











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La variabilidad de colores del pelaje en los gatos domésticos (Felis catus) obedece a un proceso de desarrollo y funcionamiento de las estructuras implicadas en la formación o génesis del pigmento. Este proceso está regulado por una serie de genes y normalmente conduce al desarrollo del denominado de tipo salvaje (atigrado). Cualquier otra variación del color en el pelaje es causada por mutaciones y obedece a una serie de genes que codifican para coloraciones que van desde el blanco hasta el naranja. Las frecuencias de alelos mutantes y silvestres de los genes que codifican la coloración del pelaje son específicas de cada población y reflejan sus estructuras genéticas (Kholin, 2012). Los colores y patrones del pelaje son muy variables en los gatos y son determinados, principalmente, por varios genes que se expresan siguiendo las características de la herencia mendeliana (Gershony, 2014).
La forma en que se han distribuido las poblaciones de gatos domésticos alrededor del mundo a lo largo de la historia está altamente relacionada con movilizaciones antrópicas, lo que se denomina la hipótesis de migración histórica (Ruiz-García et al., 2005). Se han analizado varias poblaciones en distintas partes del mundo desde la perspectiva de la genética de poblaciones biogeográficas, utilizando las características que codifican para la longitud, la distribución, la coloración del pelaje y anomalías esqueléticas. Aunque la mayoría de los estudios de las poblaciones de gatos se han realizado en Europa, en América Latina, específicamente en Colombia, se han reportado varias investigaciones sobre la genética de poblaciones de esta especie.
Ruiz-García y Álvarez (2003) estudiaron seis poblaciones latinoamericanas de gatos, incluyendo la ciudad de Bogotá, utilizando marcadores que codifican características morfológicas del pelaje y marcadores moleculares nucleares microsatélites, concluyendo que la evolución global de los marcadores utilizados obedecería a presiones naturales y no a eventos selectivos. Por otro lado, Pardo et al. (2014), en un estudio en Montería, Colombia, de la estructura genética en una población de gatos domésticos mediante marcadores del pelaje, encontraron bajos niveles de variabilidad genética tanto a nivel global como poblacional; pese a esto, los marcadores Orange y Manchado de blanco evidenciaron ausencia de equilibrio. Peña-Cruz et al. (2015), en un estudio genético-poblacional de gatos domésticos realizado en la ciudad de Cali, hacen el primer reporte para Colombia de los alelos inhibitor y ticked abyssinian. Pardo et al. (2015) evaluaron la diversidad genética en poblaciones de gatos domésticos de Lorica, donde el marcador Orange evidencia altas frecuencias alélicas y una selección natural y artificial debido a presiones antrópicas. Montes-Díaz et al. (2015) reportaron selección natural y artificial de los marcadores non-Agouti y Tabby abissynian en la población de Cartagena.
La variación fenotípica en la hibridación de especies o subespecies es una condición para que los investigadores de vida silvestre identifiquen poblaciones parentales y sus híbridos en el campo; por tal razón, los marcadores fenotípicos, especialmente los relacionados con la coloración del pelaje (Devillard, 2014), constituyen una valiosa herramienta cuando se analiza la estructura genética de las poblaciones en medios naturales, debido a su gran contenido informativo, fácil manipulación y rápida obtención de resultados.
Las frecuencias de estos marcadores genéticos comúnmente usados para dichos estudios poblacionales podrían verse afectadas por variables ambientales, selección natural, selección artificial y/o los efectos migratorios que caracterizan la historia del establecimiento de las poblaciones humanas (Ruiz-García & Álvarez, 2008). Por esta razón, la presente investigación tuvo como finalidad conocer el grado de diversidad y estructura genética en poblaciones de Felis catus mediante marcadores que codifican el pelaje en Sahagún, Córdoba, Colombia.
Materiales y Métodos
Sitio de estudio
El estudio se realizó en la zona urbana de Sahagún (8° 56` 58`` latitud Norte y 75° 26` 52`` longitud Oeste, con una temperatura promedio de 28 °C). Se muestrearon los barrios Bosque Barají, Santa Lucía, Alfonso López, Venecia, Camilo Torres y Belalcázar.
Obtención de datos
Se realizaron muestreos aleatorios entre los meses de Mayo y Agosto del año 2015. Mediante excursiones urbanas y observación directa, se realizó una clasificación fenotípica de cada uno de los individuos adultos encontrados en Sahagún; no se tuvieron en cuenta gatos callejeros y cada ruta se utilizó solo una vez, a fin de evitar el remuestreo, atendiendo a la presencia o ausencia de los marcadores autosómicos Non-agouti(a); Blotched tabby (Tb); Dilution (d), Pelo largo (l); Manchado de blanco (S) y Dominante blanco (W) y el locus ligado al sexo O (Orange); por último, se tomaron registros fotográficos de cada individuo. De igual manera, se realizó una encuesta sobre sexo, edad raza, origen y preferencias antrópicas; se construyó una base de datos y los resultados se analizaron con varios estadísticos.
Diseño estadístico
La estimación de las frecuencias alélicas de cada marcador, el coeficiente de diferenciación genética (GST) y flujo génico (Nm); se estimaron a través del programa PopGene 1.31 (Yeh, 1999). El equilibrio Hardy-Weinberg fue probado mediante un chi-cuadrado; la distancia genética entre las poblaciones, el índice de diversidad genética correspondientes a la heterocigosidad esperada (He), fueron calculadas siguiendo los parámetros de Nei (1972); y la estructura genética de las poblaciones, fue calculada atendiendo a los índices de fijación propuestos por Wright: FIS, FIT y FST, se calcularon mediante el programa FSTAT v. 2.9.3.2 (Goudet, 2001). La elaboración del dendrograma que representa los valores estimados de distancia genética se realizó a través del método UPGMA (Unweighted Pair Group Methodwith Arithmetic Mean), empleando el programa MEGA 5.2 (Tamura, 2011).
Resultados y Discusión
Se muestreó un total de 153 individuos, de los cuales el 42,2% eran machos y el 57,8% hembras. Estos porcentajes de sexos encontrados, distribuidos entre las seis subpoblaciones muestreadas de la ciudad, podrían indicar un buen tamaño de muestra para la realización del estudio.
Frecuencias alélicas
El cuadro 1 contiene las frecuencias alélicas para cada subpoblación. Se puede observar que el marcador Manchado de blanco es el más frecuente, especialmente en las subpoblaciones de Belalcázar (q= 0.7500), Santa Lucía (q= 0.6667) y Venecia (q= 0.6500). Este hecho podría estar relacionado con diversos factores, tales como: el continuo y rápido crecimiento poblacional de gatos domésticos, lo cual incrementa considerablemente el flujo genético, aumentando así los niveles de panmixia; factores ambientales como las altas temperaturas podrían favorecer tanto la presencia como el aumento de individuos portadores de dicho marcador (Eizirik, 2010). Y, por último, las preferencias antrópicas por razones estéticas también podrían estar relacionadas con el aumento en la frecuencia alélica de este gen (Pardo et al., 2015), lo cual fue evidenciado por la favorabilidad detectada al realizar el estudio, revelando una posible selección artificial.
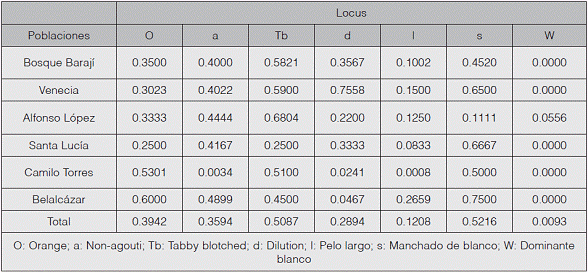
Un aspecto importante de destacar es que la frecuencia del gen Orange fue alta (p= 0.3942) en comparación con las reportadas por otros autores (Ruiz-García & Álvarez, 2003, 2008; Ruiz-García et al., 2005, Peña-Cruz et al., 2014). En ambientes urbanos, donde las densidades poblacionales son altas, los individuos que portan el marcador Orange podrían ser favorecidos, ya que tienden a ser altamente sociables con otras especies, principalmente la humana; esto hace suponer que los gatos domésticos portadores de este gen están mejor adaptados a las condiciones imperantes de una localidad con alta presión demográfica y ese podría ser un factor de selección sobre otros que no lo portan (Schmidt, 2009; Kaelin & Barsh, 2013). Por otra parte, el marcador Orange, además de estar ligado al cromosoma X, también es epistático sobre el Non agouti, (Schmidt, 2009); esta correlación negativa podría aumentar indirectamente las frecuencias de este marcador.
Por su parte, el marcador Dominante blanco, que es epistático y responsable del color blanco en la totalidad del cuerpo del animal, solo se registró en una de las subpoblaciones, Alfonso López, siendo este el marcador menos frecuente en la población de Sahagún. Esto podría ser atribuido a que los individuos de color blanco tienden a desarrollar complicaciones auditivas (David, 2014), lo que coloca en desventaja selectiva a aquellos que porten este marcador. Este hecho podría ocasionar la muerte a una edad temprana, disminuyendo notablemente el número de animales blancos y, por consiguiente, su frecuencia alélica.
La presencia de todos los marcadores estudiados en la población de gatos domésticos de Sahagún evidencia la gran variedad de genes disponibles en la zona. Esto podría explicarse por el hecho de que se trata de un sitio de interconexión con la troncal de oriente que viene desde el interior del país, y con la vía a la capital de Córdoba, favoreciendo el movimiento de migrantes y aumentando de forma considerable el flujo génico de personas e indirectamente de los gatos domésticos
Hardy-Weinberg
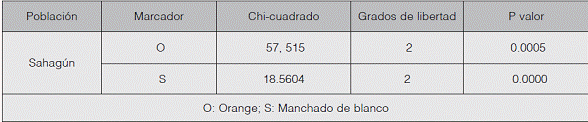
Con respecto al test de equilibrio de Hardy-Weinberg (cuadro 2), a nivel global, las poblaciones reportaron ausencia de equilibrio para los marcadores O (Orange) y S (Manchado de blanco). Una de las razones que explicarían la desviación de equilibrio es la relación directa entre las poblaciones de gatos y las poblaciones antrópicas, puesto que los seres humanos han jugado un papel significativo en el establecimiento y mantenimiento de la variación del color del pelaje en los animales domésticos, debido a que tienen preferencias por coloraciones exóticas y variantes blancas (Linderholm & Larson, 2013). Esto indicaría que la estructura actual de colores en poblaciones de gatos domésticos podría ser el resultado de continuas presiones selectivas que han reducido la frecuencia de alelos silvestres y han inducido el aumento de mutantes, lo que ha favorecido algunas características fenotípicas más que otras. Además, las alternativas del marcador Orange conocidas como carey y calicó son preferidas por los habitantes del lugar, debido a que su pelaje presenta tres colores (hembras heterocigotas); esto se constató durante el muestreo y podría explicar el exceso de heterocigotos evidenciados en los índices de fijación FIS, FIT y FST, (cuadro 3).
Otro aspecto que podría afectar esta desviación es la cercanía geográfica entre las subpoblaciones estudiadas, lo que causa un aumento considerable del flujo génico entre ellas, dado que la existencia de un constante intercambio de genes previene eventos de endogamia al interior de las poblaciones y demuestra que los apareamientos fueron aleatorios (Siepielski, 2013), provocando, por lo tanto, un aumento de genotipos heterocigotos en la población, lo que conduce a una desviación del equilibrio de Hardy-Weinberg.
Estructura poblacional
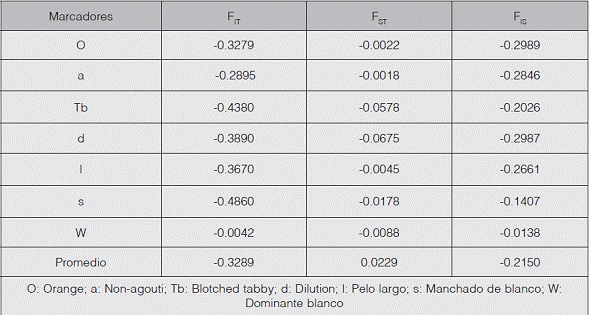
Los valores negativos de FIS en todos los marcadores que muestra el cuadro 3 evidencian un exceso de heterocigotos de los individuos con respecto a cada población y, por lo tanto, se asume ausencia de consanguinidad, con valores que oscilan entre -0.0138 para el marcador Dominante blanco y -0.2989 para el gen Orange. Con relación al estadístico FIT, en promedio se obtuvo un exceso de heterocigotos de los individuos con respecto a la población total, abarcando valores de -0.0042 a -0.4860 para los marcadores Dominante blanco y Manchado de blanco, respectivamente. Por otro lado, el valor promedio de FST resultó ser bajo, lo que indica poca diferenciación génica entre las poblaciones.
El exceso de heterocigotos obtenidos a través de los distintos índices de fijación (FIS y FIT) muestra que estos presentan una estructura casi homogénea. Esto podría estar relacionado con la cercanía geográfica de las poblaciones, lo que ocasionaría un aumento del flujo génico entre ellas. Los dueños de las mascotas no solo se desplazan por toda la ciudad, sino que también llegan de otros lugares llevando consigo animales domésticos; por esta razón, se podría intuir que el flujo génico de estas poblaciones de gatos es generalmente alto y era de esperarse que las frecuencias alélicas de los genes evaluados no hayan diferido sustancialmente entre las subpoblaciones urbanas en Sahagún, lo que fue confirmado por el estadístico FST, ya que todas las zonas analizadas se comportan como una sola población.
El valor de FST muestra que los barrios no se comportan como poblaciones independientes, lo cual puede demostrarse por el dominio antrópico en la generación de descendientes con paternidades simultáneas de gatos domésticos, aumentando así el flujo genético dentro de las poblaciones, disminuyendo la endogamia e incrementando la panmixia. Asimismo, no existen barreras que limiten el flujo de los gatos, por medio del movimiento de sus dueños de un lado al otro de la ciudad; debe aclararse que el valor del FST se debe al sistema de apareamiento y no a la subdivisión poblacional (Ruiz-García & Álvarez, 2008; Rosenfeld, 2010).
Diferenciación genética y flujo génico
El nivel de diferenciación genética en la población de gatos domésticos de Sahagún fue muy bajo (cuadro 4). Esto indica que aproximadamente el 6,7% de la variación detectada se debe a diferencias entre las subpoblaciones; por lo tanto, los seis barrios no fueron significativamente diferentes para los marcadores estudiados y esto sugiere que todas ellas se comportan como una sola población. El valor de flujo génico permite suponer que las poblaciones mantienen un grado de intercambio genético, asumiéndose un total de dos migrantes por generación; además, la cifra obtenida resultó ser mayor que 1, lo que indica que las subpoblaciones (barrios) se encuentran muy relacionadas genéticamente y se comportan como una sola población, la deriva génica es despreciable y el flujo génico hace que todas las poblaciones evolucionen de manera cohesiva como un conjunto, situación a la que se atribuye la aproximación de todas las poblaciones desde el punto de vista estructural (Pardo, 2015).
Distancia genética
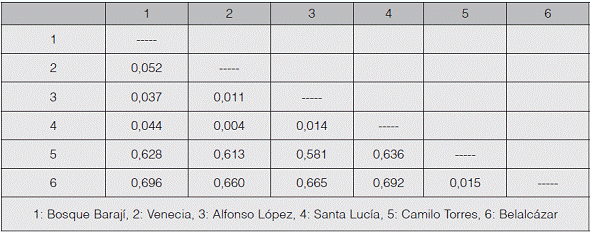
La distancia genética de Nei (1972) entre las poblaciones mostró a Venecia y Santa Lucía como las más cercanas (cuadro 5), mientras Bosque Barají y Belalcázar resultaron ser las de mayor diferencia génica. La población de Alfonso López presentó los mayores valores de distancia genética en comparación con el resto de las poblaciones. Los bajos valores de distancias podrían obedecer a la cercanía geográfica, lo que explicaría el hecho de que no hay una diferenciación entre las subpoblaciones, sugiriendo análisis generales y no de subestructura poblacional.

En el dendograma UPGMA elaborado a partir de los valores de distancia genética de Nei (1972) para las seis subpoblaciones de Sahagún (figura 1), se evidencia la similitud genética entre Venecia y Santa Lucía, a las cuales se asocia Alfonso López; asimismo, las poblaciones Camilo Torres y Belalcázar forman otra asociación. Esa agrupación estaría relacionada con la antigüedad de los barrios, pues en subpoblaciones cercanas las presiones evolutivas ejercen presiones similares y se esperaría que fueran más parecidas entre sí que con subpoblaciones más contemporáneas.
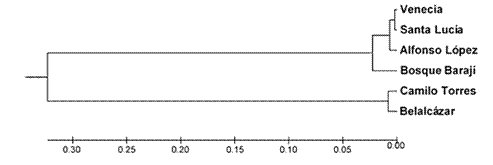
Figura 1: Dendograma construido con el método UPGMA basado en la distancia genética de Nei (1972) de poblaciones de Felis catus en Sahagún.
Conclusiones
El marcador con la mayor frecuencia fue Manchado de blanco, debido a que podría estar siendo favorecido por las elevadas temperaturas y las preferencias antrópicas por razones estéticas. Esto evidencia una posible selección artificial, no solo por este marcador, sino también por el marcador Orange, debido a que las hembras heterocigotas (tricolor) son preferidas frente a otros individuos que portan otros marcadores, de tal manera que sus frecuencias aumentaron progresivamente. La presencia de todos los genes estudiados en la población de gatos de Sahagún muestra la gran variedad de genes disponibles en la zona. La diversidad genética total encontrada en el lugar fue moderada. La mayor parte de la diversidad génica se encontró dentro de las poblaciones y poca entre las poblaciones, lo cual evidencia que las poblaciones locales comparten una gran proporción de la diversidad total, razón por la cual se comportan como una sola. Se encontró, además, ausencia del equilibrio de Hardy-Weinberg para los alelos O (Orange) y S (Manchado de blanco); esta desviación podría estar relacionada con una posible presión selectiva, antrópica y natural en estos marcadores.

