











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction.
Malaxis Sol. ex Sw. (Epidendroideae: Malaxideae: Malaxidinae) in its traditional circumscription is a cosmopolitan genus comprising ca. 300 species. It has a broad pantropical distribution and few species in the temperate regions of the Americas, Asia, and Europe (Cribb 2005). However, preliminary molecular phylogenetic analyses of Malaxidinae suggest that Malaxis in a broad sense is polyphyletic and that the genus in the strict sense may rather be restricted to America (Cameron 2005, Radins et al. 2014). In the Americas, there are 143 species of Malaxis sensu lato currently recognized; Mexico harbors the highest species diversity with 71 published species, 45 of them endemic, followed by Costa Rica with 22 species, six of them endemic, and then Panama with 17 species, two of them endemic (Bernet et al. 2021, Chinchilla 2019, Chinchilla et al. 2020a, Dodson 2002, Dressler 2003, 2009, Espejo Serna et al. 2002, Flora do Brasil 2021, González Tamayo et al. 2008, Ulloa Ulloa et al. 2018 onwards, Villaseñor 2016). Nonetheless, as detailed taxonomic studies on Malaxis are carried out in local and regional floras, the species richness increases in countries where the genus has previously been underestimated, such as Colombia, Ecuador, and Peru (Chinchilla 2019).
In Costa Rica, species of Malaxis grow at elevations from 200 to 3650 m, in tropical wet to subalpine pluvial forests. However, the highest species diversity is found between 1000 and 2800 m in the country (Chinchilla 2019, Chinchilla et al. 2020a). The montane forests of the main mountain ranges of Costa Rica, especially the Talamanca mountain range, harbor the greatest diversity and endemism of Malaxis (Chinchilla 2019, Chinchilla et al. 2020a, Dressler 2003).
Despite that numerous botanical expeditions have been carried out in different locations of Costa Rica, there are still little explored areas where new or unrecorded species of orchids may be harbored (Bogarín 2011, Bogarín et al. 2020, Chinchilla et al. 2020b, Fernández et al. 2014, Karremans & Díaz-Morales 2016, Karremans et al. 2012). To update floristic inventories and contribute to the knowledge of Malaxis, we have carried out botanical explorations in these less explored areas to collect, document, and identify their species (Chinchilla 2019). In the Talamanca mountain range, specimens of Malaxis were collected that did not match any of the previously reported species in the country. The examination of herbarium specimens revealed they coincide with the Panamanian Malaxis pittieri (Schltr.) Ames, which is here recorded for the first time in Costa Rica. Additionally, a Costa Rican specimen of Malaxis kept at the United States National Herbarium (US), does not match any of the species described for the genus and is described here.
Materials and methods
This study was carried out between 2015 and 2019. It is based on collections from Costa Rica and Panama. Living plants of M. pittieri collected in Costa Rica were photographed using a Nikon® 7100 camera, and the herbarium specimens were deposited at JBL and USJ. Sketches of M. pittieri were prepared from fresh material, while sketches of M. excentrica were prepared using rehydrated flowers from the holotype specimen. Both were digitized and diagrammed in a composite plate using Adobe Photoshop CS6® and digitally delineated and shaded with an Apple Pencil® in Procreate application for iPad Pro® tablet (Apple Inc.). Specimens of Malaxis were examined in the following herbaria: AMES, CR, F, HLDG, JBL, MO, SEL, US and USJ (physically), and B, BM, BRIT, COLO, DAO, G, GH, GM, GOET, IBUG, K, L, M, NY, P, PMA, UCH and W (through digital images). A distribution map was prepared using the QGIS 3.8 Zanzibar program (QGIS Development Team 2019), based on satellite imagery updated to 2021 from Microsoft Bing Maps, Microsoft Corporation©, Earthstar Geographics SIO, 2021© TomTom. The locations of the examined specimens were classified according to the Holdridge life zone system (Holdridge 1967). The phenology was determined from examination of herbarium specimens and their labels. The conservation status of each species was assessed according to the methodology of the International Union for Conservation of Nature (IUCN 2019). For M. pittieri, the Extent of Occurrence (EOO) and Area of Occupancy (AOO) were estimated based on the studied specimens using geographical data with the Geospatial Conservation Assessment Tool (GeoCAT; Royal Botanic Gardens, Kew: http:// geocat.kew.org).
Taxonomic treatment
Malaxis excentrica Chinchilla, Karremans & M.A.Blanco, sp. nov. (Fig. 1).
TYPE: Costa Rica. Puntarenas: cantón Coto Brus, distrito Sabalito, Reserva de la Biosfera de La Amistad, cerca Estación Biológica Las Alturas de Cotón, 08°57'00,3''N, 082°49'56,8''W, 2000 m, 08 julio 1994, W. J. Kress & J. L. Tuxill 94-4775 (holotype: US!-3314101, barcode 00509021).
Diagnosis: Vegetatively similar to Malaxis simillima (Rchb.f.) Kuntze, but differs in having one leaf (vs. two leaves) per sympodial unit, shorter (1.0−1.2 mm vs. 2.3−3.8 mm) petals that are obovate, erect, parallel to the column (vs. linear, recurved, surrounding the ovary), the lip with triquetrous, acute, convex (vs. trifid, concave) apical margin, and the disc cavity divided (vs. non-divided) by a costa.
A terrestrial, erect, rhizomatous herb 25.3 cm tall, including the inflorescence. Roots flexuous, short, up to 4.5 cm long × 2 mm in diameter, pubescent. Pseudobulbs 32 × 13 mm, epigeous, heteroblastic, conical, separated by an ascending rhizome with two internodes each subtended by one cataphyll, 6 cm long, when immature covered with leaf sheaths becoming scarious and papyraceous with age, and two ovate, obtuse, cataphylls 30-55 × 10-20 mm, the margins free or connate up to – of their length, shedding as the pseudobulb matures. Leaves one per sympodial unit, produced from the base of the pseudobulb, long-petiolate, conduplicate, soft-textured; sheaths with the margins connate for – of their length, forming a pseudostem, 8.20 × 0.45 cm; the blade ovate, rounded, acute, the margins undulate, nine-veined, 12.2 × 6.2 cm, ascending, green. Inflorescence produced from the apex of the pseudobulb, erect, corymbose, 22.5 cm long; peduncle 21.0 × 0.25 cm, covered near the proximal half with leaf sheaths, seven-keeled; rachis 9.5 mm long. Floral bracts ovate, acute to obtuse, concave, green, 1.2-2.5 × 0.6-1.2 mm. Pedicellate ovary terete, inconspicuously keeled, with a 180-degree twist, 6-10 mm long. Flowers non-resupinate, spreading, green. Dorsal sepal elliptic, obtuse, convex, adpressed to the ovary, 2.7-3.5 × 1.0-1.7 mm, three-veined. Lateral sepals elliptic, obtuse, convex, divergent, margins recurved, 2.5-3.6 × 1.0-1.6 mm, connate at the base 0.5-0.8 mm, three-veined. Petals obovate, rounded, erect, lightly arcuate, parallel to the column, one-veined, 1.0-1.2 × 0.4-0.5 mm. Lip simple, concave, triangular, truncate, acute, papillose, fleshy, margins entire, thick, rounded, that apically converge into a triangular rim, 2.0-3.2 × 1.0-1.8 mm; the disc cavity ovate in outline, rounded, obtuse, 1.4-2.0 × 0.7-1.3 mm × 0.5-0.6 mm deep, divided by an obcuneate, compressed costa, adaxially provided with a carina, subcavities narrowly elliptic, apically delimited by a semilunate emergent thickening; apical margin triquetrous, convex, porrect, acute, 0.7-0.9 mm long. Column 0.75-1.0 × 0.6-0.7 mm, rectangular, dorsiventrally compressed, the apex ventrally four-lobed; rostellum erect, adaxially slightly convex, emarginate; stigma obcordate, concave, longitudinally bilobed, ca. 0.4 × 0.5 mm. Pollinia four in two obovoid, ventrally concave hemipollinaria, 0.50- 0.55 × 0.2-0.25 mm. Fruits unknown.
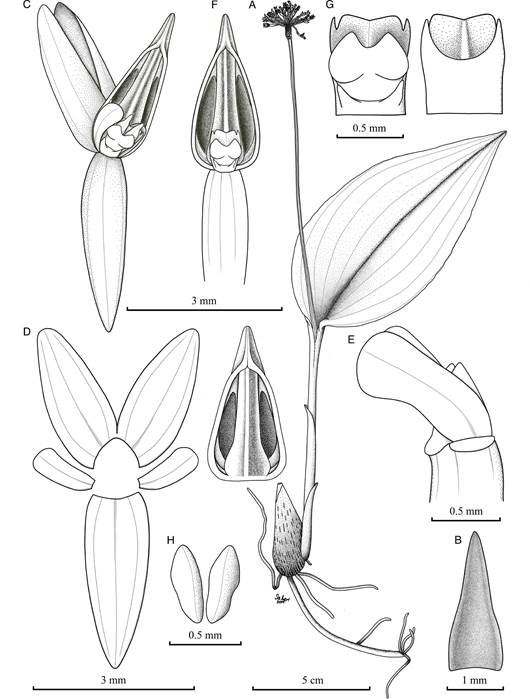
A. Habit.
B. Floral bract, adaxial view.
C. Flower, oblique view.
D. Dissected perianth (the lip on the right), flattened.
E. Apex of ovary, petals and column, lateral view.
F. Ovary, lip and column.
G. Column, dorsal (left) and ventral (right) views.
H. Pollinia.
Drawn from the holotype by Isler F. Chinchilla.
Figure 1 Malaxis excentrica.
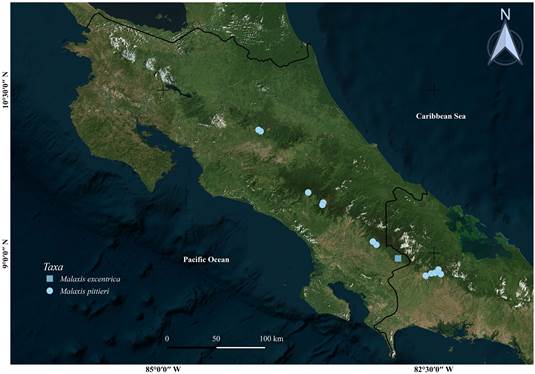
Prepared by Isler F. Chinchilla.
Figure 2 Distribution of Malaxis excentrica and Malaxis pittieri, based on the specimens examined. Satellite image source: Microsoft Bing Maps (2021).

Photographs by Isler F. Chinchilla.
Figure 3 Flowers of Malaxis simillima. From left to right: front, oblique, lateral and back views (Chinchilla et al. 3183, JBL-spirit). Scale bar = 3 mm.
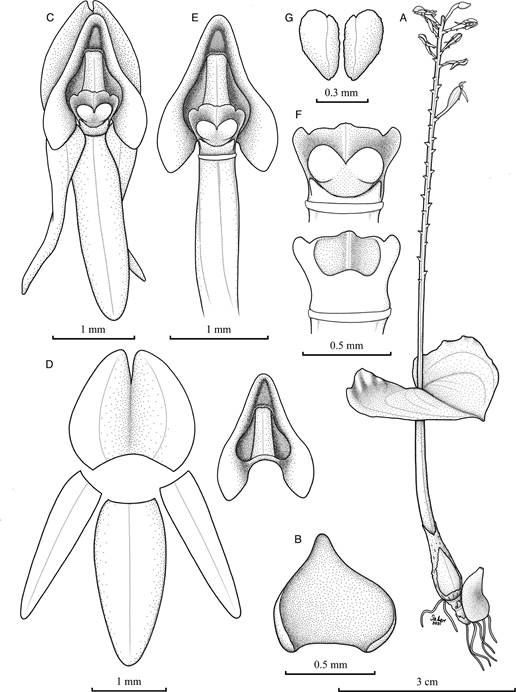
A. Habit.
B. Floral bract, adaxial view.
C. Flower, front view.
D. Dissected perianth (the lip on the right), flattened.
E. Ovary, lip and column.
F. Column, dorsal (top) and ventral (bottom) views.
G. Pollinia. Based on Chinchilla 3340 (JBL-spirit).
Illustration by Isler F. Chinchilla.
Figure 4 Malaxis pittieri.
Etymology: The specific epithet comes from the Latin excentricus, meaning eccentric or out from the center, in reference to the shape and position of petals, which are atypical in Malaxis.
Distribution and habitat: Currently known only from the type specimen collected at Las Alturas de Cotón Biological Station in Las Tablas Protective Zone, on the Pacific slope south of the Talamanca mountain range in Costa Rica (Fig. 2). It grows in the lower montane very wet forest at 2000 m in elevation.
Phenology: The only known specimen was collected in flower in July.
Malaxis excentrica has obovate, erect petals, that are parallel to the column and barely exceed the length of the column. This unique feature distinguishes the species from all other members of the genus. Furthermore, the position of the petals is reminiscent of those of species belonging to the genus Hippeophyllum Schltr., another genus of the Malaxidinae endemic to Melanesia (Cribb 2005).
Due to the ascending rhizome, conical pseudobulb, ovate leaf blade, and corymbose, compact inflorescence, M. excentrica is similar to M. simillima, but the latter has up to three sympodial units with leaves that are foveate, deeply lustrous, with undulate margins, the yellowish-green to yellow flowers, the petals incurved, arcuate, the lip deeply concave, with margins thin (vs. thick), incurved, apically with erect, thin, diverging lateral teeth, the mid-tooth porrect, thin, flat, exceeding more than twice the length of the teeth lateral, the column robust, fleshy, the rostelum deeply emarginate and the hemipollinaria narrowly (vs. obovoid) claviform (Fig. 3).
The dried flowers of Malaxis excentrica may be confused with those of Malaxis fastigiata (Rchb.f.) Kuntze, a species ranging from Mexico and Guatemala. The latter can be distinguished from M. excentrica by the longer (ca. 2.5 mm), linear, recurved petals, the lip subcordate (vs. rounded) at the base, with disc cavity occupying up to ½ (vs. more than –) of its length, divided by a pandurate (vs. obcuneate) costa, and the triangular, complanate ( vs . triquetrous, convex) apical margin.
Conservation status: Malaxis excentrica is endemic to Costa Rica, known from a single location in Las Tablas Protective Zone, Talamanca mountain range. Fortunately, the forest where it was found is conserved. Further studies on the ecology and population size of this species are required to guarantee its long-term conservation. Therefore, the species is listed as Least Concern (LC), following the IUCN (2019: section 10.4) recommendations.

A-B. Chinchilla 3338.
C. Chinchilla 3340.
D. Chinchilla 3341.
All the vouchers at JBL-spirit.
Photographs by Isler F. Chinchilla.
Figure 5 Plants of Malaxis pittieri in situ.
Malaxis pittieri (Schltr.) Ames, Proc. Biol. Soc. Washington 35: 84. 1922. (Fig. 4, 5, 6, 7). Basionym: Microstylis pittieri Schltr., Repert. Spec. Nov. Regni Veg. 12: 203. 1913.
TYPE: Panama. Chiriqui: Feuchte Wälder zwischen Alto de las Palmas und dem Gipfel des Cerro de Horqueta, 2100−2268 m ü. d. M., blühend im (11) März 1911, H. Pittier 3277 (holotype: B, destroyed (tracing of Schlechter's floral analysis of the holotype: AMES-24153/00101736 (Fig. 8A)); isotypes: AMES-00082841, a flower conserved in glycerine; US-677667/00093447, designated here as lectotype (Fig. 8B) (drawings of the lectotype: AMES24152/00101737 (Fig. 8C)); Schlechter's floral analysis from the holotype, reproduced in Mansfeld (1931: t. 17, no. 61; Fig. 8D).
A terrestrial or epiphytic, erect, subcaespitose to rhizomatous herb 4.5-20.0 cm tall, including the inflorescence. Roots flexuous, short, up to 3.5 cm long × 2.0 mm in diameter, pubescent. Pseudobulbs 7−20 × 3−10 mm, epigeous, heteroblastic, conical, separated by an ascending rhizome with two internodes, subtended by one cataphyll, 0.3-6.0 cm long, pale greenish, when immature, covered with leaf sheaths becoming scarious and papyraceous with age, and one greenish, ovate, obtuse cataphyll with inconspicuous tufts of hairs at the base, 12-40 × 5-10 mm, the margins connate, shedding as the pseudobulb matures. Leaves one per sympodial unit, produced from the base of the pseudobulb, long-petiolate, conduplicate, soft-textured; sheaths whitish-green, four-angled, the margins connate forming a pseudostem, 3.5−9.5 × 0.15−0.30 cm; the blade ovate, cordate-amplexicaul, acute, the margins undulate, five-seven-veined, 1.7−4.6 × 1.8−3.8 cm, horizontal, adaxially dark green, lustrous, abaxially light green to whitish-green, dull. Inflorescence produced from the apex of the pseudobulb, erect, racemose, lax, 4-18 cm long; peduncle 3.5−13.5 × 0.1−0.2 cm, covered up to on lower 3/4 with leaf sheaths, sometimes with a 180 degree twist, five-seven-keeled; rachis 5-65 mm long. Floral bracts ovate, acuminate, papillose, concave, greenish, pellucid, 0.5-1.5 × 0.7-1.0 mm. Pedicellate ovary terete, inconspicuously keeled, with a 360-degree twist, 3-7 mm long, white, pellucid. Flowers non-resupinate, spreading, lustrous, pellucid; sepals and petals white, lip white with a light to dark green stripe along the costa that extends to the apex of the lip, the column greenish with the operculum and rostellum white; becoming orangish with age. Dorsal sepal elliptic, obtuse, convex, adpressed to the ovary, one-veined, 2.2-2.5 × 0.8-1.0 mm. Lateral sepals asymmetrically elliptic, acute to obtuse, convex, convergent, the outer margins becoming recurved as anthesis progresses, 1.8-2.3 × 0.7-1.0 mm, the inner margins connate along 1.3−1.7 mm, one-veined. Petals narrowly lanceolate, obtuse to acute, margins revolute, recurved, arcuate, surrounding the ovary, one-veined, 2.0-2.3 × 0.3-0.5 mm. Lip trilobed, concave, sagittate, papillose, membranaceus, margins entire, thin, 1.7-2.4 × 1.2-1.6 mm; lateral lobes triangular, obtuse to acute, recurved, spreading, 0.5-0.8 × 0.4-0.5 mm; midlobe triangular, 1.4-1.7 × 0.9-1.1 mm, with the disc cavity ovate in outline, subcordate, obtuse, 0.7-1.0 × 0.4-0.8 mm × 0.3-0.5 mm deep, divided by a faint, compressed costa, basally wider, subcavities narrowly lanceolate, pellucid, apically delimited by a semilunate emergent thickening, shortly raised into a rounded margin; apical margin triangular, obtuse, concave, porrect, 0.4-0.6 mm long. Column 0.5-0.7 × 0.5-0.7 mm, rectangular, dorsiventrally compressed, the apex ventrally fourlobed; rostellum erect, truncate, bearing two viscaria at the apex; stigma transversally oblong, subcordate, concave, longitudinally bilobed, 0.15-0.25× 0.35-0.40 mm. Pollinia four in two obovoid, ventrally concave hemipollinaria, 0.30-0.50 × 0.15-0.30 mm. Fruits ellipsoid capsules, 3.0-8.0 × 1.5-3.0 mm.
Etymology: The specific epithet honors the Swiss botanist Dr. Henri François Pittier (1857-1950), who collected the type specimen. Pittier was one of the most influential scientists for Costa Rican botany; his work, leadership, and academic knowledge contributed to the formation of what is now the National Herbarium of Costa Rica and the development of scientific research on the flora of the country.
Distribution and habitat: From Costa Rica and Panama, where it grows at 1500−3036 m of elevation. Malaxis pittieri is recorded here for the first time for the flora of Costa Rica on the Pacific slope of the Central Volcanic and Talamanca mountain ranges (García Castro et al. 1993 included it in an unvouchered list of orchid species common to Costa Rica and Panama, but their report must be considered speculative). In Panama, M. pittieri it is known from Chiriqui province (Fig. 2). It inhabits in lower montane and montane pluvial forests, in understory clearings, on very moist, mossy, and well-drained soils. It usually grows as a terrestrial, but sometimes as an epiphyte up to 1.5 m in height on moss-covered tree trunks (Fig. 5).

A. From Costa Rica (Chinchilla et al. 2341, JBL-spirit).
B. From Panama (Bogarín et al. 11179, UCH).
Photographs by Isler F. Chinchilla (A) and Diego Bogarín (B).
Figure 6 Inflorescences of Malaxis pittieri bearing recently opened flowers, showing the lip with the entire apex.

A-C. From Costa Rica
A recently opened flower (Chinchilla et al. 2341, JBLspirit).
B-C. A flower in an early stage of dehydration showing the partially contracted lip (Chinchilla 3340, JBL-spirit).
D. From Panama: A dehydrated flower with a contracted lip that simulates being apically trilobed (Woodson & Schery 474, MO).
Photographs by Isler. F. Chinchilla.
Figure 7 Flowers of Malaxis pittieri.
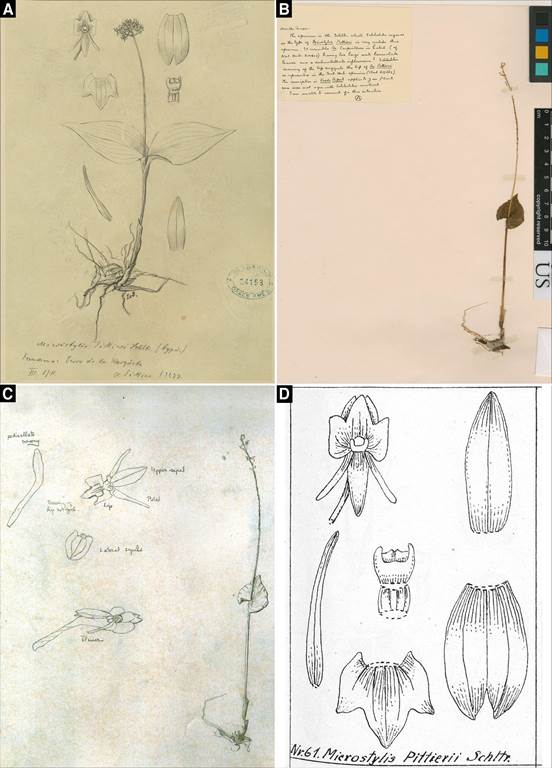
A. Tracing of the original drawings of the floral analysis of the holotype of M. pittieri (AMES-24153 / 00101736), excluding the habit of a plant of a different species. Courtesy of the Orchid Herbarium of Oakes Ames, Harvard University Herbaria.
B. Lectotype of M. pittieri (US-677667 / 00093447). Courtesy of the United States National Herbarium (US).
C. Drawings of the lectotype of M. pittieri (AMES-24152 / 00101737). Courtesy of the Orchid Herbarium of Oakes Ames, Harvard University Herbaria.
D. Schechter's drawings of the floral analysis from the holotype of M. pittieri, reproduced in Mansfeld (1931: t. 17, no. 61).
Figure 8 Type material of the Microstylis pittieri.
Phenology: Plants flower from March, June to September and bear fruits from June to August and December in the field.
Other specimens studied: COSTA RICA. Heredia: Parque Nacional Braulio Carrillo, Volcán Barva Station, path and forest between park office and crater, 10°07'20''N, 84°06'00''W, 2700−2900 m, 13 Jul 1991, H. Gay et al. 1507 (CR); Barva, San José de la Montaña, Área de Conservación Central, Parque Nacional Braulio Carrillo, sector Volcán Barva, cerca del sendero que conduce a la laguna del Volcán Barva, aprox. 230 m de la Estación del MINAE, 10°08'05''N, 84°07'10''W, 2711 m, 15 Dic 2018, I. Chinchilla et al. 4031 (JBL-spirit); Braulio Carrillo Nat. Park, canton of Barva, Barva Station, in pastures around station, near stream, 10°07'20''N, 84°06'00''W, 2580 m, 10 Sep 1990, S. Ingram et al. 507 (CR). Puntarenas: Buenos Aires, Cuenca Térraba-Sierpe. Buenos Aires, Potrero Grande, Tres Colinas, ca. 800 m, bajando hacia Helechales, 09°06'16,80''N, 83°03'53,60''W, 1600 m, 14 Jun 2006, D. Santamaría et al. 4504 (CR); Buenos Aires, P. N. La Amistad. Cuenca Térraba-Sierpe, Sabanas Esperanza, entre 1º y 3º Sabana, 09°04'33''N, 83°01'55''W, 1600 a 1800 m, 01 Jun 2006, J. F. Morales 13929 (CR). San José: Pérez Zeledón, Albergue Cuericí, mirador trail above the Albergue Cuericí, 09°33'30''N, 83°39'42''W, 2600 m, 23 Jun 2002, B. Boyle et al. 6331 (USJ); (Valle del El General, San Isidro) Vale Gen. S. Isidro, Jun, A. R. Endrés s.n. (W-1889−0030705); Pérez Zeledón, Rivas, Parque Nacional Chirripó, aprox. 200 m después del km 10, a la orilla del sendero, 09°26'35,30''N, 83°31'48,20''W, 3021 m, 23 Jul 2015, I. Chinchilla et al. 2305 (USJ); same data, I. Chinchilla et al. 2341 (JBL-spirit); same data, I. Chinchilla et al. 2342 (JBL-spirit); same locality, aprox. 300 m después del kilómetro 10, a la orilla del sendero, 09°26'33,67''N, 83°31'47,15''W, 3036 m, 13 Ago 2017, I. Chinchilla 3338 (JBL-spirit); same data, I. Chinchilla 3340 (JBL-spirit); same data, I. Chinchilla 3341 (JBL-spirit); (Pérez Zeledón) Fila Ventisqueros, Chirripó Gde., en el humus de la montaña alta (09°27'52''N, 83°31'21''W), 2000- 3000 m, Ago 1971, L. D. Gómez-Pignataro & W. C. Burger 3418 (CR). PANAMA. Chiriquí: Boquerón, Cordillera, Paso Ancho, Aguacate, Finca Aguacatal, camino a la naciente del Río Breque, 08°47'28,6''N, 82°34'39,7''W, 2126 m, 04 Ago 2014, D. Bogarín et al. 11179 (UCH); Boquete, (08°48'18''N, 82°26'16''W), 5000 ft (1524 m), Jun. 27, 1938, M. Davidson 801 (AMES, F, US); Potrero Muleto to summit, Volcán de Chiriquí, (08°48'40''N, 82°31'35''W), 13−15 Jul 1940, R. E. Woodson & R. Schery 474 (MO); Vicinity of Casita Alta, Volcán de Chiriqué (08°49'01''N, 82°29'35''W), ca. 1500−2000 m, 28 Jun-02 Jul 1938, R. E. Woodson et al. 830 (AMES, BM, US).
Malaxis pittieri is distinguished by having a single leaf per sympodial unit with the margins of the sheaths connate, forming a pseudostem and the blade cordateamplexicaul at the base, the inflorescence racemose, lax, bearing small, white flowers, the lateral sepals connate for more than half their length forming a galea behind the lip, and the lip trilobed, sagittate, with a green stripe along the costa that extends to the apex of the lip. Historically, it has been confused with Malaxis majanthemifolia Schltdl. & Cham., a species native from Mexico to Costa Rica (Fig. 9). However, M. pittieri differs in having white pedicels with a 360-degree twist (vs. greenish, not twisted), the sepals, petals, and lip white (vs. green to yellowish-green), the lip with a vertical green stripe (vs. dark brownish-green on the distal half), the disc cavity divided (vs. non-divided) by a costa into two subcavities, ending in a raised (vs. non-raised), rounded, concave (vs. flat) apical margin. Malaxis pittieri is vegetatively similar to M. acianthoides (Schltr.) Ames from Mexico and Guatemala, but the latter has green to yellowish-green flowers, the petals unguiculate, obliquely and transversely rhombic (vs. simple, narrowly lanceolate), the lip with lateral lobes of similar length to the mid lobe (vs. up to half the length of the midlobe), with ciliate-papillose (vs. entire) margins, and the disc cavity non-divided (vs. divided by a costa). Malaxis cobanensis Archila, Szlach. & Chiron, a Guatemalan endemic, is also vegetatively similar. But the latter has green sepals, petals and lip, the lip with disc cavity occupying up to ⅓ (vs. more than –) of its length, non-divided (vs. divided by a costa), and basally delimited by a transverse thickening (vs. not with a transverse thickening).

Photographs by Juan. F. Morales.
Figure 9 Plant habit (A) and inflorescence (B) of Malaxis majanthemifolia in situ (J. F. Morales 21193, CR).
The first collection of M. pittieri in Costa Rica was made by the French botanist and orchidologist Auguste R. Endrés between 1866 and 1874 (Endrés s.n., W-1889−0030705!; see Ossenbach et al. 2010). Subsequently, it has been collected multiple times in the country, but specimens have been misidentified as M. majanthemifolia and its presence has gone unnoticed for over 147 years. For example, the specimen cited as a Costa Rican voucher of M. majanthemifolia by both Pupulin (2002) and Dressler (2003) (Ingram et al. 507, CR) is a plant of M. pittieri. Similarly, the specimens cited as a Panamanian vouchers of M. majanthemifolia (see Bogarín et al. 2014, Dressler 2009, Williams 1946) are indeed M. pittieri. Malaxis majanthemifolia to date has not been recorded in Panama.
Nomenclatural notes: Schlechter (1913) described Malaxis pittieri (as Microstylis pittieri Schltr.) with the lip trilobed at the apex as shown in the drawings of the type (Fig. 8 C, D). The tracing of Schlechter's drawings of the holotype of M. pittieri at AMES (24153/00101736; Fig. 8A) show a floral analysis that coincides with the protologue of M. pittieri (and also with the original drawings of Schlechter, published posthumously by Mansfeld, 1931: t. 17, no. 61 ; Fig. 8D), but includes a plant habit drawing from a different species of Malaxis, because it has two leaves in the same sympodial unit, with opposite blades rounded at the base, and a corymbose inflorescence; inflorescence; Fig. 8A, center). This indicates a confusion with part of the type material when tracing the flower analysis and the plant habit of the holotype of M. pittieri in the Berlin herbarium (many of the flower analyses and plant habit of the types of orchid species that Schlechter described were traced after their publication upon Ames's request, for his own herbarium; Ames 1944). Oakes Ames, in a herbarium annotation on the specimen here designated as lectotype (US-677667/00093447, visible in the upper left corner in Fig. 8B) suggested that this drawing by the morphology of the leaves and inflorescence resembles Microstylis carpinterae Schltr. (Malaxis carpinterae (Schltr) Ames). But it is impossible to determine with certainty from this drawing alone. However, M. pittieri consistently has one leaf cordateamplexicaul at the base, and the lip with the entire apex (Fig. 6A-B); what happens is that when dehydrated, the lip apex undergoes a strong contraction that cleaves the apical margin, simulating a trilobed apex (Fig. 7A−D). The artifact described above was also verified with fresh material collected near the type locality of M. pittieri (D. Bogarín et al. 11179, UCH; Fig. 6B).
The isotype of the Microstylis pittieri designated here as the lectotype was previously indicated by Dressler (2009) as the holotype, and he also wrote it in an annotation on the specimen sheet. However, the holotype specimen that Schlechter used to describe M. pittieri was destroyed in 1943 in the bombing of the Berlin-Dahlem Botanical Museum (Ames 1944). In his annotation on the lectotype specimen, Ames also indicated that there was a duplicate in Berlin, which was undoubtedly the holotype.
Conservation status: Malaxis pittieri is a native of Costa Rica and western Panama. It is known from 11 locations (six in Costa Rica, five in Panama), in the Central Volcanic (Costa Rica) and Talamanca mountain ranges (Costa Rica and Panama). Its extent of occurrence (EOO) was estimated at 3562.297 km2 with an area of occupancy (AOO) of 48 km2. This species is known from five protected areas: Braulio Carrillo National Park and Chirripó National Park in Costa Rica; Volcán Barú National Park in Panama; and Talamanca Range-La Amistad Reserves / La Amistad National Park in both Costa Rica and Panama. In Costa Rica, five locations are found in protected areas, and one location is also in a forest near a protected area; similarly, in Panama, four locations are in protected areas, and one location near a protected area. Therefore, the species is listed on a global level and national level as Least Concern (LC).

