











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkBroughtonia R.Br., is a genus of six species, endemic to the Caribbean region. Three of species are present in Cuba: Broughtonia cubensis (Lindl.) Cogn.,B. ortgiesiana (Rchb.f.) Dressler, and B. lindenii (Lindl.) Dressler. While B. cubensis and B. ortgiesiana are both Cuban endemics, B. lindenii is also present in the Bahamas. Like Cuba, Jamaica also harbors two endemic Broughtonia species, B. negrilensis Fowlie, and B. sanguinea (Sw.) R.Br. On Hispaniola and Mona island (Puerto Rico) to the east, another species is found, B. domingensis (Lindl.) Rolfe (Díaz 2014, in Ackerman and collaborators). In Cuba, the distribution of B. lindenii and B. ortgiesiana is widespread. B. cubensis has been reported for several localities of the provinces Mayabeque (Loma La Coca), Artemisa (north coast of the municipality Mariel) and Pinar del Río (Los Pretiles and Guanahacabibes Peninsula). In Loma La Coca, the species is now extirpated due to habitat destruction and depredation. Reports for the North coast of Mariel and Los Pretiles have yet to be substantiated by the authors. In the Guanahacabibes Peninsula, B. cubensis is found in Cabo Corrientes and Cabo San Antonio in association with B. lindeniiand B. ortgiesiana where they are protected by law within the framework of Cuba's National System of Protected Areas. Although it is very difficult to differentiate between the plants on the basis of vegetative characters, their floral morphology and timing of anthesis clearly differ (Díaz Dumas, 1997a, 1997b).
Until now, it was assumed that B. cubensis and B. lindenii are not sympatric. However, field work carried out at Barra La Sorda (Cabo San Antonio) in 2005 revealed that both species were indeed sympatric, affixed to branches of Plumeria tuberculata (Apocynaceae) trees. Although their flowering seasons normally do not overlap, occasional natural crosspollination between B. cubensis and B. lindenii cannot be ruled out.
In 2004, at the site where we have studied spatial and temporal dynamics of B. cubensis (Figure 1), one individual of B. ortgiesiana was observed in flower (Figure 2) representing the first such report for this species in Cabo San Antonio. In 2007 the authors observed three fruits in this plant and later (2010) two more fruits were documented.


Collectively, these observations confirm that the species are indeed sympatric and possibly capable of eventual natural cross-pollination given their overlap in anthesis and geographic location. Evidence for sympatry is not limited to our 2004 observation. In January 2015, both species flowered concurrently again, with a second B. ortgiesianayielding six flowers. Further exploration of this site revealed five individual orchids with intermediate floral characteristics of both species (Figure 3-Figure 4).

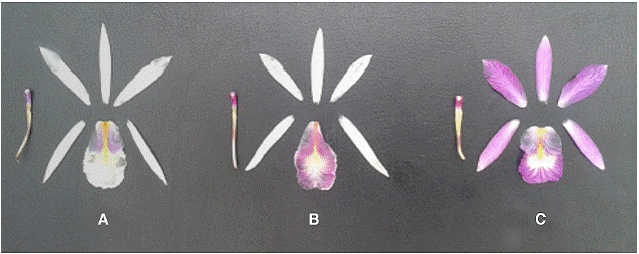
Figure 4:Dissected perianths of A -Broughtoniacubensis; B -B. ×guanahacabibensis; C -B. ortgiesiana.
Closer examination of B. cubensis individuals often reveals subtle floral variation (morphs), but never in the same proportion as those seen in other genera, namely Encyclia Hook. andTolumnia Raf.whose morphs are often problematic for taxonomists. Based on our direct field experiences, only rarely do morphs of B. cubensis re-occur in the same location or in different sites.
To our knowledge, all Broughtonia species are incapable of self-pollination. This fact, coupled with capsule/seed set in B. ortgiesiana, argues in favor of natural hybridization with B. cubensis- the only other Broughtonia species in flower at that location at that time. At two other sites where B. cubensis is present, B. ortgiesiana was not detected, nor were individuals possessing flowers intermediate in form. Taken together, we conclude that the intermediate forms in the Guanahacabibes Peninsula are the result of natural hybridization.
Broughtonia × guanahacabibensis Mújica, E.GonzálezetJ.M.Díaz, nothosp. nov.
Broughtoniacubensis (Lindl.) Cogn. × Broughtonia ortgiesiana (Rchb.f.) Dressler.
TYPE: Cuba. Pinar del Río: Sandino, Cabo San Antonio, near Roncali Lighthouse, originally collected on 23 January 2015. Type: E. Mujica 241 (holotype:Herbarium of the Soroa Orchid Garden, not registered).
Plants epiphytic, to 30 cm tall. Roots white, numerous, 2-3 mm in diameter. Pseudobulbsclustered, cylindric, fusiform, to 3 cm long, 1 cm wide, enclosed by scarious sheaths when young, with 2-3 internodes. Leaves 1-2, fleshy, rigid, erose, narrowly oblong, rounded to apiculate, to 8 cm long, 1 cm wide. Inflorescences a few-flowered racemes to 30 cm long; peduncle bracts sheathing. Floral bracts ovate, acute. Pedicellate ovary slender, to 2.5 cm long Flowers showy, with white sepals, the petals white with a purple central vein, the lip white, yellow at the center, with purple margin. Sepals lanceolate, acute, the dorsal one to 2 cm long, 4 mm wide, the lateral ones 2.3 cm long, 3 mm wide. Petals obovate-elliptic, acute, 1.8 cm long, 5 mm wide; labellum oblong, to 2 cm long, 1.2 cm wide, slightly emarginate, with undulate margins, enclosing the column at the base, the disc provided with branched yellow lines. Column white to pink, arcuate, to 1 cm long. Pollinia 8, yellow. Fruit an ellipsoid capsule, beaked, 12-13 mm long, on slender pedicel 8-10 mm long.
Paratype: Cuba. Pinar del Río: Sandino, Cabo San Antonio, near Roncali Lighthouse, January 2015, E. Mujica et E. González 179 (Herbarium of the Soroa Orchid Garden [not registered], flowers in spirit).
Etymology: From the Guanahacabibes peninsula, where the nothospecies was found.
DistributionandEcology: The habitat of Broughtonia × guanahacabibensis consists of xeromorphic subcoastal vegetation, colonizing Plumeria tuberculata - the dominant tree species in the area. It is also sympatric with other orchid species: Encyclia bocourtii Mújica & Pupulin, E. plicata (Lindl.) Britton & Millspaugh, Polystachya concreta (Jacq.) Garay & Sweet and Tolumnia guibertiana (A.Rich.) Braem.
The plants of B. cubensis and B. ortgiesiana are not easy to differentiate morphologically on the basis of their vegetative habit. However, their flowers are clearly different, the latters being very distinctive for their fringed white color labellum with purple color margins. The petals of B. ortgiesiana are very similar to those of B. cubensis but they differ in having a central nerve purple (fig. 4). This hybrid appears to comprise a viable population, thus introgression may be occurring in this site, resulting in a more variable population.

