











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Durante los últimos cinco años, el Laboratorio de Ingeniería de Tejidos (LAINTEC) del Centro de Investigación en Biotecnología (CIB) del Instituto Tecnológico de Costa Rica (ITCR) se ha dedicado a la implementación, establecimiento y optimización de protocolos y estrategias para el cultivo in vitro de células de la piel con fines terapéuticos. Mediante técnicas de ingeniería de tejidos es posible reconstruir in vitro epitelios autólogos o alogénicos para tratar lesiones y afecciones en la piel.
Los queratinocitos son las principales células constituyentes de la epidermis, la capa más externa de la piel. El cultivo in vitro de estas células ha permitido la formación de epitelio estratificado y queratinizado para utilizarlo con fines terapéuticos en quemaduras y úlceras crónicas (Arango, Chamorro, Restrepo, Correa y Henao, 2009; Gil, 2011), campo en el que más de 30 años de experiencias clínicas a nivel internacional han demostrado resultados satisfactorios (Atiyeh y Costagliola, 2007).
Sin embargo, los queratinocitos presentan gran dificultad para crecer in vitro, ya que carecen de la producción de una serie de sustancias necesarias para su adhesión y proliferación (Arango et al., 2009 y Gil, 2011). Se ha demostrado que el desarrollo in vivo de las células epiteliales depende de complejas interacciones y diferentes estímulos procedentes de los fibroblastos (Arvelo, 2007), que son las principales células constituyentes de la dermis. Los fibroblastos participan activamente en la formación de la matriz extracelular, que es fundamental para el crecimiento de los queratinocitos (Arango et al., 2009). En suma, en ausencia del componente dérmico, los queratinocitos presentan deficiente organización, proliferación, expansión, diferenciación y síntesis de proteínas de la unión dermoepidérmica (membrana o lámina basal) (Arango et al., 2009 e Ikada, 2006).
Para solucionar esta situación, los queratinocitos aislados del tejido se inoculan sobre una capa de células alimentadoras (como, por ejemplo, de líneas celulares murinas tales como 3T3-J2 o 3T3-152), cuya mitosis ha sido inhibida por radiación u otros métodos. Sin embargo, estas células mantienen su actividad metabólica, produciendo los factores de crecimiento requeridos por los queratinocitos. Así, la función de esta capa alimentadora es actuar como una matriz mesenquimal in vitro (Atiyeh y Costagliola, 2007).
Las células vivas de la capa alimentadora (feeder layer) tienen la capacidad de promover la adhesión, crecimiento y diferenciación in vitro de las células epiteliales, secretan factores de crecimiento y proteínas de la matriz extracelular e inhiben el sobrecrecimiento de los fibroblastos dérmicos (Arvelo, Pérez y Cotte, 2004; Freshney, 2010; De Diego et al., 2006; Arvelo, 2007; Castro, 2009 y Arango et al., 2009).
La preparación de capas de células alimentadoras puede realizarse a partir de cultivos celulares subconfluentes mediante irradiación con rayos X o rayos γ a diferentes dosis. De manera alternativa, se pueden aplicar tratamientos químicos con mitomicina C (100μg/L) (Freshney, 2010). Ambos métodos generan células no proliferativas, mitóticamente inactivadas, que se mantienen viables hasta por tres semanas con su actividad metabólica inalterada (Castro, 2009 y Freshney, 2010). Aunque la irradiación causa un descenso en el número de células funcionales, éstas pueden seguir sintetizando moléculas específicas (Jessop y Hay, 1979).
Debido a que el ITCR no cuenta con instalaciones de irradiación gamma, se implementó un protocolo para la producción de capas alimentadoras mediante rayos X con un generador de rayos X (TOSHIBA, EX 250G) de la Escuela de Ciencia e Ingeniería de los Materiales de la institución.
Materiales y Métodos
Condiciones generales de cultivo
Como materia prima para la preparación de las capas alimentadoras, se utilizó la línea celular de fibroblastos murinos 3T3-Swiss albino (ATCC, CCL-92), a partir de pasaje 10. Se utilizó el medio de cultivo DMEM (4,5 g/L glucosa, GIBCO) suplementado con 7-10% de SFB (SIGMA), 2% glutamina (4mM, GIBCO), 1% penicilina-estreptomicina (10 000 u/ml penicilina G, 10 000 µg/ml de sulfato de estreptomicina; GIBCO) y 1% piruvato de sodio (0.11mg/ml, SIGMA), con un pH final de 7,5-7,7. Las células se mantuvieron en frascos de cultivo celular con un área de superficie de 25 cm2 y 75 cm2, en un ambiente al 5% de CO2, 37 ºC y alta humedad. Se realizaron cambios de medio de cultivo cada 2-4 días, dependiendo de la confluencia. Los subcultivos se realizaron al alcanzar 70-90% de confluencia, mediante el método enzimático con tripsina 0,05% y EDTA 0,02% (Freshney, 2010).
Establecimiento de la dosis de rayos X
Las células se cultivaron hasta alcanzar 70-90% de confluencia en frascos de cultivo de 25 cm2 con 4 ml de medio de cultivo. El día de la irradiación, se cambió el medio de cultivo con 4 ml de medio fresco y se colocó el frasco de cultivo dentro de una bolsa regular de plástico sellada herméticamente. Las células fueron irradiadas con diferentes dosis (30, 60, 70 y 80 Gy), ajustando el equipo a 250 kV, 5mA y a una distancia de 20 cm de la fuente, con las botellas de cultivo en posición horizontal. La dosis de irradiación fue determinada por el tiempo de exposición de las células a estas condiciones, correspondiendo a 12 min para 30 Gy, 24 min para 60 Gy, 28 min para 70 Gy y 32 min para 80 Gy. Luego de la irradiación, se suspendieron las células por el método de tripsina-EDTA y se inoculó 1 ml a una concentración de 50.000 células/ ml por pozo en placas de 24 pozos. Cada dos o tres días se resuspendieron las células de tres pozos al azar, donde se determinó la concentración celular por conteo al hematocitómetro y la viabilidad celular mediante MTT (3-[4,5-dimetiltiazol-2-il]-2,5-bromuro de difeniltetrazolio). En todos los casos (menos con la dosis de 30 Gy) se hicieron controles de células no irradiadas, mantenidas durante el mismo tiempo y bajo las mismas condiciones de cultivo. Se realizó una repetición de tres réplicas para las dosis de 60 y 70 Gy, y tres repeticiones de tres réplicas cada una para la dosis de 80 Gy.
Concentración celular
Se resuspendieron las células mediante el método de tripsina-EDTA y se tiñeron con azul tripán (0,4%, SIGMA) en una proporción 1:2. Se realizaron dos conteos por muestra de células vivas (incoloras, refringentes) y células muertas (azules) en una cámara de Neubauer (Freshney, 2010). Los porcentajes de viabilidad y mortalidad celular se obtuvieron comparando la concentración de células vivas y muertas con la concentración celular total.
Viabilidad celular por MTT
La viabilidad celular se calculó mediante el método descrito por Freshney (2010) con algunas modificaciones. En resumen: las células se incubaron durante 3 a 6 horas con MTT (0.5 mg/ ml, SIGMA) disuelto en el medio de cultivo, el cual se retiró para disolver las sales de formazán en etanol 95% filtrado (0.22 μm). El producto de la reacción fue cuantificado mediante espectrofotometría (OD 570 nm) y se reporta como el porcentaje de viabilidad en comparación con el control de células no irradiadas.
Eficiencia de plateo
Se utilizó el procedimiento descrito por Freshney (2001) y Franken, Rodermond, Stap, Haveman y Van Breel (2006). Se resuspendieron las células mediante el método de tripsina-EDTA y se inocularon en placas multipozo a concentraciones crecientes pero en baja densidad. Luego de aproximadamente dos semanas, se fijaron las células con metanol y se tiñeron con cristal violeta. Para establecer la mínima concentración necesaria para obtener colonias en la línea 3T3, se utilizaron inóculos de 50, 100, 150, 200, 250, 300, 350 y 400 células/cm2 en placas de 24 pozos con 600 µl de medio de cultivo, en tres repeticiones con células en el mismo pasaje. A partir de estos datos, se utilizaron inóculos de 400 y 800 células/cm2, por triplicado, para evaluar el efecto de los tratamientos de irradiación.
Migración celular (scratch wound healing)
Se incubaron las células hasta alcanzar el 90-100% de confluencia. Utilizando una punta de micropipeta estéril, se trazaron estrías sobre la superficie del cultivo y se monitoreó periódicamente al microscopio la migración y proliferación celular hasta el cierre de la estría (o hasta el cierre de la estría en el tratamiento control) (Ho, Mou, Chiang, Weng y Chow, 2005 y Rodríguez, Wu y Guan, 2005).
Tinción de ADN
Se cultivaron las células hasta un 60-70% de confluencia en frascos de 25 cm2. El día antes de la extracción, se agregaron 4 ml de medio de cultivo fresco (para favorecer la mitosis). Se agregaron 400 µl de colchicina (0,01g/L en PBS) y se incubó bajo condiciones normales por 45 min. Se cosecharon las células por el método de tripsina-EDTA, se centrifugaron a 1200 rpm por 5 min y se resuspendió el precipitado en 0,5 ml de PBS, al cual se le añadieron 2 ml de solución hipotónica (KCl 0,075M) y se incubó a temperatura ambiente por 20 min. Se fijó la muestra con 2 ml de una solución fijadora de metanol y ácido acético (3:1 v/v, 4°C) y se centrifugó a 1200 rpm por 5 min. Se descartó el sobrenadante y se resuspendió el precipitado en 3 ml de solución fijadora (4 °C). Se colocó una gota de la preparación sobre un portaobjetos precalentado a 37 °C y se dejó secar al aire. Se le agregó una gota de GIEMSA y se observó al microscopio (Multani et al., 1999 y Freshney, 2010); la resolución del método permite únicamente diferenciar entre presencia y ausencia de cromosomas en la muestra.
Análisis estadístico
Cuando fue pertinente, se calculó el promedio, la desviación estándar y el coeficiente de varianza de los tratamientos, los cuales se compararon mediante una ANOVA de un factor o una prueba T-student, para α = 0,05. Todos los análisis se realizaron con el programa Microsoft Excel.
Resultados y discusión
Las radiaciones ionizantes generan radicales libres e intermediarios reactivos de oxígeno, los cuales causan graves lesiones en las estructuras celulares, resultando en la incapacitación metabólica de la célula. El daño más crítico se produce al haber ruptura del ADN, lo cual induce a la muerte celular. Esta puede ocurrir por apoptosis (muerte celular programada) o necrosis (Arias, Aller, Arias y Lorente, 2001).
La respuesta celular a las radiaciones puede ser de cuatro tipos, incluyendo: (a) muerte en interfase, cuando ocurre el cese de las funciones metabólicas; (b) retraso mitótico, cuando se da un bloqueo temporal de las células dentro del ciclo mitótico; (c) fallo reproductivo o muerte diferida, lo cual ocasiona que las células irradiadas efectúen pocos ciclos antes de morir; y (d) modificación celular, cuando se da una transformación de la célula, la cual puede conservar su capacidad reproductiva y dar lugar a una neoplasia maligna (carcinogénesis) (Cura, Pedraza y Gayete, 2009). Para la obtención de una capa alimentadora es necesario que las células detengan su ciclo celular y mueran en un período relativamente corto de tiempo, aunque manteniendo sus funciones metabólicas y sin transformarse. Debido a ello, la muerte celular en interfase y la modificación celular son indeseables, siendo que la respuesta ideal esperada es la inactivación mitótica; sin embargo, el fallo reproductivo o muerte diferida podría ser una variante aceptable.
En el presente trabajo se evaluó el efecto de cuatro dosis de irradiación con rayos X (30, 60, 70 y 80 Gy) sobre fibroblastos murinos 3T3. Para determinar el efecto de estas dosis sobre la viabilidad celular, se utilizó la prueba de viabilidad por MTT, la cual se basa en la propiedad de esta sustancia de ser reducida a cristales de formazán (insolubles en agua) por los mecanismos de oxidación-reducción en la mitocondria de las células metabólicamente activas, generando un cambio de color cuya intensidad es proporcional a la cantidad de células viables presentes en la muestra (Freshney, 2010 y Montuenga, Ruiz y Calvo, 2009). Por otro lado, se emplearon ensayos de eficiencia de plateo y migración celular para determinar la capacidad de adhesión y supervivencia de las células, así como su capacidad de migración en el cultivo, respectivamente.
Dado que las células 3T3 son adherentes, su adhesión al recipiente de cultivo es indispensable para su supervivencia y eventual proliferación, mientras que su capacidad de migración es un indicador del mantenimiento de sus funciones diferenciadas in vitro. Los resultados obtenidos mostraron que las dosis de 60, 70 y 80 Gy permitieron generar células mitóticamente inactivas, con resultados óptimos obtenidos con la dosis de 80 Gy.
Dosis de irradiación: 30 Gy
Para la dosis de 30 Gy, se evalúo la concentración y viabilidad celular a lo largo de 22 días desde la irradiación. Se observó un crecimiento desordenado de las células y, lo más importante, la concentración celular se triplicó al segundo día de la irradiación, aunque decreció progresivamente con el tiempo (figura 1), lo que sugiere una respuesta del tipo fallo reproductivo o muerte diferida. Aunque tanto la concentración como la viabilidad celular mostraron una tendencia a decrecer progresivamente, a los 22 días de la irradiación todavía parecía existir actividad mitótica y metabólica, lo cual es indeseable para los requerimientos de nuestro laboratorio, ya que se busca que las células a utilizar como capas alimentadoras carezcan completamente de actividad mitótica y estén completamente ausentes al finalizar el período de 22 días.
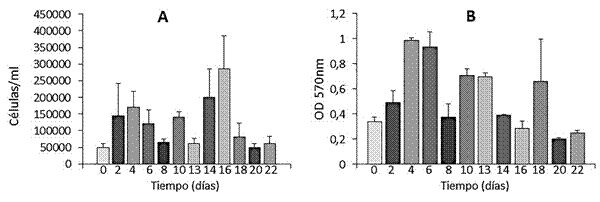
Figura 1 Concentración (A) y viabilidad celular (B) de fibroblastos 3T3 irradiados con una dosis de 30 Gy (rayos X). La viabilidad celular se determinó mediante MTT, donde la absorbancia a 570 nm es proporcional a la cantidad de células viables (B), mientras que la concentración celular se determinó mediante recuento al hematocitómetro (A). Cada grupo de datos representa el promedio de tres réplicas. La concentración celular se refiere solo a células vivas (azul tripán).
Aunque para la dosis de 30 Gy no se contó con un control de células no irradiadas, los controles de los ensayos de irradiación con otras dosis demostraron que la línea 3T3 utilizada, bajo condiciones normales, presenta un crecimiento congruente con la curva de crecimiento esperada para cualquier cultivo celular (Freshney, 2010), con una fase exponencial de crecimiento hasta los 6-10 días desde la inoculación o siembra y una fase estacionaria entre los 10-20 días, seguida por una fase de senescencia. Por lo tanto, las anomalías observadas en las células irradiadas con 30 Gy (rayos X) demuestran que esta dosis causó daños importantes en el metabolismo celular, pero no fue suficiente para inducir el arresto mitótico.
Dosis de irradiación: 60 y 70 Gy
Las células tratadas con 60 y 70 Gy (figuras 2 y 3) evidenciaron un aparente arresto mitótico, ya que no se encontró diferencia significativa (p>0,05) entre la concentración celular (de las células irradiadas) del día inicial respecto a la concentración celular del día final de cada ensayo. Por otro lado, aunque la viabilidad de las células irradiadas fue significativamente reducida (p<0,05) con respecto a los controles en todos los muestreos, se evidenció la presencia de células metabólicamente activas en los cultivos irradiados. Lo anterior sugiere que ambas dosis fueron efectivas para detener la división celular, manteniendo células metabólicamente activas hasta por 21 días (60 Gy) y 16 días (70 Gy). Sin embargo, no se pudo determinar el tiempo de pérdida completa de la viabilidad celular.
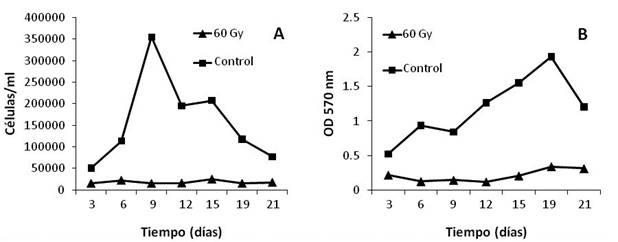
Figura 2 Concentración (A) y viabilidad celular (B) de fibroblastos 3T3 irradiados con una dosis de 60 Gy (rayos X). La concentración celular se determinó mediante recuento al hematocitómetro, mientras que la viabilidad celular se determinó mediante MTT. Cada punto representa el promedio de tres réplicas. La concentración celular se refiere solo a células vivas (azul tripán).
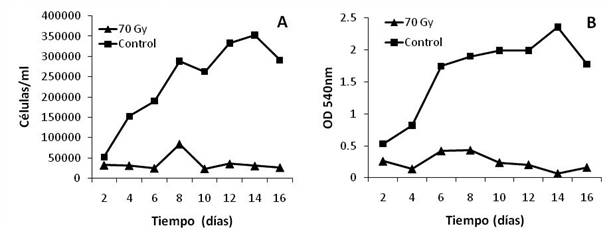
Figura 3 Concentración (A) y viabilidad celular (B) de fibroblastos 3T3 irradiados con una dosis de 70Gy (rayos X).
La concentración celular se determinó mediante recuento al hematocitómetro, mientras que la viabilidad celular se determinó mediante MTT. Cada punto representa el promedio de tres réplicas. La concentración celular se refiere solo a células vivas (azul tripán).
El efecto de la dosis de 70 Gy fue, además, evaluado mediante la observación de la eficiencia de plateo, prueba que ha sido definida como el mejor método para determinar la muerte celular reproductiva luego de tratamientos con radiaciones ionizantes (Franken et al., 2006). El tamaño de las colonias se correlaciona con la capacidad de proliferación celular, mientras que la cantidad de colonias se asocia con la supervivencia y la capacidad de adhesión. En este sentido, no fue posible obtener colonias aisladas y la mayoría de las células se concentraron en una única colonia en el centro del pozo; sin embargo, sin importar la cantidad de células sembradas en el inóculo inicial, hubo ausencia total de formación de colonias en las células irradiadas, mientras que los controles prácticamente alcanzaron la confluencia al finalizar el tiempo de incubación (figura 4). Esto implica que hubo una reducida adhesión y, por lo tanto, supervivencia de las células irradiadas, y a la vez comprueba que no hubo proliferación, por lo que se verifica la efectividad de la dosis de irradiación utilizada.
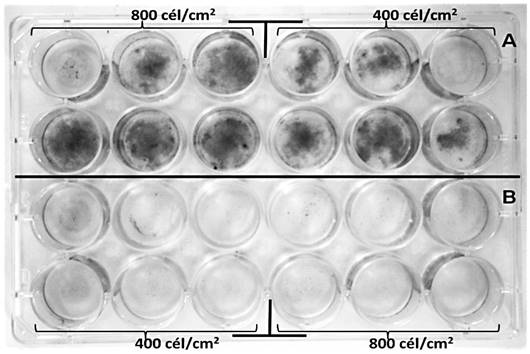
Figura 4 Eficiencia de plateo de fibroblastos 3T3 irradiados a 70 Gy (rayos X). Las células no irradiadas (control) se muestran en A, mientras que en B se observan las células irradiadas. Se presentan los datos observados a las dos semanas desde la irradiación e inoculación. Las manchas negras (cuyo color original es morado) son las células teñidas con cristal violeta.
Dosis de irradiación: 80 Gy
Todas las repeticiones demostraron que una dosis de 80 Gy fue suficiente para detener la división celular (figura 5), ya que no hubo aumentos significativos (p>0,05) en la concentración de las células irradiadas (excepto un aumento anómalo en el día 8 de la repetición 1), pero sí una disminución significativa (p<0,05) respecto al inóculo inicial (50.000 células/ml). Sin embargo, en el último día de muestreo en todas las repeticiones aún se contabilizaron algunas células metabólicamente activas en los cultivos. El arresto mitótico se verificó mediante la tinción de cromosomas de las células en la muestra, donde se evidenció la ausencia de cromatina condensada en las células irradiadas (figura 6).
Por otro lado, al igual que con las dosis de 60 y 70 Gy, la viabilidad de las células irradiadas a 80 Gy fue significativamente reducida (p<0,05) con respecto a los controles en todos los muestreos, excepto para el día 2 en todas la repeticiones y el día 4 en la repetición 2, donde no se encontró diferencia con los controles (p>0,05). Estos datos demuestran que las células irradiadas mantuvieron una actividad metabólica restringida, que decreció progresivamente hasta una reducción del 88% en relación con la viabilidad celular inicial, a los 25 días desde la irradiación.
Considerando que los queratinocitos pueden tardar aproximadamente de 2 a 5 semanas en alcanzar la confluencia en un cultivo primario (Atiyeh, Hayek y Gunn, 2005), se consideró que las células 3T3 tratadas con 80 Gy pueden conformar una capa alimentadora adecuada para el cultivo de epidermis.
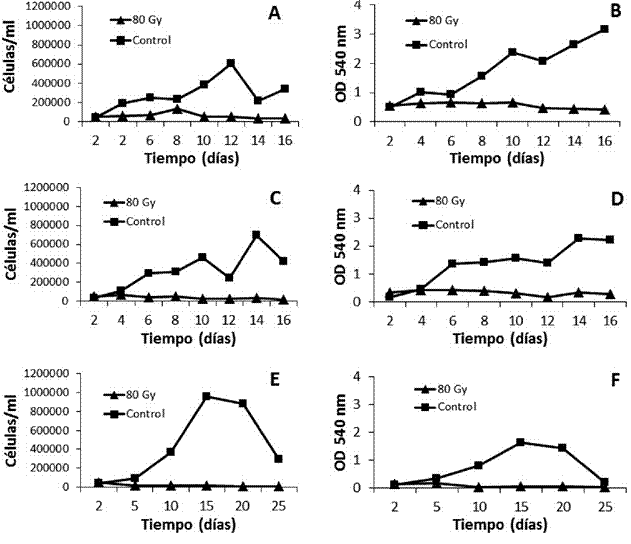
Figura 5 Concentración (A, C y E) y viabilidad celular (B, D y F) de fibroblastos 3T3 irradiados con una dosis de 80 Gy (rayos X). La concentración celular se determinó mediante recuento al hematocitómetro, mientras que la viabilidad celular se determinó mediante MTT. Cada punto representa el promedio de tres réplicas. La concentración celular se refiere solo a células vivas (azul tripán). A y B, repetición 1; C y D, repetición 2; E y F, repetición 3.

Figura 6 Tinción (GIEMSA) de núcleos de células tratadas con una dosis de irradiación de 80 Gy (rayos X) (1000x).
Se muestran los núcleos de las células a las 24 horas de la irradiación. En las células irradiadas (A) se observa la cromatina dispersa por todo el núcleo, mientras que en las células control no irradiadas (B) se observa la cromatina condensada. La condensación de la cromatina es una característica del período de mitosis celular.
Los ensayos de eficiencia de plateo de las células 3T3 tratadas con 80 Gy mostraron que, sin importar la cantidad de células sembradas en el inóculo inicial y, al igual que con la dosis de 70 Gy, hubo reducida o total ausencia de formación de colonias en las células irradiadas, lo que implica mínima adhesión y supervivencia de éstas, verificando la eficacia de la dosis de irradiación utilizada (figura 7). La resolución de las colonias en las células control (no irradiadas) fue mejor que en ensayos previos, demostrándose que una adecuada agitación de los frascos de cultivo durante las primeras horas después de la siembra mejoró la distribución de las colonias.
La mayor cantidad de colonias en los pozos centrales de los frascos de cultivo podría deberse al efecto de borde intrínseco de estos dispositivos; es posible que los pozos centrales mantengan mayor humedad respecto a los pozos periféricos. Esto implica que en futuros ensayos con estos frascos de cultivo se debe emplear la distribución al azar de las réplicas.
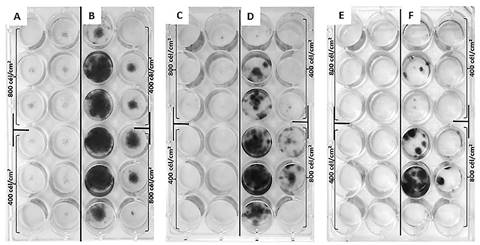
Figura 7 Eficiencia de plateo de fibroblastos 3T3 irradiados a 80 Gy (rayos X) en tres repeticiones. Las células no irradiadas (control) se muestran en B, D y F, mientras que en A, C y E se observan las células irradiadas. Se presentan los datos observados a las dos semanas desde la irradiación e inoculación. Las manchas negras (cuyo color original es morado) son las células teñidas con cristal violeta.
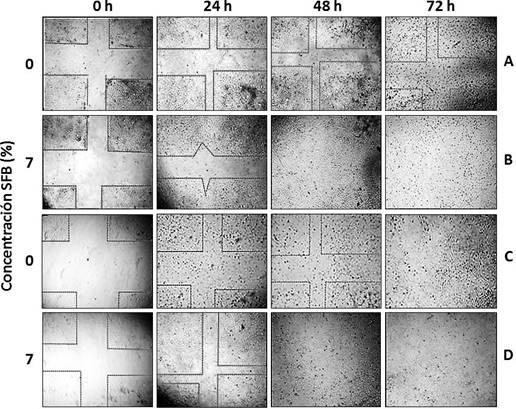
Finalmente, se realizaron pruebas de migración celular (scratch wound healing) (Ho et al., 2005), que permiten verificar el mantenimiento de la funcionalidad diferenciada de las células adherentes in vitro a través de la observación de su comportamiento motil, ya que las células en cultivo, aunque se trate de células adherentes como los fibroblastos, son móviles y pueden recorrer o migrar distancias significativas a través del sustrato (Freshney, 2010). Esta movilidad depende de la densidad celular y la presencia de agentes como factores de crecimiento. La migración celular cesa cuando las células alcanzan confluencia, lo cual va acompañado de una reducción en la tasa de división celular (inhibición por contacto) (Freshney, 2010).
Este tipo de ensayos permite simular in vitro el proceso de reparación de una herida o lesión, pudiéndose observar la capacidad de migración y proliferación de las células en cuestión para realizarlo. Al realizar una estría en cultivos con medios que contienen suero, la migración es acompañada por proliferación celular, mientras que en medios de cultivo carentes o deficientes en suero, se reduce o inhibe la proliferación celular, de manera que la repoblación de la estría se debe casi exclusivamente a la migración de las células (Ho et al., 2005).
Al evaluar la migración celular de fibroblastos 3T3 irradiados con una dosis de 80 Gy (rayos X) y cultivados en ausencia y presencia de SFB (figura 8), se encontró que la invasión del sitio de la lesión es al azar y que, en presencia de suero, tanto en las células irradiadas como en las células control, la zona de la lesión es repoblada a las 48 horas, mientras que se aprecia un leve retraso cuando se elimina el SFB, registrándose una migración más lenta. No obstante, no se apreciaron diferencias observables entre las células control y las células irradiadas, lo cual indica que las células irradiadas mantuvieron propiedades de migración similares a las del control.
Conclusiones
Se verificó que la irradiación con rayos X es un método viable y efectivo para la reducción de la supervivencia y proliferación celular en los fibroblastos murinos 3T3, ya que una dosis de radiación correspondiente a 80 Gy generó daños suficientes en la línea celular para causar el arresto mitótico, pero manteniendo una actividad metabólica basal durante 25 días. A pesar de la irradiación, los fibroblastos mantuvieron su capacidad de migración, sugiriendo que las células tratadas conservan algunas de sus propiedades diferenciadas in vitro.

