Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción:
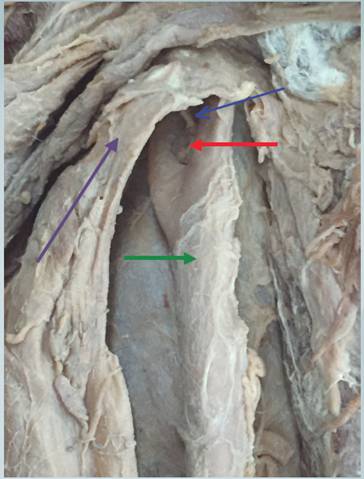
Se utiliza un cadáver preservado en DRP fórmula embalsamiento de un adulto mayor de sexo masculino. Se diseca el cuello con la técnica descrita en el Manual de Disección para identificar las estructuras vásculo-nerviosas del triángulo carotídeo y sus relaciones. Se procede a identificar la arteria carótida común hasta el seno carotídeo y su bifurcación en arteria carótida externa e interna derecha y a marcarla, se procede a realizar una disección minusiosa del cuerpo carotídeo y la rama del nervio glosofaríngeo como se muestra en la figura 3 de la region carotídea.
Discusión:
Generalidades
En conjunto, el cuerpo carotideo y el seno carotideo son considerados el principal baroquimioreceptor periférico.
El seno carotideo consiste en una dilatación ubicada en la bifurcación aórtica que posee mecanoreceptores en el segmento más externo de su túnica media y en la túnica adventicia que recolectan información relacionada con presión arterial media, cambio en la presión arterial, presión de pulso y frecuencia cardiaca (1). La información evocada por cualquiera de estos factores es sensada y transmitida via terminaciones aferentes hacia los centros de integración. (2)
La historia del seno carotideo se remonta al siglo XIX y la primera mitad del siglo XX cuando diversos investigadores documentaron variaciones ocurridas en el comportamiento, coloración de la piel, visualización de lechos vasculares o cambios de frecuencia cardiaca en animales tras la sección o estimulación de diversas estructuras del cuello. Herings y Koch documentaran los cambios de presión arterial secundarios a la estimulación mecánica del seno carotideo enseres humanos, ya antes comentada por Czermak en 1865 (3). Posterior a lo que se condujeron una serie de estudios electrofisiológicos en perros. Así comenzó una línea de investigación dedicada al entendimiento y descripción de las bases fisiológicas del baroreflejo (4, 1). Simultaneamente, Fernando de Castro, un discípulo del acreedor del Premio Novel Santiago Ramon y Cajal demostró que la estructura anatómica conocida como cuerpo carotideo consistía en un tejido altamente vascularizado situado en la bifurcación de la carótica y describió su innervación e irrigación. (5, 6)
El investigador Corneille Heymans y sus colaboradores estaban muy interesados en las hipótesis Castro y trabajaron en forma conjunta para probar sus hipótesis. Tres años después de la publicación de la investigación de Hering sobre el seno carotideo, en 1930, Heymans y colaboradores describen la función quimiorreceptora del cuerpo carotideo.
Ellos realizaron observaciones de las modificaciones vasomotoras y electrofisiológicas inducidas por cambios en la concentración arterial de Oxígeno (PaO2) y de dióxido de carbono (acidemia), y la transmisión de las señales vía ganglio petroso y nervio glosofaríngeo hacia los centros respiratorios y vasomotores del tronco encefálico. Sus estudios le valieron a Heymans el Premio Nobel de fisiología y Medicina en 1938.
Desde esa época inicia una carrera por descubrir todas las posibles implicaciones médicas y quirúrgicas de estas dos estructuras. En la actualidad se conducen investigaciones que ponen al cuerpo carotideo como blanco para el tratamiento de enfermedades variadas, desde desordenes del equilibrio ácido base en pacientes en estado crítico, hasta el manejo de enfermedades crónicas como la diabetes y la hipertensión arterial. Algunos autores han documentado el fallo de esta estructura tras la resección bilateral de paragangliomas, lesiones bulbares dorsales, siringobulia, tras radioterapia, endarterectomia bilateral carotidea y traumatismos cervicales; con la consecuente aparición de hipertensión, taquicardia, flushing, labilidad emocional y cefalea en los pacientes afectados. (7)
Cabe destacar que aunque tradicionalmente se reconoce que las terminaciones nerviosas del seno carotideo son principalmente sensibles a cambios de presión y las del cuerpo carotideo reaccionan principalmente a la composición química del plasma, esa distinción no es absoluta, y se ha visto que las células del cuerpo carotideo también reaccionan ante estímulos mecánicos desencadenando respuestas que son sinérgicas pero independientes a las del seno carotideo (8). De modo que en la actualidad, el cuerpo carotideo se estudia ya no solo desde su importancia como el principal quimiorreceptor periférico, capaz de sensar los cambios de PaO2, PaCO2 y pH, sino ahora también como un baroreceptor importante por sus posibles implicaciones directas en la regulación de la presión arterial. (9).
Anatomía Macroscópica:
Se trata de una estructura derivada del tercer arco faríngeo, redondeada y en algunos casos bilobulada, situada en la adventicia de la región posteromedial de la bifurcación aórtica (10). Sus dimensiones no son claras, pues al encontrarse rodeado por gran cantidad de tejido adiposo y tejido conectivo, sus medidas varían en gran manera de acuerdo a la preparación del tejido por parte del investigador. En forma aproximada podríamos decir que su dimensiones oscilan entre los siguientes rangos: 3,3-7mm de alto, por 3-4mm de profundidad y 1.7- 2mm de ancho (9, 10, 11). Sin embargo, un estudio reciente conducido por Ortega-Saenz que se dedicó al análisis morfológico y funcional de cuerpos carotideos resecados en cadáveres humanos mostró que existen diferencias muy importantes en el volumen de los mismos entre una personas y otra; incluso habiendo diferencias entre los dos cuerpos carotideos del mismo paciente. Las mediciones realizadas en esa investigación arrojaron un volumen promedio en adultos jóvenes (definido como edad menor a 50 años) de 19,3±4.7mm3 y de 22.9±6.6mm3 en adultos mayores (definido como edad mayor a 50 años). Además el estudio vio una tendencia a un mayor volumen en los sujetos masculinos, aunque esta no alcanzó la significancia estadística (11).
La irrigación del cuerpo carotideo proviene también del tercer arco faríngeo. En condiciones normales, la arteria carótida común origina las arterias glómicas que penetran la capsulas de tejido conectivo del cuerpo carotideo para irrigarlo (10). En ocasiones una rama de la arteria faríngea ascendente (originada de la cara posterior de la arteria carótida externa), provee una irrigación accesoria al cuerpo carotideo. Esto último es de particular importancia en tumores del cuerpo carotideo donde no ligar adecuadamente estas estructuras vasculares puede traer complicaciones quirúrgicas (12)
Tanto el seno carotideo como el cuerpo carotideo reciben su innervación aferente de fibras del ganglio petroso a través del nervio del seno carotideo. Estas fibras se unen al nervio glosofaríngeo y las señales transmitidas son integradas en el tallo cerebral dorsomedial, específicamente en los centros respiratorio y vasomotor. (13, 14, 15)
Anatomía Microscópica
Es un tejido altamente vascularizado, organizado en unos 12 acúmulos (glomérulos o lobulillos) de células Glómicas Parenquimatosas Tipo I y células de sostén o Tipo II. Las células de sostén poseen un fenotipo funcional glial, semejante a los astrocitos del sistema nervioso central y son importantes no solo en términos estructurales, sino que participan en la regulación metabólica del tejido. El parénquima del cuerpo carotideo deriva de las crestas neurales por lo que no es de extrañarse que las células Glómicas tipo 1 posean fenotipo funcional que se asemeja al de las neuronas.
Se trata de grandes células esféricas acopladas química y eléctricamente a través de sinápsis químicas y canales de hendidura con las otras células del tejido y los axones a los que transmiten las señales desde este órgano. Entre estos lobulillos se ubican tabiques de espesor variable de tejido conectivo que contienen abundantes terminaciones nerviosas y capilares. El acople sináptico entre las células Glómicas Tipo I y las terminaciones neuronales aferentes ocurren gracias a la presencia de numerosas vesículas con neurotransmisores que contienen dopamina, acetilcolina, serotonina y adensintrifostato (ATP) entre otros péptidos dentro de la célula. (11) (16) (17) (18)
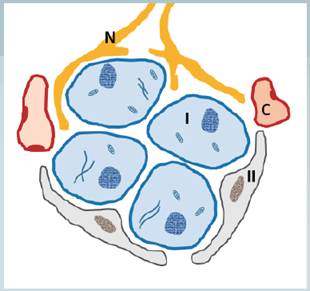
Fig 1 Dibujo esquemático de un Lobulillo del Cuerpo Carotideo. Se observan las Células Glómicas tipo I (I) rodeadas por Células Glómicas tipo II (II). También se representa la presencia de capilares (C) rodeando al lobulillo y las terminaciones nerviosas aferentes (N).
Está documentado que algunas de las células Glómicas tipo I, a pesar de estar diferenciadas y ser funcionales dentro del tejido, conservan su potencial mitótico, con capacidad de reproducirse para mantener la población de células tipo I.
Además, en una de sus investigaciones, Pardal J y colaboradores lograron documentar que dentro de los tabiques de tejido conectivo y entre las islas de células tipo II existen células madres quiescentes, que en condiciones de hipoxia son capaces de diferenciarse y proliferar en cualquiera de las células del cuerpo carotideo. (19)
Aspectos Fisiologicos
Como se mencionó, el cuerpo carotideo esta irrigado por una densa red de capilares que llevan sangre a todo el tejido. Las células glómicas tipo I poseen la capacidad de interactuar con el plasma y desencadenar una respuesta a la hipoxia y cambios en el pH liberando neurotransmisores que causan una despolarización de las terminaciones nerviosas de los axones asociados. (20). Numerosos estudios han buscado esclarecer el mecanismo a través del cual las células Glómicas tipo I censan y transmiten la señal. Hasta el momento el mecanismo no es completamente claro. El modelo aceptado de muestra en la figura 1 y postula que el cierre de canales de potasio tipo TASK y BK desencadenan despolarización de la membrana celular, esta a su vez favorece la entrada transmembranal de calcio a través de canales voltaje-dependientes. Esta elevación del calcio intracelular produce la movilización de las vesículas intracelulares que contienen los neurotransmisores hacia la membrana celular para su liberación, con la subsecuente generación de un potencial de acción en los axones de las neuronas aferentes (21). Además se ha documentado la presencia de uniones comunicantes y por lo tanto un acoplamiento eléctrico entre estas células y las terminaciones nerviosas por lo que la simple generación de un potencial de acción en la célula glómica podría transmitirse a la fibra axonal sin la necesidad de la transducción mediada por un neutrotransmisor (18, 22, 23, 24, 25, 26).
Los somas de estos axones se ubican el núcleo del tracto solitario y tienen conexiones con la sustancia reticular, predominantemente en el centro respiratorio y generan una respuesta hiperventilatoria bifásica. Esta respuesta bifásica se caracteriza por un aumento de la ventilación de entre 20 y 30 minutos, seguido por una fase de declinación variable (21).
Dentro de los avances más recientes en el campo se ha documentado una sensibilidad a la concentración de glucosa extracelular en este tejido, y se postula que tiene un rol importante su regulación (27, 28, 29, 30). Incluso se ha postulado que hiperosmolaridad plasmática, hipercalcemia e hipertermia también podrían tener un rol en la activación del cuerpo carotideo (31).
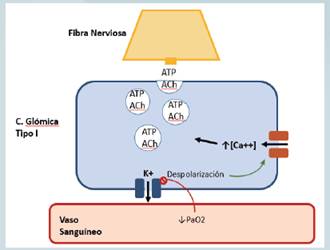
Fig 2. Dibujo esquemático del modelo fisiológico para la sensibilidad al oxígeno en las células Glómicas tipo I. Un descenso en la concentración sanguínea de oxígeno produce un cierre de canales de Potasio. Esto desencadena una despolarización de la membrana que a su vez ocasiona una apertura de canales de calcio voltaje-dependientes. La elevación intracelular del calcio favorece la movilización de las vesículas con neurotransmisor a la membrana celular para su liberación. ATP: Adenosintrifosfato; ACh: acetilcolina, K+: potasio; Csa++: Calcio.