [ Links ]
Davie, SJ; Martie Van Der Walt; Stassen, PJC. 1995. The energy demand for fruit production of Sensation mangoes and its effect on fruit size. S. A. Mango Growers’ Assoc. Yearbook 15:18-20. [ Links ]
Davie, SJ; Stassen, PJC. 1997a. Mango model: starch distribution of different tissues on “Sensation” mango trees of varying ages. Acta Horticulturae 455 (1):143-150. [ Links ]
Davie, SJ; Stassen, PJC. 1997b.The effect of fruit thinning and tree pruning on tree starch reserves and on fruit retention of “Sensation” mango trees. Acta Horticulturae 455(1):161-167. [ Links ]
Davie, SJ; Janse Van Vuuren, BPH. 1998. Phenology and Physiology. In Villiers E.A. ed. The Cultivation of Mangoes. Nelspruit. Sudáfrica, Institute for Tropical and Subtropical Crops. p. 27-30. [ Links ]
Davie, SJ; Stassen, PJC; Grove, HG. 2000. Starch reserves in the mango tree. Acta Horticulturae 509(1):335-346. [ Links ]
Fournier, LA. 1974. Un método cuantitativo para la medición de características fenológicas en árboles. Turrialba 24(4):422-423. [ Links ]
Galán, V. 1999. El cultivo del mango. Ediciones Mundiprensa. Madrid. 297 p. [ Links ]
Gamboa, JR. 2008. Variación estacional en el contenido de almidón y minerales en árboles de Mangifera indica L (mango) en Liberia Guanacaste. Tesis para optar por el título de Magister Scientiae en Biología. Universidad de Costa Rica. 104 p. [ Links ]
Gamboa, J & J Mora. 2010. Guía para el cultivo del mango (Mangifera indica L) en Costa Rica. Ministerio de Agricultura y Ganadería. San José, CR. 61 p. [ Links ]
Gillott, R. 1997. Cultivo del mango en Costa Rica. Consejo Nacional de Producción. DRPC N. 391-97. Barranca, Puntarenas. 4 p. [ Links ]
Guzmán, C. 1996. Modifying the harvesting period of mango implications and perspectives. IICA. Tropical Fruits Newsletter 20:10-11. [ Links ]
IMN (Instituto Meteorológico Nacional, CR). 2002. Descripción del clima de Liberia. San José, Costa Rica. 23 p. [ Links ]
IMN (Instituto Meteorológico Nacional, CR) 2006. Boletínes Meteorológicos Mensuales (en línea). San José. Consultados 7 marzo 2007. Disponibles en http://www.imn.ac.cr/boletines/index.html. [ Links ]
Keller JD; Loescher, WH. 1989. Nonstructural carbohydrate partitioning in perennial parts of sweet cherry. J. Amer. Soc. Hort. Sci. 114 (6): 969-975. [ Links ]
Kozlowsky, TT. 1992. Carbohydrate Sources and Sinks in Woody Plants. Bot. Rev. 58: 107-222. [ Links ]
Kozlowsky, TT; Pallardy, SG; Kramer, PJ. 1997. Physiology of woody plants. Academic Press. San Diego, California. 411 p. [ Links ]
Loescher, WH; McCamant, T; Keller, JD. 1990. Carbohydrate reserves, translocation, and storage in woody plant roots. HortScience 25(3):274-281. [ Links ]
Moncur, M; Winston, EC; Wrigth, RM. 1985. Tabular descriptions of crops grown in the tropics. Mango (Mangifera indica L.). Technical memorandum 85/17. Institute of biological resources, division of water and land resources, Canberra. Australia. 49 p. [ Links ]
Monselise SP; Goldschmidt, EE. 1982. Alternate bearing in fruit trees. Hort. Rev. 4:128-173. [ Links ]
Nuñez-Elisea, R. 1986. Producción temprana de mango “Haden” y “Manila” con aspersiones de nitrato de potasio. Secretaría de Agricultura y Recursos Hidráulicos SARH. Folleto para productores N.8. Tecomán, México. 9 p. [ Links ]
Ponchner, S. 1992. Carbohydrate partitioning to roots of “French Prune” trees as affected by crop load and potassium nutrition. Thesis Master of Science. University of California Davis. 81 p. [ Links ]
Rasmunssen, T; Henry, RJ. 1990. Starch determination in horticultural plant material by an enzimic-colorimetric procedure. J. Sci. Food Agric. 52:159-170. [ Links ]
Robert, JP; Wolstenholme, BN. 1992. Phenological cycles, carbohydrates status, and CPPU spray trials for four mango cultivars-current research in the Nkwalini Valley, Natal. South Africa Mango Growers’ Association Yearbook 12:9-13. [ Links ]
Robert, JP; Wolstenholme, BN. 1993. Phenological cycles and seasonal fluctuations of three mango cultivars: an update on current research in the Nkwalini Valley, Natal. South Africa Mango Growers’ Association Yearbook 13:2-10. [ Links ]
Ryugo, K; NII, N; Iwata, M; Carlson, RM. 1977. Effect of fruiting on carbohydrate and mineral composition of stems and leaves of french prunes. J. Amer. Soc. Hort. Sci. 102:813-816. [ Links ]
Singh, LB. 1968. The mango, botany, cultivation, and utilization. Leonard Hill, London. 439 p. [ Links ]
Schaffer, B; Whiley, A; Crane, J. 1994. Mango. In Schaffer, B; Andersen, P. eds. Handbook of environmental physiology of fruit crops, volume II: Sub tropical and tropical crops. Florida. p. 165-197. [ Links ]
Schroeder, W. 1992. Phenological cycle of the mango cultivar Sensation at Nelspruit and the implications for management. S.A. Mango Growers’ Assoc. Yearbook 12:43-44. [ Links ]
Southgate, DAT. 1976. Determination of food carbohydrates. University of Cambridge and Medical Research Council, England. 178 p. [ Links ]
Stasssen, PJC; Janse Van Vuuren, BPH. 1997. Storage, redistribution and utilization of starch in young bearing “Sensation” mango trees. Acta Horticulturae 455(1): 151-159. [ Links ]
Tromp, J. 1983. Nutrient reserves in roots of fruit trees, in particular carbohydrates and nitrogen. Plant and Soil. 71:401-413. [ Links ]
Wood, BW; McMeans. JL. 1989. Carbohydrate changes in various organs of bearing and nonbearing pecan trees. J. Amer. Soc. Hort. Sci. 106(6):758-761. [ Links ]
Villalpando, JF; Ruiz, JA. 1993. Observaciones agrometeorológicas y su uso en la agricultura, UTEHA, México. 134 p. [ Links ]
Yahía, EM; Ornelas Paz, J de J; Flores, RA. 2006. El mango. Editorial Trillas. México. 224 p. [ Links ]
*Correspondencia a:
Jimmy R. Gamboa-Porras. Instituto Nacional de Innovación y Transferencia en Tecnología Agropecuaria (INTA). jgamboa@inta.go.cr
Walter Marín-Méndez. Escuela de Biología, Universidad de Costa Rica. walter.marin@ucr.ac.cr
1. Este trabajo forma parte del contenido de la tesis de maestría del autor principal para optar por el título de Magister Scientae en Biología.
2. Instituto Nacional de Innovación y Transferencia en Tecnología Agropecuaria (INTA). jgamboa@inta.go.cr
3. Escuela de Biología, Universidad de Costa Rica. walter.marin@ucr.ac.cr
Recibido: 21 de julio, 2011. Aceptado: 12 de marzo, 2012


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}