










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgronomía Costarricense
versión impresa ISSN 0377-9424
Agron. Costarricense vol.39 no.1 San Pedro de Montes de Oca ene./jun. 2015
Herencia y relación genética asociados al rendimiento, madurez en híbridos de tomate [Solanum lycopersicum L. (Mill.)]
Resumen
Durante 2010-2011 se evaluaron en invernadero, 30 híbridos de tomate, con el objetivo de determinar las relaciones genéticas entre los distintos caracteres afines al rendimiento, el número de días a la madurez y duración en anaquel. Los resultados mostraron que el híbrido 5 fue el mejor en textura, para grados brix los mejores híbridos fueron: 2, 29, 25, 16 y 10, para número de días a la madurez, el híbrido 12. El híbrido 8 para tamaño y ancho de fruto, para longitud de fruto el híbrido 4 y el híbrido 9 para rendimiento. En el pH hubo efectos adicionales y en la textura de fruto, los grados brix y el número de días a la madurez, efectos dominantes. En el rendimiento, tamaño, longitud y ancho de fruto hubo efectos aditivos y dominantes. La herencia fue mayor para grados brix, tamaño, longitud y ancho de fruto. El progenitor 70 es portador de genes para menor pérdida de textura, mayor ganancia de grados brix, mayor pérdida de pH, mayor tamaño y mayor rendimiento de fruto. El progenitor 71 fue el portador de genes con menor número de días a la madurez, mayor longitud y mayor ancho de fruto. La correlación mostró una relación moderada y negativa (r=-0,62), entre tipo de crecimiento de la planta y forma predominante de fruto; además hubo correlaciones moderadas y negativas (r=-0,51) entre forma predominante de fruto, color exterior del fruto maduro y número de lóculos (r=-0,41). Se identificaron 2 componentes principales: el primero relacionado con textura de fruto, grados brix, tamaño, longitud y ancho de fruto; el segundo componente identificó al pH, número de días a la madurez y rendimiento por planta. La correspondencia múltiple mostró que en los híbridos evaluados predominan frutos redondos con pericarpio y la parte externa de color rojo.
Palabras clave: Anaquel; genes efectos aditivos; efectos dominantes; heredabilidad.
Abstract
In the year 2010-2011, in the greenhouse, 30 tomato hybrids were evaluated to determine the genetic relationships between the different characters related to yield, number of days to maturity and shelf life. Results showed that hybrid 5 was the best for texture. For brix degrees, the best hybrids were: 2, 29, 25, 16 and 10. Hybrid 12 for number of days to maturity. Hybrid 8 for fruit size and width. For fruit length, hybrid 4; and hybrid 9 for yield. The pH is controlled by additive effects, and in fruit texture, degrees brix and the number of days to maturity, dominant effects are involved. In yield, size, length and width of fruit there were additive and dominant effects involved. Inheritance was higher for degrees brix, size, length and width of fruit. Progenitor 70 carries genes for less loss of texture, higher gain of degrees brix, greater loss of pH, larger size and greater fruit yield. Instead, parent 71 is the carrier of genes for fewer days to maturity, greater length and greater width of the fruit. Correlation analysis showed a moderate and negative (r=-0.62) and relationship between type of plant growth and predominant fruit shape. Also, moderate and negative correlations (r=-0.50) were observed between dominant form of fruit, external color of the ripe fruit and number of locules (r=-0.41). Characterization showed a wide variability in quantitative variables. Two main components, were identified: the first related to fruit texture, brix degrees, size, length and width of fruit; the second component identified pH, number of days to maturity and yield per plant. Multiple correspondence analyses indicated that in the hybrids evaluated predominate round fruits with pericarp and the outside colored red.
Keywords: Shelf; genes; additive effects; dominant effects; heritability.
Introducción
La identificación y selección de líneas parentales son necesarias en cualquier programa de hibridación para generar nuevos y potenciales genotipos valiosos para combinar y lograr los efectos de genes deseados en una línea homocigótica. En ese aspecto la información relativa de los distintos tipos de acción génica, la magnitud relativa de la variación genética y la combinación de las estimaciones de habilidad combinatoria son parámetros importantes y vitales para lograr la composición genética de los cultivos (Hannan et ál. 2007).
La información es importante y podría proveer de una estrategia esencial para los mejoradores de tomate en la selección de las mejores combinaciones parentales para continuar el proceso de mejoramiento. La explotación de la heterosis depende principalmente de la detección y selección de germoplasma que se podría producir por las mejores combinaciones para los más importantes caracteres agronómicos. Toda la variabilidad genética observada en el análisis de cada carácter se fraccionó en sus componentes, es decir, la aptitud combinatoria general (ACG) y la aptitud combinatoria específica (ACE) tal como lo define Sprague (1966) y los efectos recíprocos, descritos por Griffing (1956). Ellos afirmaron que los efectos de ACG fueron debido a un tipo de acción génica de tipo aditivo y los efectos de ACE se debieron a la acción génica no aditiva (dominante o epistático).
Muchos estudios de la aptitud combinatoria para los componentes de rendimiento fueron evaluados en muchas especies. Algunos encontraron la predominancia de la ACG y que podría ser más importantes que la ACE (Khan et ál. 1991, Yaqoob et ál. 1997), entre tanto otros sugirieron que la ACE es más importante (Ortiz y Golmirzaie 2004, Biswas et ál. 2005).
El presente estudio tuvo como objetivo determinar las relaciones genéticas entre los componentes asociados al rendimiento, el número de días a la madurez y duración en anaquel para 30 híbridos de tomate.
Materiales y Métodos
El experimento se implementó en un invernadero de la Fundación PROINPA en la zona de El Paso en Cochabamba, geográficamente ubicada a 17°21’00.98” de latitud sur y de 66°15’47.58” de longitud oeste, a una altitud de 2617 msnm. Esta zona tiene una temperatura media anual de 18°C, con 55% de HR y una precipitación media anual de 569,9 mm.
Se utilizaron 30 híbridos de tomate (Cuadro 1) obtenidos en la Fundación PROINPA de la cruza entre 21 líneas parciales con 3 ciclos de autofecundación, una línea silvestres de Solanum lycopersicum con resistencia al tospovirus o peste negra (TSWV): 70 (89R Sw-5/Sw-5) y una línea susceptible a tospovirus: 71 (895 Laching Sw-5) (Gabriel et ál. 2013).
Las 21 líneas parentales fueron seleccionadas de 74 accesiones de la colección de trabajo de la Fundación PROINPA que fueron las mismas evaluadas por sus atributos de rendimiento, resistencia a enfermedades y otros caracteres agronómicos de interés (Gabriel et ál. 2008). Las líneas parentales provinieron de diferentes empresas productoras de tomate como Reuter Seeds (España), Enza Zaden (Holanda), Sperli (Holanda), Numhems Zaden (Holanda), Seminis (Chile), NeikerTecnalia (España) y del Programa Nacional de Hortalizas del INIAF (Bolivia).
El suelo se preparó manualmente, con una remoción y desterronado de las platabandas; luego se realizó la aplicación de fertilizante nitrofoska en una cantidad de 2,5 kg por platabanda. Se hicieron platabandas niveladas de 1 x 28,50 x 0,15 m.
Antes de la siembra se realizó el lavado de las bandejas de plastoformo con agua y detergente, luego se realizó la desinfestación con una solución de agua y cloro (50 ml de Cl/1 l de agua) durante 5 a 10 min. Se preparó sustrato en una relación: 1:1:1 de cascarilla de arroz, tierra vegetal y arena fina que se esterilizó en una caldera a vapor durante 35 min a 90°C. Se llenó los hoyos de las bandejas compactándola para evitar la formación de bolsas de aire. Se realizó la siembra en un hoyo de 0,5 a 1 cm de profundidad y un diámetro de un lápiz donde se depositó una semilla por hoyo y 55 semillas por tratamiento, finalmente se cubrió con lama desinfestada y con riego a capacidad de campo.
El transplante se realizó en sistema de plantación tresbolillo, para lo que se cavaron hoyos a una profundidad de 0,15 m a una distancia de 0,46 m entre hoyos dentro de la hilera y una distancia de 0,60 m entre surcos.
El control de malezas se realizó manualmente. Se controlaron plagas y enfermedades. En las plantas de hábito de crecimiento determinado, la poda se realizó en 3 ramas principales con eliminación de las ramas restantes. Las hojas viejas y los brotes se dejaron para la formación de ramas nuevas debido a que las 3 ramas principales terminan en una inflorescencia. En las indeterminadas, la poda se realizó en 2 ramas principales y se eliminó las ramas restantes. En este caso las hojas viejas y los brotes se eliminaron para evitar la formación de otras ramas secundarias. El tutoraje se realizó después de la poda. En las determinadas se colocaran 3 tutores, un tutor por cada rama principal. En las indeterminadas también se instaló un tutor por rama principal.
Para el análisis de anaquel se recolectaron 18 frutos por híbrido. Los frutos fueron recolectados en el estado fenológico de 1/3 verde y 2/3 maduro (pintonas). Se implementaron 8 tratamientos para evaluar la duración en anaquel y composición química de los frutos: 0, 3, 6, 9, 13, 16 y 21 días en anaquel. Las variables evaluadas fueron: el color, el peso, la textura, los grados brix y el pH.
Variables de respuesta
Las variables de respuesta de las características morfológicas, agronómicas y agroindustriales fueron evaluadas según las recomendaciones del IPGRI (2008). Las variables cuantitativas fueron: tamaño de fruto (TmF), longitud de fruto (LnF), ancho de fruto (AnF) y rendimiento (Y). Las variables agronómicas fueron: número de días a la floración (NoDM). Las variables para duración en anaquel fueron: pérdida de textura (PTXT), pérdida de pH (PpH) y grados brix (G°Bx). Las variables cualitativas fueron: tipo de crecimiento de la planta (TCP), forma predominante del fruto (FPF), color exterior del fruto maduro (CFM), color de la carne del pericarpio (CCP), forma del corte transversal del fruto (CTF) y número de lóculos (NOL).
Diseño experimental y análisis estadístico
El estudio de durabilidad en anaquel se implementó por medio de un diseño completamente aleatorio.
El estudio de las características morfológicas, agronómicas y de resistencia a estreses bióticos y abióticos se implementó en diseño de bloques completos al azar con submuestreo con 4 repeticiones. Los tratamientos fueron los híbridos de tomate, que fueron distribuidos en 4 bloques con 10 plantas por híbrido por unidad experimental y 3 plantas para el submuestreo.
Sobre la base del modelo definido, se realizaron análisis de varianza para probar hipótesis acerca de los efectos fijos y comparaciones de medias de los híbridos. El análisis de varianza también sirvió para estimar los componentes de varianza para los efectos aleatorios. Los análisis indicados se realizaron por medio del Proc GLM del SAS (SAS 2004).
Para el análisis genético de los híbridos se utilizó el Diseño I de Carolina de Norte, en el que cada progenitor masculino se apareó con m hembras (Martínez 1988). El análisis de este diseño permitió estimar las varianzas genéticas de las variables de respuesta y determinar la aptitud combinatoria general (ACG) del macho. También se estimó la herencia en sentido estrecho (h2) y amplio (H2).
Finalmente se realizaron análisis entre correlación entre las variables, así como un análisis de componentes principales para las variables cuantitativas y un análisis de correspondencia múltiple para las variables cualitativas (López e Hidalgo 2003).
Resultados y Discusión
Análisis de varianza en anaquel
En el análisis de varianza, (Cuadro 2) se encontraron diferencias significativas (p<0,05) entre híbridos para las variables pérdida de textura de fruto y ganancia en grados brix; esto indica que la respuesta de al menos uno de los híbridos fue diferente para cada variable evaluada. En cambio, la variable pH no presentó ninguna diferencia significativa (p<0,05), esto indica que no existieron diferencias en el pH en ninguno de los híbridos. Por otro lado la comparación de medias al p<0,01 de probabilidad, para la variable PTxT, mostró que el híbrido con menor pérdida de textura o mayor firmeza de fruto fue la cruza 5. Por otra parte, los híbridos con mayor ganancia en grados Brix fueron los híbridos 21, 29, 25, 16 y 10. En cambio, para la variable pH no existieron diferencias significativas entre híbridos.
Componentes de varianza genética en anaquel
En la estimación de varianzas genéticas estimados (Cuadro 3) para pérdida en pH mostró que la varianza aditiva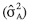 es mayor a cero. Esto indicaría que la pérdida en pH estaría gobernada por efectos aditivos. En cambio, para pérdida de TxT y ganancia en grados brix la varianza de dominancia
es mayor a cero. Esto indicaría que la pérdida en pH estaría gobernada por efectos aditivos. En cambio, para pérdida de TxT y ganancia en grados brix la varianza de dominancia 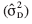 , fue mayor a cero. Esto significaría que la pérdida en TxT y ganancia en grados brix estarían gobernados por efectos de dominancia. Para las variables pérdida en textura de fruto y ganancia en grados brix, los efectos de dominancia fueron un número infinito de veces más importantes que los efectos aditivos.
, fue mayor a cero. Esto significaría que la pérdida en TxT y ganancia en grados brix estarían gobernados por efectos de dominancia. Para las variables pérdida en textura de fruto y ganancia en grados brix, los efectos de dominancia fueron un número infinito de veces más importantes que los efectos aditivos.
Por otra parte, en el caso de la pérdida en pH los efectos aditivos fueron más importantes respecto a los efectos de dominancia. Resultados similares fueron hallados por Gragera (2003), quien encontró que el pH, tuvo una mayor expresión de genes de efectos aditivos (d/a=0,03) durante un estudio de herencia en tomates de industria de origen español. Asimismo, en los mismos genotipos encontró que los efectos de dominancia (d/a=2,76) fueron más importantes para grados brix. Esto significaría que hay genes mayores y menores involucrados en la herencia de estos caracteres. Por otro lado, para TxT resultados similares fueron hallados por Pereira da Costa et ál. (2009), quienes reportaron que en accesiones de S. lycopersicum, existe una mayor expresión de genes con efectos de dominancia, donde los efectos de dominancia fueron 1,33 veces más importantes que los efectos aditivos.
La estimación de la heredabilidad a partir de los efectos genéticos (Cuadro 3), mostró una heredabilidad en sentido amplio alta para ganancia en grados brix, lo que sugiriere que existe una amplia variación genética y pobre variación ambiental para este carácter. En cambio existe una baja heredabilidad en sentido amplio para pérdida en textura. Esto propone que este carácter, estuvo ampliamente influenciado por el ambiente. Por otra parte, la heredabilidad en sentido estrecho para estas 2 variables fue igual a cero; dicho aspecto confirma que estos caracteres estarían gobernados solo por efectos de dominancia. Resultados similares fueron reportados en tomates mexicanos evaluados por Hernández (2013). En dicho experimento hubo una alta heredabilidad en sentido amplio (H2>0,5) y una baja heredabilidad en sentido estrecho (h2<0,5) para grados brix. Por otra parte, Moreno et ál. (2002), encontraron una heredabilidad en sentido estrecho alta (h2>0,45) para grados brix, que indica que los efectos aditivos son de importancia en esta variable. Esto sugeriría que este carácter estaría gobernado tanto por efectos aditivos como de dominancia. Pereira da Costa et ál. (2009) también encontraron una heredabilidad en sentido estrecho similar a la del presente trabajo (h2<0,1) para textura de fruto. En cambio la heredabilidad en sentido amplio fue moderada (H2=0,39), lo que confirma los efectos de dominancia más importantes. Para la variable pH, las heredabilidades en sentido amplio y estrecho fueron bajas e idénticas, aspecto que señala la no existencia de efectos de dominancia para esta variable (tal como se observó en la importancia de los efectos genéticos) y que está muy influenciada por el ambiente. En cambio, Moreno et ál. (2002) encontraron una heredabilidad en sentido estrecho alta (h2>0,45), que indica una baja influencia del medio ambiente. De todos modos, se confirma la importancia de los efectos aditivos para esta variable.
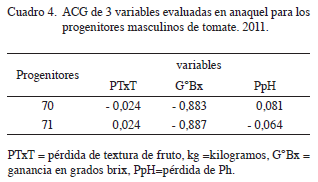
ACG de los machos para la prueba en anaquel
La estimación de la ACG de los machos (Cuadro 4), mostró variabilidad en el comportamiento de los progenitores para las diferentes variables de respuesta en anaquel. El progenitor 70 mostró la menor ACG para pérdida en textura de fruto. Asimismo, tuvo las mayores ACG para ganancia en grados brix y para pérdida en pH, que sugiriere a este progenitor como el portador de genes para menor pérdida de firmeza de fruto, ganancia en grados brix y mayor pérdida en pH del jugo de tomate en anaquel durante 21 días en el mismo ambiente. Estudio realizado por Lagos (2012), en la evaluación de material de tomate procedente de Colombia y Ecuador, reportó que las varianzas de los efectos de ACG fueron relativamente bajas (menores a uno) en pH, sólidos solubles totales (grados brix) y textura de fruto, indica de que esto se debe a la uniformidad genética de los parentales en estas variables. En resumen, el parental 70 aporta genes para menor pérdida de textura de fruto, mayor contenido de sólidos solubles (grados brix) y mayor pérdida de pH de fruto. Por tanto, este progenitor es una buena fuente para desarrollar futuros programas de mejoramiento sobre la vida en anaquel de frutos de tomate.
Análisis de varianza morfológico y agronómico
El análisis de varianza para híbridos (Cuadro 5) mostró diferencias altamente significativas (p<0,01) para las variables número de días a la madurez, longitud de fruto, ancho de fruto, tamaño de fruto y rendimiento; esto significa que el comportamiento de al menos uno de los híbridos evaluados es diferente en cada una de las variables. La comparación de medias (p<0,01) mostró que el híbrido 12 alcanzó la menor cantidad de días a la madurez, el híbrido 8 obtuvo el mayor tamaño de fruto, el híbrido 4 exhibió la mayor longitud de fruto, los híbridos 8,17, 6 y 14 obtuvieron el mayor ancho de fruto y 9 fue el híbrido con mayor rendimiento de fruto.
Componentes de varianza genética para variables morfológicas y agronómicas
Los componentes de varianza genética estimados (Cuadro 6), para número de días a la madurez muestran que la varianza de dominancia es mayor a cero. Esto indica que este carácter está gobernado solo por efectos de dominancia como lo indica la importancia de los efectos genéticos (γ), donde los efectos de dominancia son un infinito número de veces más importantes que los efectos aditivos. En cambio, para tamaño de fruto, longitud de fruto, ancho de fruto y rendimiento las varianzas aditiva y de dominancia son mayores a cero, lo cual indica que tanto efectos de dominancia como aditivos están involucrados en herencia de las 4 variables. Sin embargo, al considerar la importancia de los efectos genéticos (γ) para tamaño de fruto, los efectos de dominancia son 5,4 veces más importantes respecto a los efectos aditivos, para longitud de fruto, los efectos de dominancia son 5,6 veces más importantes respecto a los efectos aditivos, para el ancho de fruto, los efectos de dominancia fueron 1,39 veces más importantes que los efectos aditivos y finalmente para el rendimiento, los efectos de dominancia fueron 1,81 veces más importantes respecto a los efectos aditivos. Resultados similares fueron hallados por Hernández (2013), en genotipos mexicanos, quien reportó una mayor expresión de genes con efectos de dominancia parcial para los caracteres precocidad, longitud, ancho, y tamaño de fruto. Así mismo, un efecto similar fue observado por Grandillo y Tanksley (1996), en el tamaño de fruto y por Burdick (1954), en la precocidad. Sin embargo, dicha dominancia suele ser relativa en variables de precocidad debido a que su expresión depende de la etapa fisiológica y condiciones ambientales (Burdick 1954). Por otra parte, para rendimiento se encontraron efectos (Meneses 2002), donde la varianza de dominancia para peso de fruto, presentó valores muy bajos y/o negativos, lo que permite considerar que la varianza genética total estuvo determinada por efectos aditivos de los genes, lo que es muy favorable y factible de explotarse a través de cualquier esquema de selección. Estos resultados demuestran que la precocidad, tamaño, longitud, ancho y rendimiento de fruto, estarían controlados principalmente por genes con efectos de dominancia.
La estimación de la heredabilidad a partir de los efectos genéticos (Cuadro 6), mostró valores altos para la heredabilidad en sentido amplio (H2) para los variables tamaño, longitud y ancho de fruto, lo que sugiriere la presencia de alta variabilidad genética e insignificante variación ambiental. En cambio, las heredabilidades en sentido estrecho fueron bajas para todas las variables en general, lo cual indica que los efectos aditivos son de poca importancia, y plantea que la selección de híbridos de tomate sería más efectiva a través del tamaño, longitud y ancho de fruto. Los valores altos para la heredabilidad en sentido amplio y bajos para heredabilidad en sentido estrecho de estas 2 variables confirman que los efectos de dominancia son más importantes que los efectos aditivos. Estudios en tomates mexicanos realizados por Gaspar et ál. (2012) coinciden con los resultados encontrados en el presente trabajo, que identifica la heredabilidad en sentido amplio, como alta (H2>0,5) para longitud de fruto y baja para peso de fruto. Por otro lado, para ancho de fruto valores semejantes de heredabilidad (H2=0,66) en sentido amplio fueron estimados por (Hernández 2013). En general, la investigación realizada presenta las heredabilidades en sentido estrecho como bajas para las 5 variables (Cuadro 6). Resultados similares fueron encontrados por Pérez et ál. (2010), en material de Cuba y Florida, donde encontraron heredabilidades en sentido estrecho bajas para las variables longitud, ancho y tamaño de fruto, esto se debe a que la componente de varianza aditiva tiene un valor bajo, por lo que la transferencia genética es baja y la determinación de este carácter está influida por las condiciones ambientales en que se desarrollan los individuos. Así mismo, para la variable número de días a la madurez la heredabilidad en sentido amplio y estrecho concuerdan con los resultados encontrados en accesiones de Solanum lycopersicum en material de origen Argentino (Pereira da Costa et ál. 2009), quiénes obtuvieron una heredabilidad en sentido amplio intermedia (H2=0,36) y una heredabilidad estrecha nula (h2=0) para número de días a la madurez.
ACG para machos de variables morfológicas y agronómicas
Las ACGs estimadas para los 2 progenitores machos (Cuadro 7), mostró una amplia diferencia en el comportamiento de los progenitores en las diferentes variables de respuesta. El progenitor 71 mostró la menor ACG para número de días a la madurez, que sugiriere al progenitor como el portador de genes para menor número de días a la madurez. En cambio, el progenitor 70 mostró las mayores ACG’s para el tamaño de fruto y peso de fruto, lo que propone a este progenitor como el portador de genes para mayor tamaño y rendimiento de fruto. Por otra parte, el progenitor 71 mostró las mayores ACGs para longitud y ancho de fruto; por tanto, este progenitor sería el portador de genes para mayor longitud y ancho de fruto. Los valores de mejora de ACG con el parental 70 para Y y TmF son 0,114 y 0,005; por tanto, este progenitor contribuyó en general con 114 gramos al rendimiento por planta y en 0,13 mm al tamaño de los híbridos. En cambio, con el parental 71 los valores de mejora son 0,555 y 0,484 para LnF y AnF, lo cual significa que este progenitor contribuyó con 555 y 484 micrómetros a la longitud y ancho de fruto de los híbridos respectivamente. Asimismo, este parental contribuyó con la reducción de 1,8 días para NoDM. Resultados similares, fueron encontrados por (Sahagún et ál. 1999), en Physalis ixocarpa Brot. Donde hubo diferencias significativas (p≤0,05) en las ACGs de 3 genotipos para precocidad expresada a través de rendimiento en la primera cosecha, con el genotipo Salamanca que aportó mayor peso en la primera cosecha.
Análisis de correlación de variables cuantitativas y cualitativas
En el análisis de variables cuantitativas en anaquel no hubo ninguna correlación significativa al pr<0,01 de probabilidad entre las variables pérdida en textura de fruto, ganancia en grados brix y pérdida en pH. Murillo et ál. (2004), mencionan que ante la ausencia de correlación se debe realizar un análisis multivariado de Factores Principales de variación, que permite reducir la dimensionalidad de las variables, permite explorar la acción conjunta de las variables y determinar el efecto de cada variable en presencia de las otras (Uriel y Aldás 2005).
En el análisis de caracteres cualitativos agromorfológicos hubo una correlación significativa entre las variables TCP y FPF. Esta correlación fue moderada y negativa (r=-0,62). Esto indicó que la forma predominante del fruto depende del tipo de crecimiento de la planta. En las indeterminadas predominan los frutos redondos y redondos achatados (92%) y el 5% fueron redondos alargados. En cambio, en las determinadas el porcentaje de frutos redondos alargados fue mayor (30%) que en las indeterminadas y el porcentaje de frutos redondos y redondos achatados fue inferior que en las indeterminadas (65%). Asimismo, se presentaron frutos cordiformes (tipo pera) en un 5%. Resultados similares fueron hallados por Florido et ál. (2008), cuando cruzaron genotipos indeterminados de frutos redondos achatados con genotipos determinados de frutos redondos alargados. En las progenies indeterminadas se encontró un 85% de frutos redondos achatados y un 15% frutos redondos alargados. Por otra parte, en las progenies determinadas se encontró frutos redondos alargados en un 40% y frutos redondos achatados en un 60%. Por otra parte, la variable FPF se correlacionó negativa y moderadamente con la variable CFM (r=-0,51). Esto significaría que entre los frutos alargados predomina el color naranja y entre los frutos redondos los colores rojo y rosado. También esta variable FPF se correlacionó moderada y negativamente con la variable NOL (r=-0,41). Esto indicó que en los frutos alargados predomina la presencia de 3 lóculos por fruto y que en los frutos redondos predomina la presencia de 4 lóculos en la mayoría de los híbridos. Tales resultados concuerdan con los reportados por Medina y Lobo (2001), quiénes encontraron de 2 a 3 lóculos/fruto en frutos alargados y de 4 a 5 lóculos/fruto en frutos redondeados en la mayoría de las introducciones evaluadas.
En los caracteres cuantitativos agromorfológicos se encontró una correlación moderada y positiva se dio entre las variables LnF con AnF (r=0,67), esto significaría que a mayor longitud de fruto sería más ancho el fruto. Por otra parte, TmF estuvo moderadamente y positivamente correlacionada con LnF (r=0,56) y AnF (r=0,55). Por tanto, entre los híbridos evaluados, los híbridos con mayor tamaño de fruto exhibieron mayor ancho y longitud de fruto. Por otro lado hubo una correlación positiva baja entre las variables TmF y Y (r=0,35). Esto permite deducir que a mayor tamaño de fruto existe mayor rendimiento por planta. Hernández (2013), también encontró una correlación moderada y positiva (r=0,40) entre tamaño de fruto y rendimiento. Así mismo, Lippman y Tanksley (2001), encontraron una correlación positiva entre tamaño y rendimiento de fruto en poblaciones derivadas de S. pimpinellifolium y genotipos de S. lycopersicum de fruto grande. En nuestro caso la selección para mayor rendimiento y tamaño de fruto se podría realizar con base en híbridos de mayor longitud y ancho de fruto.
Análisis de componentes principales (ACP)
Una vez generados los componentes, se utilizaron los criterios propuestos por Pla (1986), Se retuvo 2 componentes cuyo valor propio fue ≥1 y que expresaron más del 70% de la varianza total (López e Hidalgo 2003). En el análisis de anaquel el primer componente contribuyó con el 41,19% de la varianza total. Aportó en forma positiva las variables PTxT y G°Bx; esto denotó que dicho componente identificó a los híbridos con mayor pérdida en textura y ganancia en contenido de grados brix o azúcar en los frutos. Resultados similares fueron hallados por Bonilla (2013), donde también encontró que la textura o firmeza de fruto fue una de las variables que más contribuyó a este componente. Por otro lado, la contribución de los grados brix a este componente concuerda con los estudios realizados por Salgado (2010), con material de México, quien encontró que el componente uno explicaba más del 46% de la variabilidad de las características evaluadas donde el porcentaje de los sólidos solubles fue muy importante para este componente. El segundo componente contribuyó con el 34,28% de la varianza total, en el cual la variable pH aportó positivamente. Por lo que este componente distinguió a los híbridos con mayor pérdida en contenido de pH.
En el análisis morfológico el primer componente contribuyó con el 52,35% del total de la varianza y aportó en forma positiva las variables TmF, LnF y AnF. Esto denotó que este componente identificó a los híbridos con mayor tamaño de fruto, longitud de fruto y ancho de fruto. Bonilla (2013), en materiales de los estados de Puebla y Oaxaca, encontró que el tamaño de fruto, longitud de fruto y ancho de fruto fueron las variables que más contribuyeron a este componente. Por otro lado, el segundo componente contribuyó con el 17,53% de la varianza total, en el cual la variable Y aportó positivamente. Por otra parte, la variable NoDM, contribuyó en forma negativa. Por lo que este componente distinguió a los híbridos con mayor rendimiento y precocidad.
Análisis de correspondencia múltiple
El análisis de correspondencia múltiple mostró variabilidad genética dentro de los 30 híbridos de tomate. Aunque la mayoría de los híbridos se agruparon por características similares, existieron casos aislados como el híbrido 20 que no se agrupo con ninguno de los híbridos. Este híbrido presenta frutos con color exterior rosado y carne del pericarpio del mismo color. Por otro lado, los híbridos 2, 17 y 23 se agruparon al presentar frutos redondos alargados con un color exterior naranja y la carne del pericarpio del mismo color. Otros híbridos que se diferenciaron fueron los híbridos 3, 5, 18, 27 y 28 al presentar forma redonda y un color exterior rojo y la carne del pericarpio también del mismo color. En general se observó que hubo una correlación entre la forma predominante del fruto, el color exterior y el color del pericarpio.
Conclusiones
En las variables de anaquel se encontró que la pérdida en textura de fruto y ganancia en grados brix están gobernados solo por efectos de dominancia. La pérdida en pH está gobernada solo por efectos aditivos. En las variables morfológicas se observó que el número de días a la madurez está gobernado solo por efectos de dominancia. En cambio, el tamaño de fruto, longitud de fruto, ancho de fruto y rendimiento de fruto están gobernados tanto por efectos aditivos y de dominancia.
Las heredabilidades en sentido estrecho para las 3 variables de anaquel fueron bajas. La heredabilidad en sentido amplio fue alta para ganancia en grados brix, moderada para pérdida en textura y baja para pérdida en pH. Las heredabilidades en sentido estrecho fueron bajas para las 5 variables morfológicas y agronómicas. Las heredabilidades en sentido amplio fueron bajas para número de días a la madurez y rendimiento y altas para tamaño, longitud y ancho de fruto.
El progenitor 70 fue portador de genes para menor pérdida de textura de fruto, mayor ganancia en grados brix y mayor pérdida en pH, mayor tamaño de fruto y mayor rendimiento de fruto.planta-1. En cambio, el progenitor 71 fue portador de genes para menor número de días a la madurez, mayor longitud y ancho de fruto.
El análisis de correlación entre pérdida en textura de fruto, ganancia en grados brix y pérdida en pH no mostró ninguna correlación significativa entre las 3 variables. Entre las variables morfológicas se observó una moderada correlación entre la forma predominante del fruto maduro con las variables: tipo de crecimiento de la planta, color exterior de fruto maduro y número de lóculos del fruto. Asimismo, el rendimiento esta moderadamente correlacionada con las variables tamaño, longitud y ancho de fruto. Las 2 últimas variables están altamente correlacionadas entre sí.
El análisis de componentes principales de las variables en anaquel, permitió generar 2 nuevos componentes. El primer componente permitió distinguir a los híbridos con menor pérdida de textura de frutos y mayor contenido de grados brix. El segundo componente identificó a los híbridos con mayor pérdida pH. Por otra parte, el análisis de componentes principales de las variables agromorfológicas, generó 2 nuevos componentes. El primer componente distinguió a los híbridos con mayor tamaño, longitud y ancho de fruto. El segundo componente identificó a los híbridos con mayor precocidad y rendimiento de fruto.
El análisis de correspondencia múltiple permitió distinguir que los híbridos de tomate evaluados a la madurez, se relacionan con la forma predominante de los frutos, con el color exterior y el color del pericarpio.
Agradecimientos
Se agradece al apoyo económico del proyecto “Fortaleciendo capacidades de innovación participativa para luchar contra la pobreza rural” (IP – Holanda), para la realización de esta investigación.
Literatura Citada
1. Facultad de Ciencias Agrícolas, Pecuarias, Forestales y Veterinarias de la UMSS, Casilla 3392, Cochabamba, Bolivia.
2. Facultad de Ciencias Agrícolas, Pecuarias, Forestales y Veterinarias de la UMSS, Casilla 3392, Cochabamba, Bolivia. Correo electrónico: j.gabriel@proinpa.org
3. Fundación para la Promoción e Investigación de Productos Andinos (PROINPA). Casilla 4285. Cochabamba. Bolivia.
4. Fundación para la Promoción e Investigación de Productos Andinos (PROINPA). Casilla 4285. Cochabamba. Bolivia.
5. Facultad de Ciencias Agrícolas, Pecuarias, Forestales y Veterinarias de la UMSS, Casilla 3392, Cochabamba, Bolivia.
6. Fundación para la Promoción e Investigación de Productos Andinos (PROINPA). Casilla 4285. Cochabamba. Bolivia.
Inheritance and genetic relationship associated with performance, maturity in tomato hybrids [Solanum lycopersicum L. (Mill.)]
Resumen
Durante 2010-2011 se evaluaron en invernadero, 30 híbridos de tomate, con el objetivo de determinar las relaciones genéticas entre los distintos caracteres afines al rendimiento, el número de días a la madurez y duración en anaquel. Los resultados mostraron que el híbrido 5 fue el mejor en textura, para grados brix los mejores híbridos fueron: 2, 29, 25, 16 y 10, para número de días a la madurez, el híbrido 12. El híbrido 8 para tamaño y ancho de fruto, para longitud de fruto el híbrido 4 y el híbrido 9 para rendimiento. En el pH hubo efectos adicionales y en la textura de fruto, los grados brix y el número de días a la madurez, efectos dominantes. En el rendimiento, tamaño, longitud y ancho de fruto hubo efectos aditivos y dominantes. La herencia fue mayor para grados brix, tamaño, longitud y ancho de fruto. El progenitor 70 es portador de genes para menor pérdida de textura, mayor ganancia de grados brix, mayor pérdida de pH, mayor tamaño y mayor rendimiento de fruto. El progenitor 71 fue el portador de genes con menor número de días a la madurez, mayor longitud y mayor ancho de fruto. La correlación mostró una relación moderada y negativa (r=-0,62), entre tipo de crecimiento de la planta y forma predominante de fruto; además hubo correlaciones moderadas y negativas (r=-0,51) entre forma predominante de fruto, color exterior del fruto maduro y número de lóculos (r=-0,41). Se identificaron 2 componentes principales: el primero relacionado con textura de fruto, grados brix, tamaño, longitud y ancho de fruto; el segundo componente identificó al pH, número de días a la madurez y rendimiento por planta. La correspondencia múltiple mostró que en los híbridos evaluados predominan frutos redondos con pericarpio y la parte externa de color rojo.
Palabras clave: Anaquel; genes efectos aditivos; efectos dominantes; heredabilidad.
Abstract
In the year 2010-2011, in the greenhouse, 30 tomato hybrids were evaluated to determine the genetic relationships between the different characters related to yield, number of days to maturity and shelf life. Results showed that hybrid 5 was the best for texture. For brix degrees, the best hybrids were: 2, 29, 25, 16 and 10. Hybrid 12 for number of days to maturity. Hybrid 8 for fruit size and width. For fruit length, hybrid 4; and hybrid 9 for yield. The pH is controlled by additive effects, and in fruit texture, degrees brix and the number of days to maturity, dominant effects are involved. In yield, size, length and width of fruit there were additive and dominant effects involved. Inheritance was higher for degrees brix, size, length and width of fruit. Progenitor 70 carries genes for less loss of texture, higher gain of degrees brix, greater loss of pH, larger size and greater fruit yield. Instead, parent 71 is the carrier of genes for fewer days to maturity, greater length and greater width of the fruit. Correlation analysis showed a moderate and negative (r=-0.62) and relationship between type of plant growth and predominant fruit shape. Also, moderate and negative correlations (r=-0.50) were observed between dominant form of fruit, external color of the ripe fruit and number of locules (r=-0.41). Characterization showed a wide variability in quantitative variables. Two main components, were identified: the first related to fruit texture, brix degrees, size, length and width of fruit; the second component identified pH, number of days to maturity and yield per plant. Multiple correspondence analyses indicated that in the hybrids evaluated predominate round fruits with pericarp and the outside colored red.
Keywords: Shelf; genes; additive effects; dominant effects; heritability.
Introducción
La identificación y selección de líneas parentales son necesarias en cualquier programa de hibridación para generar nuevos y potenciales genotipos valiosos para combinar y lograr los efectos de genes deseados en una línea homocigótica. En ese aspecto la información relativa de los distintos tipos de acción génica, la magnitud relativa de la variación genética y la combinación de las estimaciones de habilidad combinatoria son parámetros importantes y vitales para lograr la composición genética de los cultivos (Hannan et ál. 2007).
La información es importante y podría proveer de una estrategia esencial para los mejoradores de tomate en la selección de las mejores combinaciones parentales para continuar el proceso de mejoramiento. La explotación de la heterosis depende principalmente de la detección y selección de germoplasma que se podría producir por las mejores combinaciones para los más importantes caracteres agronómicos. Toda la variabilidad genética observada en el análisis de cada carácter se fraccionó en sus componentes, es decir, la aptitud combinatoria general (ACG) y la aptitud combinatoria específica (ACE) tal como lo define Sprague (1966) y los efectos recíprocos, descritos por Griffing (1956). Ellos afirmaron que los efectos de ACG fueron debido a un tipo de acción génica de tipo aditivo y los efectos de ACE se debieron a la acción génica no aditiva (dominante o epistático).
Muchos estudios de la aptitud combinatoria para los componentes de rendimiento fueron evaluados en muchas especies. Algunos encontraron la predominancia de la ACG y que podría ser más importantes que la ACE (Khan et ál. 1991, Yaqoob et ál. 1997), entre tanto otros sugirieron que la ACE es más importante (Ortiz y Golmirzaie 2004, Biswas et ál. 2005).
El presente estudio tuvo como objetivo determinar las relaciones genéticas entre los componentes asociados al rendimiento, el número de días a la madurez y duración en anaquel para 30 híbridos de tomate.
Materiales y Métodos
El experimento se implementó en un invernadero de la Fundación PROINPA en la zona de El Paso en Cochabamba, geográficamente ubicada a 17°21’00.98” de latitud sur y de 66°15’47.58” de longitud oeste, a una altitud de 2617 msnm. Esta zona tiene una temperatura media anual de 18°C, con 55% de HR y una precipitación media anual de 569,9 mm.
Se utilizaron 30 híbridos de tomate (Cuadro 1) obtenidos en la Fundación PROINPA de la cruza entre 21 líneas parciales con 3 ciclos de autofecundación, una línea silvestres de Solanum lycopersicum con resistencia al tospovirus o peste negra (TSWV): 70 (89R Sw-5/Sw-5) y una línea susceptible a tospovirus: 71 (895 Laching Sw-5) (Gabriel et ál. 2013).
Las 21 líneas parentales fueron seleccionadas de 74 accesiones de la colección de trabajo de la Fundación PROINPA que fueron las mismas evaluadas por sus atributos de rendimiento, resistencia a enfermedades y otros caracteres agronómicos de interés (Gabriel et ál. 2008). Las líneas parentales provinieron de diferentes empresas productoras de tomate como Reuter Seeds (España), Enza Zaden (Holanda), Sperli (Holanda), Numhems Zaden (Holanda), Seminis (Chile), NeikerTecnalia (España) y del Programa Nacional de Hortalizas del INIAF (Bolivia).
El suelo se preparó manualmente, con una remoción y desterronado de las platabandas; luego se realizó la aplicación de fertilizante nitrofoska en una cantidad de 2,5 kg por platabanda. Se hicieron platabandas niveladas de 1 x 28,50 x 0,15 m.
Antes de la siembra se realizó el lavado de las bandejas de plastoformo con agua y detergente, luego se realizó la desinfestación con una solución de agua y cloro (50 ml de Cl/1 l de agua) durante 5 a 10 min. Se preparó sustrato en una relación: 1:1:1 de cascarilla de arroz, tierra vegetal y arena fina que se esterilizó en una caldera a vapor durante 35 min a 90°C. Se llenó los hoyos de las bandejas compactándola para evitar la formación de bolsas de aire. Se realizó la siembra en un hoyo de 0,5 a 1 cm de profundidad y un diámetro de un lápiz donde se depositó una semilla por hoyo y 55 semillas por tratamiento, finalmente se cubrió con lama desinfestada y con riego a capacidad de campo.
El transplante se realizó en sistema de plantación tresbolillo, para lo que se cavaron hoyos a una profundidad de 0,15 m a una distancia de 0,46 m entre hoyos dentro de la hilera y una distancia de 0,60 m entre surcos.
El control de malezas se realizó manualmente. Se controlaron plagas y enfermedades. En las plantas de hábito de crecimiento determinado, la poda se realizó en 3 ramas principales con eliminación de las ramas restantes. Las hojas viejas y los brotes se dejaron para la formación de ramas nuevas debido a que las 3 ramas principales terminan en una inflorescencia. En las indeterminadas, la poda se realizó en 2 ramas principales y se eliminó las ramas restantes. En este caso las hojas viejas y los brotes se eliminaron para evitar la formación de otras ramas secundarias. El tutoraje se realizó después de la poda. En las determinadas se colocaran 3 tutores, un tutor por cada rama principal. En las indeterminadas también se instaló un tutor por rama principal.
Para el análisis de anaquel se recolectaron 18 frutos por híbrido. Los frutos fueron recolectados en el estado fenológico de 1/3 verde y 2/3 maduro (pintonas). Se implementaron 8 tratamientos para evaluar la duración en anaquel y composición química de los frutos: 0, 3, 6, 9, 13, 16 y 21 días en anaquel. Las variables evaluadas fueron: el color, el peso, la textura, los grados brix y el pH.
Variables de respuesta
Las variables de respuesta de las características morfológicas, agronómicas y agroindustriales fueron evaluadas según las recomendaciones del IPGRI (2008). Las variables cuantitativas fueron: tamaño de fruto (TmF), longitud de fruto (LnF), ancho de fruto (AnF) y rendimiento (Y). Las variables agronómicas fueron: número de días a la floración (NoDM). Las variables para duración en anaquel fueron: pérdida de textura (PTXT), pérdida de pH (PpH) y grados brix (G°Bx). Las variables cualitativas fueron: tipo de crecimiento de la planta (TCP), forma predominante del fruto (FPF), color exterior del fruto maduro (CFM), color de la carne del pericarpio (CCP), forma del corte transversal del fruto (CTF) y número de lóculos (NOL).
Diseño experimental y análisis estadístico
El estudio de durabilidad en anaquel se implementó por medio de un diseño completamente aleatorio.
El estudio de las características morfológicas, agronómicas y de resistencia a estreses bióticos y abióticos se implementó en diseño de bloques completos al azar con submuestreo con 4 repeticiones. Los tratamientos fueron los híbridos de tomate, que fueron distribuidos en 4 bloques con 10 plantas por híbrido por unidad experimental y 3 plantas para el submuestreo.
Sobre la base del modelo definido, se realizaron análisis de varianza para probar hipótesis acerca de los efectos fijos y comparaciones de medias de los híbridos. El análisis de varianza también sirvió para estimar los componentes de varianza para los efectos aleatorios. Los análisis indicados se realizaron por medio del Proc GLM del SAS (SAS 2004).
Para el análisis genético de los híbridos se utilizó el Diseño I de Carolina de Norte, en el que cada progenitor masculino se apareó con m hembras (Martínez 1988). El análisis de este diseño permitió estimar las varianzas genéticas de las variables de respuesta y determinar la aptitud combinatoria general (ACG) del macho. También se estimó la herencia en sentido estrecho (h2) y amplio (H2).
Finalmente se realizaron análisis entre correlación entre las variables, así como un análisis de componentes principales para las variables cuantitativas y un análisis de correspondencia múltiple para las variables cualitativas (López e Hidalgo 2003).
Resultados y Discusión
Análisis de varianza en anaquel
En el análisis de varianza, (Cuadro 2) se encontraron diferencias significativas (p<0,05) entre híbridos para las variables pérdida de textura de fruto y ganancia en grados brix; esto indica que la respuesta de al menos uno de los híbridos fue diferente para cada variable evaluada. En cambio, la variable pH no presentó ninguna diferencia significativa (p<0,05), esto indica que no existieron diferencias en el pH en ninguno de los híbridos. Por otro lado la comparación de medias al p<0,01 de probabilidad, para la variable PTxT, mostró que el híbrido con menor pérdida de textura o mayor firmeza de fruto fue la cruza 5. Por otra parte, los híbridos con mayor ganancia en grados Brix fueron los híbridos 21, 29, 25, 16 y 10. En cambio, para la variable pH no existieron diferencias significativas entre híbridos.
Componentes de varianza genética en anaquel
En la estimación de varianzas genéticas estimados (Cuadro 3) para pérdida en pH mostró que la varianza aditiva
es mayor a cero. Esto indicaría que la pérdida en pH estaría gobernada por efectos aditivos. En cambio, para pérdida de TxT y ganancia en grados brix la varianza de dominancia , fue mayor a cero. Esto significaría que la pérdida en TxT y ganancia en grados brix estarían gobernados por efectos de dominancia. Para las variables pérdida en textura de fruto y ganancia en grados brix, los efectos de dominancia fueron un número infinito de veces más importantes que los efectos aditivos. Por otra parte, en el caso de la pérdida en pH los efectos aditivos fueron más importantes respecto a los efectos de dominancia. Resultados similares fueron hallados por Gragera (2003), quien encontró que el pH, tuvo una mayor expresión de genes de efectos aditivos (d/a=0,03) durante un estudio de herencia en tomates de industria de origen español. Asimismo, en los mismos genotipos encontró que los efectos de dominancia (d/a=2,76) fueron más importantes para grados brix. Esto significaría que hay genes mayores y menores involucrados en la herencia de estos caracteres. Por otro lado, para TxT resultados similares fueron hallados por Pereira da Costa et ál. (2009), quienes reportaron que en accesiones de S. lycopersicum, existe una mayor expresión de genes con efectos de dominancia, donde los efectos de dominancia fueron 1,33 veces más importantes que los efectos aditivos.
La estimación de la heredabilidad a partir de los efectos genéticos (Cuadro 3), mostró una heredabilidad en sentido amplio alta para ganancia en grados brix, lo que sugiriere que existe una amplia variación genética y pobre variación ambiental para este carácter. En cambio existe una baja heredabilidad en sentido amplio para pérdida en textura. Esto propone que este carácter, estuvo ampliamente influenciado por el ambiente. Por otra parte, la heredabilidad en sentido estrecho para estas 2 variables fue igual a cero; dicho aspecto confirma que estos caracteres estarían gobernados solo por efectos de dominancia. Resultados similares fueron reportados en tomates mexicanos evaluados por Hernández (2013). En dicho experimento hubo una alta heredabilidad en sentido amplio (H2>0,5) y una baja heredabilidad en sentido estrecho (h2<0,5) para grados brix. Por otra parte, Moreno et ál. (2002), encontraron una heredabilidad en sentido estrecho alta (h2>0,45) para grados brix, que indica que los efectos aditivos son de importancia en esta variable. Esto sugeriría que este carácter estaría gobernado tanto por efectos aditivos como de dominancia. Pereira da Costa et ál. (2009) también encontraron una heredabilidad en sentido estrecho similar a la del presente trabajo (h2<0,1) para textura de fruto. En cambio la heredabilidad en sentido amplio fue moderada (H2=0,39), lo que confirma los efectos de dominancia más importantes. Para la variable pH, las heredabilidades en sentido amplio y estrecho fueron bajas e idénticas, aspecto que señala la no existencia de efectos de dominancia para esta variable (tal como se observó en la importancia de los efectos genéticos) y que está muy influenciada por el ambiente. En cambio, Moreno et ál. (2002) encontraron una heredabilidad en sentido estrecho alta (h2>0,45), que indica una baja influencia del medio ambiente. De todos modos, se confirma la importancia de los efectos aditivos para esta variable.
ACG de los machos para la prueba en anaquel
La estimación de la ACG de los machos (Cuadro 4), mostró variabilidad en el comportamiento de los progenitores para las diferentes variables de respuesta en anaquel. El progenitor 70 mostró la menor ACG para pérdida en textura de fruto. Asimismo, tuvo las mayores ACG para ganancia en grados brix y para pérdida en pH, que sugiriere a este progenitor como el portador de genes para menor pérdida de firmeza de fruto, ganancia en grados brix y mayor pérdida en pH del jugo de tomate en anaquel durante 21 días en el mismo ambiente. Estudio realizado por Lagos (2012), en la evaluación de material de tomate procedente de Colombia y Ecuador, reportó que las varianzas de los efectos de ACG fueron relativamente bajas (menores a uno) en pH, sólidos solubles totales (grados brix) y textura de fruto, indica de que esto se debe a la uniformidad genética de los parentales en estas variables. En resumen, el parental 70 aporta genes para menor pérdida de textura de fruto, mayor contenido de sólidos solubles (grados brix) y mayor pérdida de pH de fruto. Por tanto, este progenitor es una buena fuente para desarrollar futuros programas de mejoramiento sobre la vida en anaquel de frutos de tomate.
Análisis de varianza morfológico y agronómico
El análisis de varianza para híbridos (Cuadro 5) mostró diferencias altamente significativas (p<0,01) para las variables número de días a la madurez, longitud de fruto, ancho de fruto, tamaño de fruto y rendimiento; esto significa que el comportamiento de al menos uno de los híbridos evaluados es diferente en cada una de las variables. La comparación de medias (p<0,01) mostró que el híbrido 12 alcanzó la menor cantidad de días a la madurez, el híbrido 8 obtuvo el mayor tamaño de fruto, el híbrido 4 exhibió la mayor longitud de fruto, los híbridos 8,17, 6 y 14 obtuvieron el mayor ancho de fruto y 9 fue el híbrido con mayor rendimiento de fruto.
Componentes de varianza genética para variables morfológicas y agronómicas
Los componentes de varianza genética estimados (Cuadro 6), para número de días a la madurez muestran que la varianza de dominancia
es mayor a cero. Esto indica que este carácter está gobernado solo por efectos de dominancia como lo indica la importancia de los efectos genéticos (γ), donde los efectos de dominancia son un infinito número de veces más importantes que los efectos aditivos. En cambio, para tamaño de fruto, longitud de fruto, ancho de fruto y rendimiento las varianzas aditiva y de dominancia son mayores a cero, lo cual indica que tanto efectos de dominancia como aditivos están involucrados en herencia de las 4 variables. Sin embargo, al considerar la importancia de los efectos genéticos (γ) para tamaño de fruto, los efectos de dominancia son 5,4 veces más importantes respecto a los efectos aditivos, para longitud de fruto, los efectos de dominancia son 5,6 veces más importantes respecto a los efectos aditivos, para el ancho de fruto, los efectos de dominancia fueron 1,39 veces más importantes que los efectos aditivos y finalmente para el rendimiento, los efectos de dominancia fueron 1,81 veces más importantes respecto a los efectos aditivos. Resultados similares fueron hallados por Hernández (2013), en genotipos mexicanos, quien reportó una mayor expresión de genes con efectos de dominancia parcial para los caracteres precocidad, longitud, ancho, y tamaño de fruto. Así mismo, un efecto similar fue observado por Grandillo y Tanksley (1996), en el tamaño de fruto y por Burdick (1954), en la precocidad. Sin embargo, dicha dominancia suele ser relativa en variables de precocidad debido a que su expresión depende de la etapa fisiológica y condiciones ambientales (Burdick 1954). Por otra parte, para rendimiento se encontraron efectos (Meneses 2002), donde la varianza de dominancia para peso de fruto, presentó valores muy bajos y/o negativos, lo que permite considerar que la varianza genética total estuvo determinada por efectos aditivos de los genes, lo que es muy favorable y factible de explotarse a través de cualquier esquema de selección. Estos resultados demuestran que la precocidad, tamaño, longitud, ancho y rendimiento de fruto, estarían controlados principalmente por genes con efectos de dominancia. La estimación de la heredabilidad a partir de los efectos genéticos (Cuadro 6), mostró valores altos para la heredabilidad en sentido amplio (H2) para los variables tamaño, longitud y ancho de fruto, lo que sugiriere la presencia de alta variabilidad genética e insignificante variación ambiental. En cambio, las heredabilidades en sentido estrecho fueron bajas para todas las variables en general, lo cual indica que los efectos aditivos son de poca importancia, y plantea que la selección de híbridos de tomate sería más efectiva a través del tamaño, longitud y ancho de fruto. Los valores altos para la heredabilidad en sentido amplio y bajos para heredabilidad en sentido estrecho de estas 2 variables confirman que los efectos de dominancia son más importantes que los efectos aditivos. Estudios en tomates mexicanos realizados por Gaspar et ál. (2012) coinciden con los resultados encontrados en el presente trabajo, que identifica la heredabilidad en sentido amplio, como alta (H2>0,5) para longitud de fruto y baja para peso de fruto. Por otro lado, para ancho de fruto valores semejantes de heredabilidad (H2=0,66) en sentido amplio fueron estimados por (Hernández 2013). En general, la investigación realizada presenta las heredabilidades en sentido estrecho como bajas para las 5 variables (Cuadro 6). Resultados similares fueron encontrados por Pérez et ál. (2010), en material de Cuba y Florida, donde encontraron heredabilidades en sentido estrecho bajas para las variables longitud, ancho y tamaño de fruto, esto se debe a que la componente de varianza aditiva tiene un valor bajo, por lo que la transferencia genética es baja y la determinación de este carácter está influida por las condiciones ambientales en que se desarrollan los individuos. Así mismo, para la variable número de días a la madurez la heredabilidad en sentido amplio y estrecho concuerdan con los resultados encontrados en accesiones de Solanum lycopersicum en material de origen Argentino (Pereira da Costa et ál. 2009), quiénes obtuvieron una heredabilidad en sentido amplio intermedia (H2=0,36) y una heredabilidad estrecha nula (h2=0) para número de días a la madurez.
ACG para machos de variables morfológicas y agronómicas
Las ACGs estimadas para los 2 progenitores machos (Cuadro 7), mostró una amplia diferencia en el comportamiento de los progenitores en las diferentes variables de respuesta. El progenitor 71 mostró la menor ACG para número de días a la madurez, que sugiriere al progenitor como el portador de genes para menor número de días a la madurez. En cambio, el progenitor 70 mostró las mayores ACG’s para el tamaño de fruto y peso de fruto, lo que propone a este progenitor como el portador de genes para mayor tamaño y rendimiento de fruto. Por otra parte, el progenitor 71 mostró las mayores ACGs para longitud y ancho de fruto; por tanto, este progenitor sería el portador de genes para mayor longitud y ancho de fruto. Los valores de mejora de ACG con el parental 70 para Y y TmF son 0,114 y 0,005; por tanto, este progenitor contribuyó en general con 114 gramos al rendimiento por planta y en 0,13 mm al tamaño de los híbridos. En cambio, con el parental 71 los valores de mejora son 0,555 y 0,484 para LnF y AnF, lo cual significa que este progenitor contribuyó con 555 y 484 micrómetros a la longitud y ancho de fruto de los híbridos respectivamente. Asimismo, este parental contribuyó con la reducción de 1,8 días para NoDM. Resultados similares, fueron encontrados por (Sahagún et ál. 1999), en Physalis ixocarpa Brot. Donde hubo diferencias significativas (p≤0,05) en las ACGs de 3 genotipos para precocidad expresada a través de rendimiento en la primera cosecha, con el genotipo Salamanca que aportó mayor peso en la primera cosecha.
Análisis de correlación de variables cuantitativas y cualitativas
En el análisis de variables cuantitativas en anaquel no hubo ninguna correlación significativa al pr<0,01 de probabilidad entre las variables pérdida en textura de fruto, ganancia en grados brix y pérdida en pH. Murillo et ál. (2004), mencionan que ante la ausencia de correlación se debe realizar un análisis multivariado de Factores Principales de variación, que permite reducir la dimensionalidad de las variables, permite explorar la acción conjunta de las variables y determinar el efecto de cada variable en presencia de las otras (Uriel y Aldás 2005).
En el análisis de caracteres cualitativos agromorfológicos hubo una correlación significativa entre las variables TCP y FPF. Esta correlación fue moderada y negativa (r=-0,62). Esto indicó que la forma predominante del fruto depende del tipo de crecimiento de la planta. En las indeterminadas predominan los frutos redondos y redondos achatados (92%) y el 5% fueron redondos alargados. En cambio, en las determinadas el porcentaje de frutos redondos alargados fue mayor (30%) que en las indeterminadas y el porcentaje de frutos redondos y redondos achatados fue inferior que en las indeterminadas (65%). Asimismo, se presentaron frutos cordiformes (tipo pera) en un 5%. Resultados similares fueron hallados por Florido et ál. (2008), cuando cruzaron genotipos indeterminados de frutos redondos achatados con genotipos determinados de frutos redondos alargados. En las progenies indeterminadas se encontró un 85% de frutos redondos achatados y un 15% frutos redondos alargados. Por otra parte, en las progenies determinadas se encontró frutos redondos alargados en un 40% y frutos redondos achatados en un 60%. Por otra parte, la variable FPF se correlacionó negativa y moderadamente con la variable CFM (r=-0,51). Esto significaría que entre los frutos alargados predomina el color naranja y entre los frutos redondos los colores rojo y rosado. También esta variable FPF se correlacionó moderada y negativamente con la variable NOL (r=-0,41). Esto indicó que en los frutos alargados predomina la presencia de 3 lóculos por fruto y que en los frutos redondos predomina la presencia de 4 lóculos en la mayoría de los híbridos. Tales resultados concuerdan con los reportados por Medina y Lobo (2001), quiénes encontraron de 2 a 3 lóculos/fruto en frutos alargados y de 4 a 5 lóculos/fruto en frutos redondeados en la mayoría de las introducciones evaluadas.
En los caracteres cuantitativos agromorfológicos se encontró una correlación moderada y positiva se dio entre las variables LnF con AnF (r=0,67), esto significaría que a mayor longitud de fruto sería más ancho el fruto. Por otra parte, TmF estuvo moderadamente y positivamente correlacionada con LnF (r=0,56) y AnF (r=0,55). Por tanto, entre los híbridos evaluados, los híbridos con mayor tamaño de fruto exhibieron mayor ancho y longitud de fruto. Por otro lado hubo una correlación positiva baja entre las variables TmF y Y (r=0,35). Esto permite deducir que a mayor tamaño de fruto existe mayor rendimiento por planta. Hernández (2013), también encontró una correlación moderada y positiva (r=0,40) entre tamaño de fruto y rendimiento. Así mismo, Lippman y Tanksley (2001), encontraron una correlación positiva entre tamaño y rendimiento de fruto en poblaciones derivadas de S. pimpinellifolium y genotipos de S. lycopersicum de fruto grande. En nuestro caso la selección para mayor rendimiento y tamaño de fruto se podría realizar con base en híbridos de mayor longitud y ancho de fruto.
Análisis de componentes principales (ACP)
Una vez generados los componentes, se utilizaron los criterios propuestos por Pla (1986), Se retuvo 2 componentes cuyo valor propio fue ≥1 y que expresaron más del 70% de la varianza total (López e Hidalgo 2003). En el análisis de anaquel el primer componente contribuyó con el 41,19% de la varianza total. Aportó en forma positiva las variables PTxT y G°Bx; esto denotó que dicho componente identificó a los híbridos con mayor pérdida en textura y ganancia en contenido de grados brix o azúcar en los frutos. Resultados similares fueron hallados por Bonilla (2013), donde también encontró que la textura o firmeza de fruto fue una de las variables que más contribuyó a este componente. Por otro lado, la contribución de los grados brix a este componente concuerda con los estudios realizados por Salgado (2010), con material de México, quien encontró que el componente uno explicaba más del 46% de la variabilidad de las características evaluadas donde el porcentaje de los sólidos solubles fue muy importante para este componente. El segundo componente contribuyó con el 34,28% de la varianza total, en el cual la variable pH aportó positivamente. Por lo que este componente distinguió a los híbridos con mayor pérdida en contenido de pH.
En el análisis morfológico el primer componente contribuyó con el 52,35% del total de la varianza y aportó en forma positiva las variables TmF, LnF y AnF. Esto denotó que este componente identificó a los híbridos con mayor tamaño de fruto, longitud de fruto y ancho de fruto. Bonilla (2013), en materiales de los estados de Puebla y Oaxaca, encontró que el tamaño de fruto, longitud de fruto y ancho de fruto fueron las variables que más contribuyeron a este componente. Por otro lado, el segundo componente contribuyó con el 17,53% de la varianza total, en el cual la variable Y aportó positivamente. Por otra parte, la variable NoDM, contribuyó en forma negativa. Por lo que este componente distinguió a los híbridos con mayor rendimiento y precocidad.
Análisis de correspondencia múltiple
El análisis de correspondencia múltiple mostró variabilidad genética dentro de los 30 híbridos de tomate. Aunque la mayoría de los híbridos se agruparon por características similares, existieron casos aislados como el híbrido 20 que no se agrupo con ninguno de los híbridos. Este híbrido presenta frutos con color exterior rosado y carne del pericarpio del mismo color. Por otro lado, los híbridos 2, 17 y 23 se agruparon al presentar frutos redondos alargados con un color exterior naranja y la carne del pericarpio del mismo color. Otros híbridos que se diferenciaron fueron los híbridos 3, 5, 18, 27 y 28 al presentar forma redonda y un color exterior rojo y la carne del pericarpio también del mismo color. En general se observó que hubo una correlación entre la forma predominante del fruto, el color exterior y el color del pericarpio.
Conclusiones
En las variables de anaquel se encontró que la pérdida en textura de fruto y ganancia en grados brix están gobernados solo por efectos de dominancia. La pérdida en pH está gobernada solo por efectos aditivos. En las variables morfológicas se observó que el número de días a la madurez está gobernado solo por efectos de dominancia. En cambio, el tamaño de fruto, longitud de fruto, ancho de fruto y rendimiento de fruto están gobernados tanto por efectos aditivos y de dominancia.
Las heredabilidades en sentido estrecho para las 3 variables de anaquel fueron bajas. La heredabilidad en sentido amplio fue alta para ganancia en grados brix, moderada para pérdida en textura y baja para pérdida en pH. Las heredabilidades en sentido estrecho fueron bajas para las 5 variables morfológicas y agronómicas. Las heredabilidades en sentido amplio fueron bajas para número de días a la madurez y rendimiento y altas para tamaño, longitud y ancho de fruto.
El progenitor 70 fue portador de genes para menor pérdida de textura de fruto, mayor ganancia en grados brix y mayor pérdida en pH, mayor tamaño de fruto y mayor rendimiento de fruto.planta-1. En cambio, el progenitor 71 fue portador de genes para menor número de días a la madurez, mayor longitud y ancho de fruto.
El análisis de correlación entre pérdida en textura de fruto, ganancia en grados brix y pérdida en pH no mostró ninguna correlación significativa entre las 3 variables. Entre las variables morfológicas se observó una moderada correlación entre la forma predominante del fruto maduro con las variables: tipo de crecimiento de la planta, color exterior de fruto maduro y número de lóculos del fruto. Asimismo, el rendimiento esta moderadamente correlacionada con las variables tamaño, longitud y ancho de fruto. Las 2 últimas variables están altamente correlacionadas entre sí.
El análisis de componentes principales de las variables en anaquel, permitió generar 2 nuevos componentes. El primer componente permitió distinguir a los híbridos con menor pérdida de textura de frutos y mayor contenido de grados brix. El segundo componente identificó a los híbridos con mayor pérdida pH. Por otra parte, el análisis de componentes principales de las variables agromorfológicas, generó 2 nuevos componentes. El primer componente distinguió a los híbridos con mayor tamaño, longitud y ancho de fruto. El segundo componente identificó a los híbridos con mayor precocidad y rendimiento de fruto.
El análisis de correspondencia múltiple permitió distinguir que los híbridos de tomate evaluados a la madurez, se relacionan con la forma predominante de los frutos, con el color exterior y el color del pericarpio.
Agradecimientos
Se agradece al apoyo económico del proyecto “Fortaleciendo capacidades de innovación participativa para luchar contra la pobreza rural” (IP – Holanda), para la realización de esta investigación.
Literatura Citada
BISWAS M.K., MONDAL M.A.A., HOSSAIN M., ISLAM R. 2005. Selection of suitable parents in the development of potato hybrids in Bangladesh. Chinese Potato Journal 19:193-197. [ Links ]
BONILLA O. 2013. Diversidad agronómica y morfológica de jitomates ariñonados y tipo pimento de alto valor comercial en los pueblos de Puebla y Oaxaca. Tesis de maestría, Colegio de Posgraduados, Montecillo, México. 77 p. [ Links ]
BURDICK B.A. 1954. Genetics of heterosis for earliness in the tomato. Genetics 39:488-505. [ Links ]
FLORIDO M., ALVAREZ M., LARA R., PLANA D., VARELA M., SHAGARODSKY T., MOYA C. 2008. Análisis de la variabilidad morfoagronómica en la colección de tomate (Solanum L. sección Lycopersicon subsección Lycopersicon) conservada ex situ en Cuba. Cultivos Tropicales 29(2):43-48. [ Links ]
GABRIEL J., MAGNE J., CRESPO M., PLATA G., ANGULO A., GARCÍA R. 2008. Caracterización agronómica, morfológica y de resistencia a virus en 74 colecciones de trabajo del banco de germoplasma de tomate (Lycopersicum sculentum), pp. 1-10. In: Informe anual de Proyectos gestión 2007–2008, Fundación PROINPA, Cochabamba, Bolivia. [ Links ]
GABRIEL J., SANABRIA D., VERAMENDI S., PLATA G., ANGULO A., CRESPO M. 2013. Resistencia genética de híbridos de tomate [Solanum lycopersicum L. (Mill.)] al virus del bronceado (TSWV). Agronomía Costarricense 37(1):61-69. [ Links ]
GASPAR P., CARRILLO J.C., CHAVEZ J.L., VERA A.M., PÉREZ I. 2012. Variación de caracteres agronómicos y licopeno en líneas avanzadas de tomate (Solanum lycopersicum L.). Oaxaca, México. 22 p. [ Links ]
GRAGERA J. 2003. La mejora genética de tomate de industria, en el servicio de investigación y desarrollo tecnológico de la junta de Extremadura. Guadajira, Valencia, España. 43 p. [ Links ]
GRANDILLO S., KU H., TANKSLEY S.D. 1996. Characterization offs8. La major QTL influencing fruit shape in tomato. Molecular Breeding 2:251-260. GRIFFING B. 1956. Concept of general and specific combining ability in relation to diallel crossing system. Australian Journal of Biology Science 9:463-493. [ Links ]
HANNAN M.M., BISWAS M.K., AHMED M.B., HOSSAIN M., ISLAM R. 2007. Combining Ability Analysis of Yield and Yield Components in Tomato (Lycopersicum esculentum Mill.). Turkish Journal Botany 31:550-563. [ Links ]
HERNÁNDEZ A. 2013. QTLs y heterosis en jitomate en una poblaciòn F2 derivada de uma cruza interespecífica. Tesis de maestría, Colegio de Posgraduados, Montecillo, México. 65 p. [ Links ]
IPGRI. 2008. Descriptores para el tomate (Lycopersicon spp.). Bogota, Colombia. 49 p. [ Links ]
KHAN M.A., CHEEMA K.L., MASOOD A., SADAQAT H.A. 1991. Combining ability in cotton (Gossypium hirsutum L.). Journal Agriculture Research 29:311318. [ Links ]
LAGOS L.K. 2012. Evaluacion del potencial genético de algunos parentales de tomate de árbol (Cyphomandra betacea cav. Sendt.) apartir de un dialelico parcial circulante, tesis de grado, Universidad Nacional de Colombia –Facultad de Ciencias Agropecuarias. Palmira, Colombia. 79 p. [ Links ]
LIPPMAN Z.S., TANKSLEY D. 2001. Dissenting the genetic pathway to extreme fruit size in tomato using a cross between the small-fruited wild species Lycopersicum pimpinellifolium and L. esculentum var. Giant Heirloom Genetics 158:413-422. [ Links ]
LÓPEZ F.T., HIDALGO R. 2003. Análisis Estadístico de datos de Caracterización morfológica de recursos genéticos. Instituto Internacional de recursos fitogenéticos Cali – Colombia. 89 p. [ Links ]
MARTÍNEZ A. 1988. Diseños experimentales: Métodos y elementos de teoría. Editorial Trillas, México, DF. México. 756 pp. [ Links ]
MEDINA C., LOBO M. 2001. Variabilidad morfológica en el tomate pajarito (Lycopersicum esculentum var. Cerasiforme) precursor del tomate cultivado. Revista Corpoica 3(2):39-50. [ Links ]
MENESES I., VILLANUEVA C., SAHAGUM J., ROQUE VASQUEZ T., MERRICK L.C. 2002. Componentes de varianza genética y respuesta a la selección combinada en la calabaza (Cucurbita pepo L.) bajo el sistema milpa. Universidad Autónoma de Chapingo, Chapingo, México. 14 p. [ Links ]
MORENO M., PEÑA A., SAHAGUN J., RODRÍGUEZ J.,M ORA R. 2002. Varianza aditiva, heredabilidad y correlaciones en La variedad M1-Fitotecnia de tomate de cáscara (Physais ixocarpa Brot.). Revista Fitotecnia 25(3):20-29. [ Links ]
MURILLO M., BORREGO F., RODRÍGUEZ S., RAMOS F. 2004. Caracterización del potencial bioquímico y nutritivo de genotipos sobresalientes de tomate (Lycopersicum esculentum Mill.). Tesis de maestría, Universidad Autónoma Agraria Antonio Narro. Buenavista, México. 222 p. [ Links ]
ORTIZ R., GOLMIRZAIE A.M. 2004. Combining ability analysis and correlation between breeding values in true potato seed. Plant Breeding 123:564-567. [ Links ]
PEREIRA DA COSTA J., MARTÍNEZ V., RODRÍGUEZ G., PRATTA G., ZARZOLI R. 2009. Influencia de genes exóticos sobre la firmeza de fruto y precocidad del fruto de tomate. U.N. de Rosario. Argentina. 18 p. [ Links ]
PÉREZ L., SIGARROA A., BANDERA E., RODRÍGUEZ N., VALDEZ J. 2010. Estimación de la heredabilidad en sentido ancho a caracteres de importancia agrícola evaluadas en una población de guayabo. Tesis maestría, Universidad de la Habana, Habana, Cuba. 20 p. [ Links ]
PLA L. 1986. Análisis multivariado: método de componentes principales. OEA, Washington, EE.UU. pp. 3-15. [ Links ]
SAS Institute Inc. 2004. SAS/STAT Users Guide, Version 9.2, Fourth Edition, Vol. 2, SAS Institute Inc., Cary, N.C. [ Links ]
SAHAGUN J., GÓMEZ F., PEÑA A. 1999. Efectos de aptitud combinatoria en poblaciones de tomate de cascara (Physalis ixocarpa Brot.), Universidad Autónoma Chapingo. Chapingo, México. 23 p. [ Links ]
SALGADO M.L., RAMÍREZ P., UTRERA Q. 2010. Contenido de licopeno en acervos de jitomate mexicano, pp. 35-38. In: R. Díaz, J.F Álvarez y A Huerta. (eds.). Memorias del foro Regional de Agricultura Sostenible. Sociedad Mexicana de Agricultura Sostenible A. C. Noviembre. Colegio de Postgraduados, Campus Puebla, México. [ Links ]
SPRAGUE G.F. 1966. Plant Breeding. Ames, IA, USA: Iowa State University Press. [ Links ]
URIEL E., ALDAS J. 2005. Análisis multivariante aplicado. Thomson, Madrid, España. 523 p. [ Links ]
YAQOOB M., HASSAN G., MAHMOOD G., SHAH N.H. 1997. Combining ability studies for some quality traits in cotton (Gossypium hirsutum L.). J. Pure Appl Sci. 16:47-50. [ Links ]
BONILLA O. 2013. Diversidad agronómica y morfológica de jitomates ariñonados y tipo pimento de alto valor comercial en los pueblos de Puebla y Oaxaca. Tesis de maestría, Colegio de Posgraduados, Montecillo, México. 77 p. [ Links ]
BURDICK B.A. 1954. Genetics of heterosis for earliness in the tomato. Genetics 39:488-505. [ Links ]
FLORIDO M., ALVAREZ M., LARA R., PLANA D., VARELA M., SHAGARODSKY T., MOYA C. 2008. Análisis de la variabilidad morfoagronómica en la colección de tomate (Solanum L. sección Lycopersicon subsección Lycopersicon) conservada ex situ en Cuba. Cultivos Tropicales 29(2):43-48. [ Links ]
GABRIEL J., MAGNE J., CRESPO M., PLATA G., ANGULO A., GARCÍA R. 2008. Caracterización agronómica, morfológica y de resistencia a virus en 74 colecciones de trabajo del banco de germoplasma de tomate (Lycopersicum sculentum), pp. 1-10. In: Informe anual de Proyectos gestión 2007–2008, Fundación PROINPA, Cochabamba, Bolivia. [ Links ]
GABRIEL J., SANABRIA D., VERAMENDI S., PLATA G., ANGULO A., CRESPO M. 2013. Resistencia genética de híbridos de tomate [Solanum lycopersicum L. (Mill.)] al virus del bronceado (TSWV). Agronomía Costarricense 37(1):61-69. [ Links ]
GASPAR P., CARRILLO J.C., CHAVEZ J.L., VERA A.M., PÉREZ I. 2012. Variación de caracteres agronómicos y licopeno en líneas avanzadas de tomate (Solanum lycopersicum L.). Oaxaca, México. 22 p. [ Links ]
GRAGERA J. 2003. La mejora genética de tomate de industria, en el servicio de investigación y desarrollo tecnológico de la junta de Extremadura. Guadajira, Valencia, España. 43 p. [ Links ]
GRANDILLO S., KU H., TANKSLEY S.D. 1996. Characterization offs8. La major QTL influencing fruit shape in tomato. Molecular Breeding 2:251-260. GRIFFING B. 1956. Concept of general and specific combining ability in relation to diallel crossing system. Australian Journal of Biology Science 9:463-493. [ Links ]
HANNAN M.M., BISWAS M.K., AHMED M.B., HOSSAIN M., ISLAM R. 2007. Combining Ability Analysis of Yield and Yield Components in Tomato (Lycopersicum esculentum Mill.). Turkish Journal Botany 31:550-563. [ Links ]
HERNÁNDEZ A. 2013. QTLs y heterosis en jitomate en una poblaciòn F2 derivada de uma cruza interespecífica. Tesis de maestría, Colegio de Posgraduados, Montecillo, México. 65 p. [ Links ]
IPGRI. 2008. Descriptores para el tomate (Lycopersicon spp.). Bogota, Colombia. 49 p. [ Links ]
KHAN M.A., CHEEMA K.L., MASOOD A., SADAQAT H.A. 1991. Combining ability in cotton (Gossypium hirsutum L.). Journal Agriculture Research 29:311318. [ Links ]
LAGOS L.K. 2012. Evaluacion del potencial genético de algunos parentales de tomate de árbol (Cyphomandra betacea cav. Sendt.) apartir de un dialelico parcial circulante, tesis de grado, Universidad Nacional de Colombia –Facultad de Ciencias Agropecuarias. Palmira, Colombia. 79 p. [ Links ]
LIPPMAN Z.S., TANKSLEY D. 2001. Dissenting the genetic pathway to extreme fruit size in tomato using a cross between the small-fruited wild species Lycopersicum pimpinellifolium and L. esculentum var. Giant Heirloom Genetics 158:413-422. [ Links ]
LÓPEZ F.T., HIDALGO R. 2003. Análisis Estadístico de datos de Caracterización morfológica de recursos genéticos. Instituto Internacional de recursos fitogenéticos Cali – Colombia. 89 p. [ Links ]
MARTÍNEZ A. 1988. Diseños experimentales: Métodos y elementos de teoría. Editorial Trillas, México, DF. México. 756 pp. [ Links ]
MEDINA C., LOBO M. 2001. Variabilidad morfológica en el tomate pajarito (Lycopersicum esculentum var. Cerasiforme) precursor del tomate cultivado. Revista Corpoica 3(2):39-50. [ Links ]
MENESES I., VILLANUEVA C., SAHAGUM J., ROQUE VASQUEZ T., MERRICK L.C. 2002. Componentes de varianza genética y respuesta a la selección combinada en la calabaza (Cucurbita pepo L.) bajo el sistema milpa. Universidad Autónoma de Chapingo, Chapingo, México. 14 p. [ Links ]
MORENO M., PEÑA A., SAHAGUN J., RODRÍGUEZ J.,M ORA R. 2002. Varianza aditiva, heredabilidad y correlaciones en La variedad M1-Fitotecnia de tomate de cáscara (Physais ixocarpa Brot.). Revista Fitotecnia 25(3):20-29. [ Links ]
MURILLO M., BORREGO F., RODRÍGUEZ S., RAMOS F. 2004. Caracterización del potencial bioquímico y nutritivo de genotipos sobresalientes de tomate (Lycopersicum esculentum Mill.). Tesis de maestría, Universidad Autónoma Agraria Antonio Narro. Buenavista, México. 222 p. [ Links ]
ORTIZ R., GOLMIRZAIE A.M. 2004. Combining ability analysis and correlation between breeding values in true potato seed. Plant Breeding 123:564-567. [ Links ]
PEREIRA DA COSTA J., MARTÍNEZ V., RODRÍGUEZ G., PRATTA G., ZARZOLI R. 2009. Influencia de genes exóticos sobre la firmeza de fruto y precocidad del fruto de tomate. U.N. de Rosario. Argentina. 18 p. [ Links ]
PÉREZ L., SIGARROA A., BANDERA E., RODRÍGUEZ N., VALDEZ J. 2010. Estimación de la heredabilidad en sentido ancho a caracteres de importancia agrícola evaluadas en una población de guayabo. Tesis maestría, Universidad de la Habana, Habana, Cuba. 20 p. [ Links ]
PLA L. 1986. Análisis multivariado: método de componentes principales. OEA, Washington, EE.UU. pp. 3-15. [ Links ]
SAS Institute Inc. 2004. SAS/STAT Users Guide, Version 9.2, Fourth Edition, Vol. 2, SAS Institute Inc., Cary, N.C. [ Links ]
SAHAGUN J., GÓMEZ F., PEÑA A. 1999. Efectos de aptitud combinatoria en poblaciones de tomate de cascara (Physalis ixocarpa Brot.), Universidad Autónoma Chapingo. Chapingo, México. 23 p. [ Links ]
SALGADO M.L., RAMÍREZ P., UTRERA Q. 2010. Contenido de licopeno en acervos de jitomate mexicano, pp. 35-38. In: R. Díaz, J.F Álvarez y A Huerta. (eds.). Memorias del foro Regional de Agricultura Sostenible. Sociedad Mexicana de Agricultura Sostenible A. C. Noviembre. Colegio de Postgraduados, Campus Puebla, México. [ Links ]
SPRAGUE G.F. 1966. Plant Breeding. Ames, IA, USA: Iowa State University Press. [ Links ]
URIEL E., ALDAS J. 2005. Análisis multivariante aplicado. Thomson, Madrid, España. 523 p. [ Links ]
YAQOOB M., HASSAN G., MAHMOOD G., SHAH N.H. 1997. Combining ability studies for some quality traits in cotton (Gossypium hirsutum L.). J. Pure Appl Sci. 16:47-50. [ Links ]
1. Facultad de Ciencias Agrícolas, Pecuarias, Forestales y Veterinarias de la UMSS, Casilla 3392, Cochabamba, Bolivia.
2. Facultad de Ciencias Agrícolas, Pecuarias, Forestales y Veterinarias de la UMSS, Casilla 3392, Cochabamba, Bolivia. Correo electrónico: j.gabriel@proinpa.org
3. Fundación para la Promoción e Investigación de Productos Andinos (PROINPA). Casilla 4285. Cochabamba. Bolivia.
4. Fundación para la Promoción e Investigación de Productos Andinos (PROINPA). Casilla 4285. Cochabamba. Bolivia.
5. Facultad de Ciencias Agrícolas, Pecuarias, Forestales y Veterinarias de la UMSS, Casilla 3392, Cochabamba, Bolivia.
6. Fundación para la Promoción e Investigación de Productos Andinos (PROINPA). Casilla 4285. Cochabamba. Bolivia.
Recibido: 20/05/14 Aceptado: 20/11/14


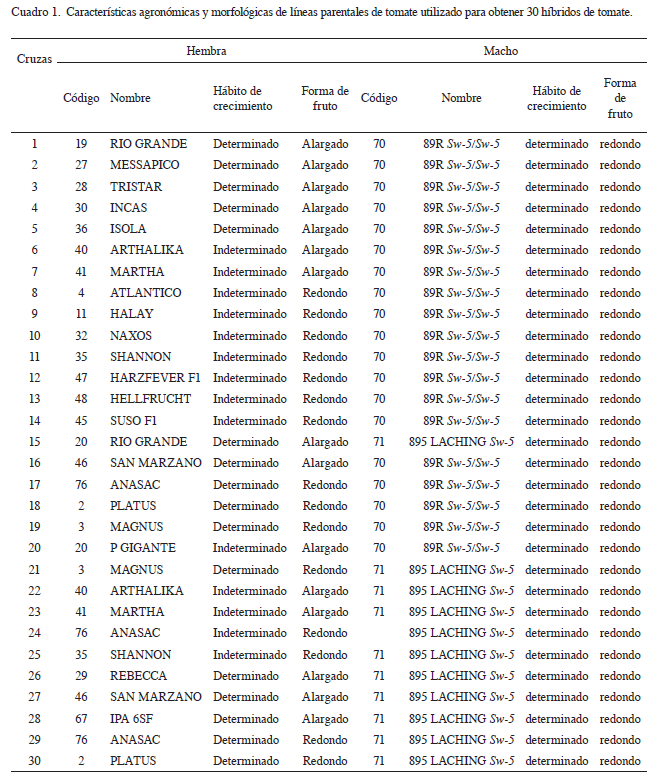{kind=link}
{kind=link}
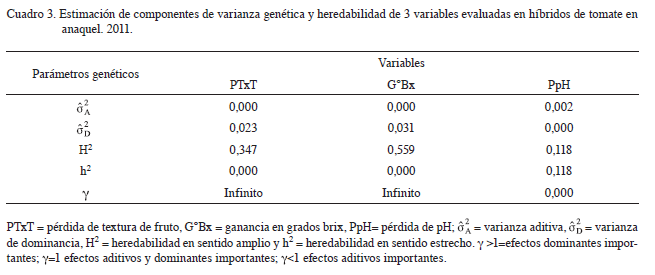{kind=link}
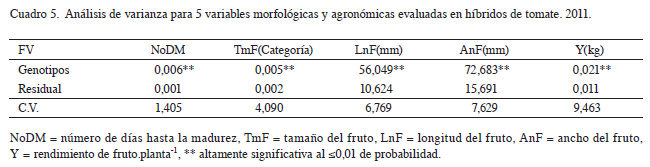{kind=link}
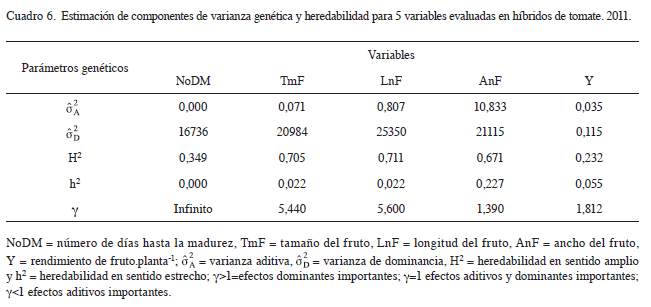{kind=link}
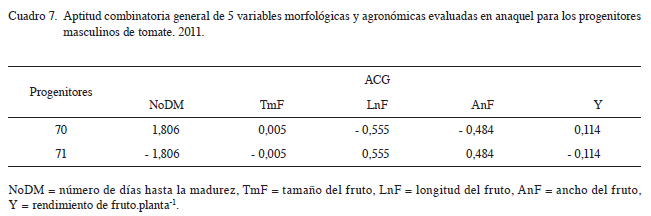{kind=link}