










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Costarricense
Print version ISSN 0377-9424
Agron. Costarricense vol.39 n.1 San Pedro de Montes de Oca Jan./Jun. 2015
Respiración, biomasa microbiana y actividad fosfatasa del suelo en dos agroecosistemas y un bosque en Turrialba, Costa Rica
Respiration, microbial biomass and soil phosphatase activity in two agroecosystems and one forest in Turrialba, Costa Rica
Respiration, microbial biomass and soil phosphatase activity in two agroecosystems and one forest in Turrialba, Costa Rica
Resumen
Con el fin de evaluar algunas características microbiológicas y bioquímicas, se realizó un estudio comparativo entre 3 sistemas de manejo del suelo en Ultisoles, ubicado en Grano de Oro, Turrialba Costa Rica. Se eligieron 3 sistemas de manejo de suelos (2 agroecosistemas, café y cafébanano) y bosque. En cada finca se colectaron 4 muestras compuestas de suelo, a las cuales se les determinó respiración, biomasa microbiana y actividad de la enzima fosfatasa. En los suelos del bosque la biomasa microbiana fue estadísticamente superior (423 mg C kg-1) comparada con la de los agroecosistemas café y café-banano (77 y 111 mg C kg-1 respectivamente). La respiración microbiana no presentó diferencias (580, 560 y 570 µg CO2 g-1.día-1 en los sistemas café, café-banano y bosque, respectivamente). También se determinó que la actividad de la enzima fosfatasa en los suelos del bosque fue estadísticamente más alta (4432 µg p-NP g-1.h-1). Los resultados sugieren que las condiciones del suelo en el bosque favorecen una mayor biomasa microbiana y mayor actividad de la fosfatasa, comparados con los sistemas agrícolas.
Palabras clave: Respiración del suelo; biomasa microbiana; fosfatasa, agroecosistema; bosque.
Abstract
In order to evaluate some microbiological and biochemical characteristics, a comparative study was carried out, as related to 3 different land uses in Ultisols located in Grano de Oro, Turrialba, Costa Rica. Three soil management systems were selected (two agroecosystems, coffee and coffee-banana) and forest. In each farm, 4 composite soil samples were collected, on which microbial biomass and respiration, and phosphatase enzyme activity analysis were performed. The microbial biomass in forest was statistically higher (423 mg C kg-1) compared to those in agroecosystems coffee and coffee-banana (77 and 111 mg C kg-1 respectively). Microbial respiration did not show differences due to land management (580, 560 and 570 µg CO2 g-1.day-1 in coffee, coffee-banana and forest systems, respectively). It was also determined that the enzyme phosphatase activity in forest soils was statistically higher (4432 µg p-NP g-1.h-1). The data suggest that soil conditions in the forest favor greater microbial activity and phosphatase biomass, as compared to agricultural systems.
Keywords: Soil respiration; microbial biomass; phosphatase; agroecosystem; forest.
Introducción
La actividad agrícola intensiva dirigida hacia una mayor productividad ha provocado consecuencias negativas a los ecosistemas. Estas se observan más acentuadas en suelos bajo monocultivos, los cuales se encuentran expuestos a factores erosivos del ambiente y al uso de altas cantidades de agroquímicos, lo que contribuye a la contaminación de las aguas y la disminución de la diversidad biológica del suelo (Muschler 2004).
En Costa Rica, el café es un cultivo muy importante, que en sus inicios se sembró como monocultivo sin sombra en tierras deforestadas, posteriormente surgieron varios modelos agroforestales que utilizaron como sombra el banano (Musa AAA.), la guaba (Inga sp.) y el poró (Eritrina sp.) entre otros, con el fin de mejorar la protección del suelo a la erosión y el aporte de nutrientes, por medio de la biomasa que forma la hojarasca (Beer et ál. 1998, Granados 2004). Por otro lado, el cultivo de café asociado con sombra diversificada con frutales y árboles maderables permite ingresos complementarios para mejorar la economía de los productores (Albertin y Nair 2004), lo que puede resultar en un beneficio socialmente más aceptable, económicamente más viable y ambientalmente más sostenible (Bote y Struik 2011).
Una de las alternativas de sombra temporal para las plantaciones de café, que sirve además para el autoconsumo familiar es el cultivo de banano. En los últimos 20 años el banano sembrado en los cafetales ha pasado a formar parte del comercio de las fincas convirtiéndose en una fuente importante de ingresos, especialmente cuando los precios del café son bajos (Siles et ál. 2010).
Los parámetros más utilizados en las evaluaciones biológicas del suelo son la respiración, la biomasa microbiana y la actividad enzimática, propiedades que responden de manera rápida ante cambios en el sistema (Ros et ál. 2002, Hernández y García 2003).
La respiración es el proceso de oxidación de sustratos orgánicos a dióxido de carbono (CO2) por los microorganismos del suelo (Paul 2007) que generan energía para el crecimiento y mantenimiento microbiano (Jacinthe y Lal 2005). La biomasa microbiana se considera el principal agente para la descomposición de la materia orgánica del suelo (MOS), las transformaciones de nutrimentos (Brookes et ál. 2008), la estabilidad estructural y como indicador de contaminación del suelo (Wright e Islam 2006). Las enzimas son proteínas especializadas que se combinan con un sustrato específico y actúan para catalizar una reacción sin experimentar cambios en su estructura; en el suelo, las enzimas son esenciales para la transformación de energía y el ciclaje de nutrientes (Coyne 2000, Paul 2007, Kandeler 2007).
Por su parte las enzimas fosfatasas participan en la hidrólisis de diversos ésteres de fosfato orgánico e inorgánico en el ciclo del fósforo (Trasar et ál. 2003). Estas enzimas son sintetizadas y secretadas extracelularmente por bacterias u hongos, que forman parte de la matriz del suelo (Tripathi et ál. 2007).
El presente estudio tuvo como objetivo comparar entre los agroecosistemas (café, asocio cafébanano) y un bosque, la respiración, biomasa microbiana y actividad fosfatasa ácida del suelo.
Materiales y Métodos
Las muestras de suelo se colectaron en mayo del 2012 en 2 sistemas de cultivo (café, asocio café-banano) y un bosque en las estribaciones de la cordillera de Talamanca en Grano de Oro, cantón Turrialba. La zona de vida corresponde a Bosque muy húmedo premontano con un rango de temperatura de 18 a 24°C y precipitación de 4000 a 8000 mm (Centro Científico Tropical y Ministerio de Agricultura 2004). Las coordenadas del punto central equidistante a los 3 sistemas son: -83,4698725 (longitud) y 9,8293777 (latitud) y la altitud es de 1124 msnm.
Las muestras se llevaron al Laboratorio de Recursos Naturales, Laboratorio de Suelos y Foliares y Laboratorio de Microbiología Agrícola del Centro de Investigaciones Agronómicas (CIA) de la Universidad de Costa Rica (UCR) para los respectivos análisis físicos, químicos y microbiológicos.
Muestreo de suelos
En cada uno de los sitios seleccionados se procedió, con la ayuda de un barreno, a colectar 4 muestras compuestas (provenientes de 15 sub-muestras) a 2 profundidades: 0-10 y de 10-20 cm. Para los análisis microbiológicos, las muestras colectadas en el campo se almacenaron a una temperatura aproximada de 4oC, hasta su procesamiento.
Análisis físico y químico de suelos
Los análisis físicos y químicos de suelos se determinaron de acuerdo con los métodos utilizados en los laboratorios del CIA descritos por Henríquez y Cabalceta (2012) mediante la utilización de las soluciones extractoras Olsen Modificado y KCl 1M en proporción 1:10. Los elementos Ca, Mg, K, Fe, Mn, Cu y Zn fueron determinados por absorción atómica y el P colorimétricamente. El pH se determinó en agua a una relación 1:2,5 y la acidez intercambiable extraída con KCl 1M por titrimetría. La textura se realizó mediante el método del hidrómetro de Bouyoucus.
Determinación de la biomasa microbiana de Carbono
La biomasa microbiana se determinó por medio de la metodología de fumigación extracción (Vance et ál. 1987), para lo cual se pesaron 10 g de muestra (control) en un erlenmeyer de 250 ml, se adicionó 50 ml de K2SO4 (0,5M), se agitó 30 min y se filtró con un papel Whatman 42. Se guardó el extracto a 4oC hasta su procesamiento. También se pesó 10 g de muestra (fumigado) en un beaker de 40 ml, se fumigó con cloroformo libre de etanol, se incubó durante 24 h a 25oC. Se procedió igual que la muestra control. Del extracto obtenido se colocó 8 ml en tubos de digestión (3 repeticiones control y 3 fumigado), se adicionó 2 ml de K2Cr2O7 (66,7 mM), 70 mg de HgO y 15 ml de mezcla ácida (2v H2SO4 (98%) y 1v H3PO4 (88%). Las muestras se digirieron a 150oC por 30 minutos y se dejaron enfriar. Se transfirieron a un erlenmeyer de 250 ml, se adicionó 80 ml de agua destilada, se agregó 3 gotas de fenontralina y se valoró el exceso de K2Cr2O7 con sal de Morh 33,3 mM (NH4)2.Fe(SO4)2.6(H2O).
Determinación de la respiración microbiana del suelo
La respiración microbiana del suelo se determinó de acuerdo con la metodología de Anderson (1982), se pesó en beakers 10 g de suelo por triplicado, la muestra se colocó en una jarra de incubación junto con un recipiente que contenía 10 ml de agua y otro con 10ml de NaOH (1M) y se incubaron a 25oC. A los 4 días se determinó la cantidad de CO2 absorbido en el NaOH, para lo cual se adicionó 10 ml de BaCl. Se transfirió esta mezcla a un erlenmeyer y se adicionó 3 gotas de fenolftaleína, se tituló la solución con HCl (0,5M). Después de 4 días se repitió este proceso. Se determinó el CO2 generado en los 8 días de incubación.
Determinación de la enzima fosfatasa
Se evaluó la actividad de las fosfatasas ácidas según la metodología de Tabatabai y Bremner (1969) en la forma descrita en Henríquez et ál. (2014).
Análisis estadístico
Los datos obtenidos provenientes de las 4 muestras de cada sitio, se analizaron por medio de un análisis de varianza. La comparación de medias de los tratamientos se realizó mediante la prueba de Tukey al 0,05% de probabilidad, por medio del programa estadístico Infostat.
Resultados y Discusión
Caracterización física y química de los suelos
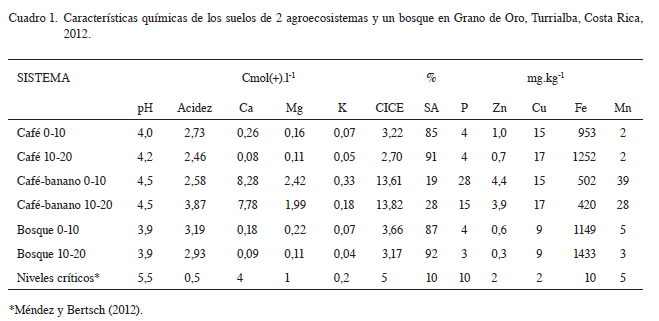
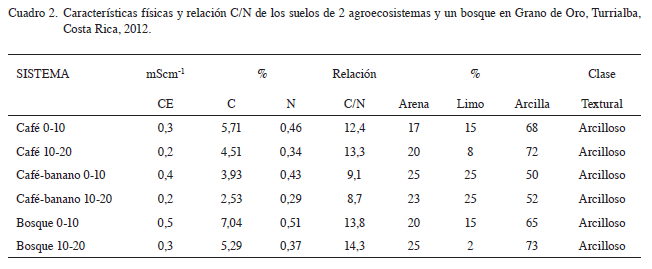
De acuerdo con las características edáficas evaluadas en el sitio, los suelos en estudio se clasificaron como Andyc Haplohumults y pertenecen al orden de los Ultisoles con características ándicas, lo que concuerda con el Mapa Digital de Suelos de Costa Rica del 2013 (Mata et ál. 2013). Se encuentran ubicados en las estribaciones de la cordillera de Talamanca a 1100 msnm con un régimen de humedad údico, con altas cantidades de materia orgánica en la parte superior del perfil, químicamente son suelos con bajos contenidos de bases y alta saturación de acidez, son arcillosos, bien drenados y con relieve ligeramente ondulado (Cuadro 1 y 2). La textura en estos suelos tiene un rol importante, en el que principalmente destacan los altos contenidos de arcilla (50% a 73%), con un dominio de texturas muy finas de más de 60 por ciento de arcillas, valor que se incrementa con la profundidad, lo cual se relaciona directamente con el comportamiento del agua y el aire, ya que estos suelos se saturan fácilmente disminuyendo el flujo de aire, en ecosistemas como el bosque con mayor cantidad de materia orgánica y mejor estructura, el proceso de aireación ocurre más rápidamente y en agro ecosistemas mas alterados al bajar los contenidos de materia orgánica los suelos permanecen más tiempo con exceso de agua.
En cuanto a los contenidos nutricionales en los sistemas café y bosque, los suelos presentaron deficiencias en la mayoría de los elementos según niveles críticos para suelos (Méndez y Bertsch 2012), lo que no se observó en el asocio cafébanano, debido posiblemente a la influencia de la fertilización al café y la materia orgánica generada por la biomasa del banano. En el Cuadro 1 se puede apreciar que tanto en los agroecosistemas como en el bosque los cationes, el carbono y el nitrógeno disminuyen en la profundidad 10-20 cm, en comparación con la de 0-10 cm.
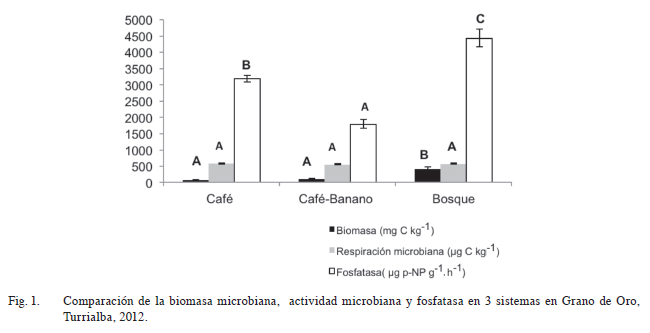
Los resultados de la Figura 1, muestran que el suelo del bosque presenta una biomasa microbiana de 423 mg C kg-1 estadísticamente superior a los agroecosistemas café-banano y café que presentaron valores de 111 y 77 mg C kg-1 respectivamente. Estos resultados concuerdan con lo reportado por Porras (2006) en sistemas agroforestales de café y en bosque, así como lo mencionado por Jenkinson y Ladd (1981), en relación con las situaciones que favorecen la acumulación de materia orgánica, incrementan la cantidad de biomasa y la proporción de esta biomasa en la materia orgánica del suelo, como sucede en los suelos del bosque. Del mismo modo Castro (1995) determinó que la biomasa microbiana en suelos sometidos a cultivos se redujo significativamente en relación con los suelos bajo barbecho. En su estudio tanto los Ultisoles como los Andisoles presentaron menor biomasa microbiana cuando estuvieron bajo cultivo. Así mismo, Paz et ál. (2006), indican que existe mayor biomasa microbiana en un sistema de café con sombra comparado con el café sin sombra, y que la mayor cantidad de biomasa microbiana en estos últimos, se asocia con el mayor contenido de materia orgánica presente en esos sistemas.
Las diferencias observadas en la biomasa microbiana encontrada entre el suelo del bosque y los 2 agroecosistemas evaluados concuerda con la pérdida de la materia orgánica del suelo que se observa claramente en el Cuadro 2, donde el carbono del suelo en los cultivos de café y asocio café-banano es menor que en el bosque, en los agroecosistemas se extrae parte de la biomasa del cultivo (cosecha y podas), mientras que en el bosque se da un constante reciclaje de la biomasa, otro factor a considerar es el efecto erosivo de la lluvia y la pendiente del terreno. Castro (1995) encontró en Ultisoles y Andisoles de 2 zonas de Costa Rica, una reducción promedio de un 40% en el contenido de C con el cambio de barbecho a cultivo. En los Ultisoles de las 2 localidades muestreadas, los valores de biomasa en el suelo en barbecho fueron muy similares; sin embargo esta se redujo al estar bajo cultivo en 42% en San Carlos y en 75% en Buenos Aires. La correlación entre los contenidos de biomasa y de materia orgánica fue de 0,91 para estos suelos.
Sparling et ál. (1992) reportan que después de 11 años de cultivo continuo de maíz y pastos, la biomasa microbiana se redujo en 49 y 60% respectivamente, en los primeros 20 cm del suelo en comparación con un suelo de pradera permanente. Del mismo modo Zagal y Córdova (2005), reportan que luego de 8 años de manejo de cultivos intensivos y con diferentes rotaciones encontraron que el C biomásico disminuyó con la mayor intensidad de uso del suelo, de 551 a 264 mg.g-1. También en Costa Rica, Sánchez et ál. (2006) encontraron al comparar la producción de café en diferentes sistemas de gestión agroforestal, que la biomasa microbiana se incrementó de 583 mg.kg-1 en un sistema agroforestal tratado de manera convencional a 913 mg.kg-1 en un sistema agroforestal orgánico.
En lo referente a la respiración microbiana, en este trabajo no se observaron diferencias entre los valores de producción de CO2, encontrados entre los 3 sistemas evaluados (Figura 1), que presentó un promedio de 580 μg CO2 g-1.día-1. Al respecto Álvarez y Anzueto (2004) encontraron en estudios realizados en maíz bajo diferentes sistemas de siembra, que la respiración microbiana fue menor en parcelas bajo cultivo anual continuo o de rotación pastizal-cultivo que en sistemas con barbecho. En otro estudio, Souza et ál. (2012) concluyeron que la respiración microbiana mostró mayores valores en los bosques de referencia en comparación con sistemas agroforestales y el sistema de café sin sombra. Así mismo, Chavarría et ál. (2012) en diferentes sistemas de manejo en café, reportan que los valores más altos de respiración microbiana se registraron en el sistema bosque (640 μg C-CO2 g-1.día-1) seguido del sistema café sin sombra (440 μg C-CO2 g-1.día-1).
Cómo se observa en la Figura 1, la actividad de la enzima fosfatasa disminuyó considerablemente en el agroecosistema café-banano (1791 µg p-NP g-1.h-1), en comparación con el café (3185 µg p-NP g-1.h-1) y el bosque (4433 µg p-NP g-1.h-1). Al respecto Acosta et ál. (2007) encontraron menores valores de fosfatasa en suelos bajo agricultura y similares para bosque y pastura (agricultura<bosque=pastura), ya que entre más labrado el sistema, los procesos de degradación de materia orgánica se aceleran debido a una mayor exposición de la misma a los factores ambientales.
Por su parte en Costa Rica, Henríquez et ál. (2014) encontraron una menor actividad de la enzima fosfatasa en el cultivo de palma (275 µg PNP g-1.h-1) en relación con el cultivo de vainilla (3505 µg PNP g-1.h-1) e indican que los resultados de la actividad enzimática depende de aspectos como el tipo de cultivo, manejo agronómico y características intrínsecas del suelo.
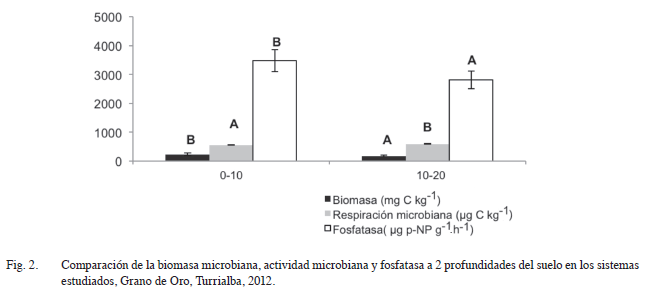
Con respecto al factor profundidad de muestreo, se observó un mayor valor de biomasa microbiana en el estrato superficial de 0-10 cm (231 mg C kg-1) que la encontrada en el estrato de 10-20 cm (176 mg C kg-1) (Figura 2), estos valores coinciden con la mayor cantidad de MOS en los primeros centímetros del suelo (Cuadro 1). Contrariamente, se observó una respiración ligeramente mayor en la profundidad de 10-20 cm (590 μg CO2 g-1.día-1) en relación al estrato superior 0-10 cm (540 μg CO2 g-1.día-1), lo cual podría deberse al efecto de diferentes factores ambientales (temperatura, nutrientes) en la producción de CO2 a través del perfil del suelo (Fierer et ál. 2003), es importante investigar el comportamiento de esta variable en relación con la profundidad, temperatura y humedad del suelo.
En la Figura 2 se puede apreciar una mayor actividad de la enzima fosfatasa en los primeros centímetros del suelo (3470μg p-NP g-1.h-1), lo cual puede deberse a la mayor cantidad de materia orgánica presente en el estrato superior (Cuadro 1), en relación con la capa más profunda de 10-20 cm (2803μg p-NP g-1.h-1). Autores como Yoshioka et ál. (2006), mencionan que en el cultivo de plátano la mayor actividad de la fosfatasa ácida se presentó a los 0-5 cm y difirió significativamente del resto de profundidades evaluadas en su trabajo. Otros investigadores también presentan resultados que demuestran que la actividad de las fosfatasas disminuye con la profundidad del suelo (Dick 1984, Sánchez de P. 2003), lo cual se asocia con la disminución de la materia orgánica, la microbiota y algunos macro y micro elementos, así como la actividad rizosférica.
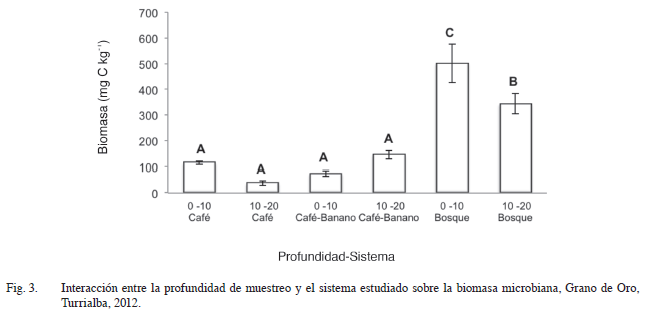
Se observó interación entre la profundidad de suelo y el sistema en la variable biomasa microbiana (Figura 3). Podemos apreciar que el sistema bosque presentó una mayor biomasa microbiana en las 2 profundidades evaluadas (502,0 y 343,5 mg C kg-1) separándose estadísticamente de los agroecosistemas evaluados. Estos resultados indican que la biomasa microbiana del bosque fue mayor que la encontrada en los agroecosistemas estudiados, donde se reduce por los factores propios del manejo de los cultivos.
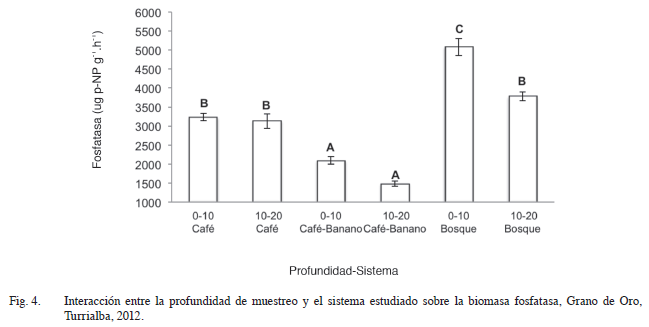
Cuando se comparó la interacción profundidad de suelo-sistema para la variable fosfatasa (Figura 4), se encontró que existen diferencias entre éstas, donde el asocio café-banano presentó la menor actividad enzimática, lo cual puede tener estrecha relación con la mayor cantidad de fósforo presente en el suelo de éste agroecosistema (Cuadro 1) y a lo citado por Schneider et ál. (2000) que mencionan que las fosfatasas se producen cuando el fósforo es el elemento que limita el crecimiento. Los mayores niveles de fosfatasa se encontraron en el bosque, lo que coincide con la ausencia de fertilización en este sistema y la importancia del aporte de P orgánico en el mismo.
Agradecimientos
A la Secretaría Nacional de Educación Superior, Ciencia, Tecnología e Innovación de Ecuador (SENESCYT) y al Instituto Nacional de Investigaciones Agropecuarias (INIAP) de Ecuador por el financiamiento y permiso para los estudios de maestría.
A los proyectos: VI-733-BO-507 “Determinación de la actividad enzimática en el suelo” y VI-733-A1-821 “Laboratorio de Microbiología Agrícola y Venta de Servicios” de la Universidad de Costa Rica, por el financiamiento de esta investigación.
Literatura Citada
ACOSTA V., CRUZ L., SOTOMAYOR D., PÉREZ L. 2007. Enzyme activities as affected by soil propertiesand land use in a tropical watershed. Applied Soil Ecology 35(1):35-45. [ Links ]
ALBERTIN A., NAIR P. 2004. Farmers’ perspectives on the role of Shade trees in coffee production systems: an assessment from the Nicoya Peninsula, Costa Rica. Human Ecology 32(4):443-463. [ Links ]
ÁLVAREZ J., ANZUETO M. 2004. Actividad microbiana del suelo bajo diferentes sistemas de producción de maíz en los altos de Chiapas, México. Agrociencia 38:13-22. [ Links ]
ANDERSON J. 1982. Measurement of CO2 evolution rates, pp. 831-871. In: K. Alef and P. Nannipieri (ed). Methods in Applied Soil Microbiology and Biochemistry. [ Links ]
BEER J., MUSCHLER R., KASS D., SOMARRIBA E. 1998. Shade management in coffee and cacao plantations. Agroforestry Systems 38:139-164. [ Links ]
BOTE A., STRUIK P. 2011. Effects of shade on growth, production and quality of coffee (Coffea arabica) in Ethiopia. Journal of Horticulture and Forestry 3(11):336-341. [ Links ]
BROOKES P., CAYUELA M., CONTIN M., DE NOBILI M., KEMMITT S., MONDINI C. 2008. The mineralisation of fresh and humified soil organic matter by the soil microbial biomass. Waste Management 28(4):716-722. [ Links ]
CASTRO L. 1995. Efecto del uso agrícola y el barbecho sobre los contenidos de biomasa microbiana de Ultisoles y Andisoles de Costa Rica. Agronomía Costarricense 19(2):59-65. [ Links ]
CENTRO CIENTÍFICO TROPICAL Y MINISTERIO DE AGRICULTURA. 2004. Mapa Ecológico de Costa Rica, según el sistema de clasificación de Zonas de Vida del mundo de L. R. Holdridge. Esc: 1: 200000. Color. [ Links ]
CHAVARRÍA N., TAPIA A., SOTO G., VIRGINIO E. 2012. Efecto de diferentes sistemas de manejo sobre la calidad del suelo, en fincas cafetaleras de la zona de Turrialba y Orosi. Revista InterSedes 8(26):85-105. [ Links ]
COYNE M. 2000. Microbiología del suelo: un enfoque exploratorio. Editorial Paraninfo, España. 416 p. [ Links ]
DICK W. 1984. Influence of long-term tillage and crop rotation combinations on soil enzyme activities. Soil Science Society America Journal 48:569-574. [ Links ]
FIERER N., ALLEN A., SCHIMEL J., HOLDEN P. 2003. Control on microbial CO2 production: a comparison on surface and subsurface soils horizons. Global Change Biology 9:1322-1332. [ Links ]
GRANADOS C. 2004. El impacto ambiental del café en la historia costarricense. Diálogos Revista Electrónica de Historia 4(2):42. [ Links ]
HENRÍQUEZ C., CABALCETA G. 2012. Guía práctica para el estudio introductorio de los suelos con un enfoque agrícola. 2 ed. San José, C.R. Asociación Costarricense de la Ciencia del Suelo. 111 p. [ Links ]
HENRÍQUEZ C., URIBE L., VALENCIANO A., NOGALES R. 2014. Actividad enzimática del suelo–deshidrogenasa, glucosidasa, fosfatas y ureasabajo diferentes cultivos. Agronomía Costarricense 38(1):43-54. [ Links ]
HERNÁNDEZ T., GARCÍA C. 2003. Estimación de respiración microbiana del suelo, pp. 311-346. In: C. García, F. Gil, T. Hernández y C. Trasar. (eds.). Técnicas de Análisis de Parámetros Bioquímicos en Suelos. Mundi-Prensa. Madrid. [ Links ]
JACINTHE P., LAL L. 2005. Respiration, pp. 1508-1512. In: R. Lal (ed.). Encyclopedia of Soil Science, Second Edition. Taylor and Francis Group, New York, USA. [ Links ]
JENKINSON D., LADD J. 1981. Microbial biomass in soils: measurement and turnover, pp. 415-417. In: E.A. Paul and J.N. Ladd (eds.). Soil Biochemistry Vol. 5. Marcel Dekker, New York. [ Links ]
KANDELER E. 2007. Physiological and biochemical methods for studying soil biota and their function, pp. 53-84. In: E. Paul (ed.). Soil microbiology, ecology and biochemistry. Academic Press. Elsevier. [ Links ]
MATA R., ROSALES A., VÁSQUEZ A., SANDOVAL D. 2013. Mapa de Subórdenes de Suelos de Costa Rica. 2a ed. San José, Costa Rica. Asociación Costarricense de la Ciencia del Suelo. Esc 1: 200.000. Color. [ Links ]
MÉNDEZ J., BERTSCH F. 2012. Guía para la interpretación de la fertilidad de los suelos en Costa Rica. ACCS. 1 ed. San José CR. 108 p. [ Links ]
MUSCHLER R. 2004. Shade management and its effect on coffee growth and quality, pp. 339-353. In: J.N. Wintgens (ed.). Coffee: growing, processing, sustainable production. WileyVCH verlag. [ Links ]
PAUL E.A. 2007. Soil microbiology, ecology and biochemistry. Academic press. 532 p. [ Links ]
PAZ I., SÁNCHEZ DE P.M., SADEGHIAN S. 2006. Relación entre dos sistemas de sombrío de café y algunas propiedades del suelo en la meseta de Popayán, CO. Acta Agronómica 54(4):1-6. [ Links ]
PORRAS C. 2006. Efecto de los sistemas agroforestales de café orgánico y convencional sobre las características de suelos en el Corredor Biológico Turrialba–Jiménez, Costa Rica. Tesis de maestría, CATIE, Turrialba, Costa Rica. 131 p. [ Links ]
ROS M., HERNÁNDEZ T., GARCÍA C. 2002. Bioremediation of Soil Degraded by Sewage Sludge: Effects on Soil Properties and Erosion Losses. Environmental Management 31:741-747. [ Links ]
SÁNCHEZ DE LEÓN Y., DE MELO E., SOTO G., JOHNSON J., LUGO J. 2006. Earthworm Populations, Microbial Biomass and Coffee Production in Different Experimental Agroforestry Management Systems in Costa Rica. Caribbean Journal of Science 42(3):397-409. [ Links ]
SÁNCHEZ DE P.M. 2003. Actividad biológica en la rizósfera del maracuyá–Pasiflora edulis var flavicarpa en diferentes sistemas de manejo, estados de desarrollo y condiciones fitosanitarias. Tesis de doctorado, Universidad Politécnica de Madrid. 261 p. [ Links ]
SCHNEIDER K., TURRION M.B., GALLARDO J.F. 2000. Modified method for measuring acid phosphatase activities in forest soils with high organic matter content. Communications in Soil Science and Plant Analysis 31:3077-3088. [ Links ]
SILES P., BUSTAMANTE O., DERAS M., MATUTE O., AGUILAR C., ROJAS J., CASTELLON J., BURKHARDT J., STAVER C. 2010. Bananos en cafetales con árboles en América Latina: estrategias preliminares para mejorar su productividad, rentabilidad y sostenibilidad. XIX Reunión Internacional ACORBAT. Medellín, CO. [ Links ]
SOUZA H., DE GOEDE R., BRUSSAARD L., CARDOSO I., DUARTE E., FERNANDES R., GOMES L., PULLEMAN M. 2012. Protective shade, tree diversity and soil properties in coffee agroforestry systems in the Atlantic rainforest biome. Agriculture Ecosystems and Environment 146:179-196. [ Links ]
SPARLING G., SHEPHERD T., KETTLES H. 1992. Changes in soil organic C, microbial C and aggregate stability under continuous maize and cereal cropping and after restoration to pasture in soils from the Manawatu region New Zealand. Soil Tillage Research 24:225-241. [ Links ]
TABATABAI M., BREMNER J. 1969. Use of p-nitrophenyl phosphate for assay of soil phosphate activity. Soil Biology and Biochemistry 1:301-307. [ Links ]
TRASAR G., GIL-SOTRES F., LEIROS M. 2003. Determinación de la actividad fosfatasa del suelo, pp 52-76. In: C. García, F. Gil, T. Hernández y C. Trasar (eds.). Técnicas de Análisis de Parámetros Bioquímicos en Suelos. Mundi-Prensa. Madrid. [ Links ]
TRIPATHI S., CHAKRABORTY A., CHAKRABARTI A., BANDYOPADHYAY B. 2007. Enzyme activities and microbial biomass in coastal soils of India. Soil Biology and Biochemistry 39:2840-2848. [ Links ]
VANCE E., BROOKES P., JENKINSON D. 1987. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry 19:703-707. [ Links ]
WRIGHT S., ISLAM K. 2005. Microbial biomass measurement methods, pp. 1067-1070. In: R. Lal (ed.). Encyclopedia of Soil Science, Second Edition. Taylor and Francis Group, New York. [ Links ]
YOSHIOKA I., SÁNCHEZ DE P.M., BOLAÑOS M. 2006. Actividad de fosfatasas ácida y alcalina en suelo cultivado con plátano en tres sistemas de manejo. Acta Agronómica 55. Consultado 3 de febrero de 2015 Disponible en http://www.redalyc.org/articulo. oa?id=169920417001 [ Links ]
ZAGAL E., CÓRDOVA C. 2005. Indicadores de calidad de la materia orgánica del suelo en un Andisol cultivado. Agricultura Técnica 65:186-197. [ Links ]
ALBERTIN A., NAIR P. 2004. Farmers’ perspectives on the role of Shade trees in coffee production systems: an assessment from the Nicoya Peninsula, Costa Rica. Human Ecology 32(4):443-463. [ Links ]
ÁLVAREZ J., ANZUETO M. 2004. Actividad microbiana del suelo bajo diferentes sistemas de producción de maíz en los altos de Chiapas, México. Agrociencia 38:13-22. [ Links ]
ANDERSON J. 1982. Measurement of CO2 evolution rates, pp. 831-871. In: K. Alef and P. Nannipieri (ed). Methods in Applied Soil Microbiology and Biochemistry. [ Links ]
BEER J., MUSCHLER R., KASS D., SOMARRIBA E. 1998. Shade management in coffee and cacao plantations. Agroforestry Systems 38:139-164. [ Links ]
BOTE A., STRUIK P. 2011. Effects of shade on growth, production and quality of coffee (Coffea arabica) in Ethiopia. Journal of Horticulture and Forestry 3(11):336-341. [ Links ]
BROOKES P., CAYUELA M., CONTIN M., DE NOBILI M., KEMMITT S., MONDINI C. 2008. The mineralisation of fresh and humified soil organic matter by the soil microbial biomass. Waste Management 28(4):716-722. [ Links ]
CASTRO L. 1995. Efecto del uso agrícola y el barbecho sobre los contenidos de biomasa microbiana de Ultisoles y Andisoles de Costa Rica. Agronomía Costarricense 19(2):59-65. [ Links ]
CENTRO CIENTÍFICO TROPICAL Y MINISTERIO DE AGRICULTURA. 2004. Mapa Ecológico de Costa Rica, según el sistema de clasificación de Zonas de Vida del mundo de L. R. Holdridge. Esc: 1: 200000. Color. [ Links ]
CHAVARRÍA N., TAPIA A., SOTO G., VIRGINIO E. 2012. Efecto de diferentes sistemas de manejo sobre la calidad del suelo, en fincas cafetaleras de la zona de Turrialba y Orosi. Revista InterSedes 8(26):85-105. [ Links ]
COYNE M. 2000. Microbiología del suelo: un enfoque exploratorio. Editorial Paraninfo, España. 416 p. [ Links ]
DICK W. 1984. Influence of long-term tillage and crop rotation combinations on soil enzyme activities. Soil Science Society America Journal 48:569-574. [ Links ]
FIERER N., ALLEN A., SCHIMEL J., HOLDEN P. 2003. Control on microbial CO2 production: a comparison on surface and subsurface soils horizons. Global Change Biology 9:1322-1332. [ Links ]
GRANADOS C. 2004. El impacto ambiental del café en la historia costarricense. Diálogos Revista Electrónica de Historia 4(2):42. [ Links ]
HENRÍQUEZ C., CABALCETA G. 2012. Guía práctica para el estudio introductorio de los suelos con un enfoque agrícola. 2 ed. San José, C.R. Asociación Costarricense de la Ciencia del Suelo. 111 p. [ Links ]
HENRÍQUEZ C., URIBE L., VALENCIANO A., NOGALES R. 2014. Actividad enzimática del suelo–deshidrogenasa, glucosidasa, fosfatas y ureasabajo diferentes cultivos. Agronomía Costarricense 38(1):43-54. [ Links ]
HERNÁNDEZ T., GARCÍA C. 2003. Estimación de respiración microbiana del suelo, pp. 311-346. In: C. García, F. Gil, T. Hernández y C. Trasar. (eds.). Técnicas de Análisis de Parámetros Bioquímicos en Suelos. Mundi-Prensa. Madrid. [ Links ]
JACINTHE P., LAL L. 2005. Respiration, pp. 1508-1512. In: R. Lal (ed.). Encyclopedia of Soil Science, Second Edition. Taylor and Francis Group, New York, USA. [ Links ]
JENKINSON D., LADD J. 1981. Microbial biomass in soils: measurement and turnover, pp. 415-417. In: E.A. Paul and J.N. Ladd (eds.). Soil Biochemistry Vol. 5. Marcel Dekker, New York. [ Links ]
KANDELER E. 2007. Physiological and biochemical methods for studying soil biota and their function, pp. 53-84. In: E. Paul (ed.). Soil microbiology, ecology and biochemistry. Academic Press. Elsevier. [ Links ]
MATA R., ROSALES A., VÁSQUEZ A., SANDOVAL D. 2013. Mapa de Subórdenes de Suelos de Costa Rica. 2a ed. San José, Costa Rica. Asociación Costarricense de la Ciencia del Suelo. Esc 1: 200.000. Color. [ Links ]
MÉNDEZ J., BERTSCH F. 2012. Guía para la interpretación de la fertilidad de los suelos en Costa Rica. ACCS. 1 ed. San José CR. 108 p. [ Links ]
MUSCHLER R. 2004. Shade management and its effect on coffee growth and quality, pp. 339-353. In: J.N. Wintgens (ed.). Coffee: growing, processing, sustainable production. WileyVCH verlag. [ Links ]
PAUL E.A. 2007. Soil microbiology, ecology and biochemistry. Academic press. 532 p. [ Links ]
PAZ I., SÁNCHEZ DE P.M., SADEGHIAN S. 2006. Relación entre dos sistemas de sombrío de café y algunas propiedades del suelo en la meseta de Popayán, CO. Acta Agronómica 54(4):1-6. [ Links ]
PORRAS C. 2006. Efecto de los sistemas agroforestales de café orgánico y convencional sobre las características de suelos en el Corredor Biológico Turrialba–Jiménez, Costa Rica. Tesis de maestría, CATIE, Turrialba, Costa Rica. 131 p. [ Links ]
ROS M., HERNÁNDEZ T., GARCÍA C. 2002. Bioremediation of Soil Degraded by Sewage Sludge: Effects on Soil Properties and Erosion Losses. Environmental Management 31:741-747. [ Links ]
SÁNCHEZ DE LEÓN Y., DE MELO E., SOTO G., JOHNSON J., LUGO J. 2006. Earthworm Populations, Microbial Biomass and Coffee Production in Different Experimental Agroforestry Management Systems in Costa Rica. Caribbean Journal of Science 42(3):397-409. [ Links ]
SÁNCHEZ DE P.M. 2003. Actividad biológica en la rizósfera del maracuyá–Pasiflora edulis var flavicarpa en diferentes sistemas de manejo, estados de desarrollo y condiciones fitosanitarias. Tesis de doctorado, Universidad Politécnica de Madrid. 261 p. [ Links ]
SCHNEIDER K., TURRION M.B., GALLARDO J.F. 2000. Modified method for measuring acid phosphatase activities in forest soils with high organic matter content. Communications in Soil Science and Plant Analysis 31:3077-3088. [ Links ]
SILES P., BUSTAMANTE O., DERAS M., MATUTE O., AGUILAR C., ROJAS J., CASTELLON J., BURKHARDT J., STAVER C. 2010. Bananos en cafetales con árboles en América Latina: estrategias preliminares para mejorar su productividad, rentabilidad y sostenibilidad. XIX Reunión Internacional ACORBAT. Medellín, CO. [ Links ]
SOUZA H., DE GOEDE R., BRUSSAARD L., CARDOSO I., DUARTE E., FERNANDES R., GOMES L., PULLEMAN M. 2012. Protective shade, tree diversity and soil properties in coffee agroforestry systems in the Atlantic rainforest biome. Agriculture Ecosystems and Environment 146:179-196. [ Links ]
SPARLING G., SHEPHERD T., KETTLES H. 1992. Changes in soil organic C, microbial C and aggregate stability under continuous maize and cereal cropping and after restoration to pasture in soils from the Manawatu region New Zealand. Soil Tillage Research 24:225-241. [ Links ]
TABATABAI M., BREMNER J. 1969. Use of p-nitrophenyl phosphate for assay of soil phosphate activity. Soil Biology and Biochemistry 1:301-307. [ Links ]
TRASAR G., GIL-SOTRES F., LEIROS M. 2003. Determinación de la actividad fosfatasa del suelo, pp 52-76. In: C. García, F. Gil, T. Hernández y C. Trasar (eds.). Técnicas de Análisis de Parámetros Bioquímicos en Suelos. Mundi-Prensa. Madrid. [ Links ]
TRIPATHI S., CHAKRABORTY A., CHAKRABARTI A., BANDYOPADHYAY B. 2007. Enzyme activities and microbial biomass in coastal soils of India. Soil Biology and Biochemistry 39:2840-2848. [ Links ]
VANCE E., BROOKES P., JENKINSON D. 1987. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry 19:703-707. [ Links ]
WRIGHT S., ISLAM K. 2005. Microbial biomass measurement methods, pp. 1067-1070. In: R. Lal (ed.). Encyclopedia of Soil Science, Second Edition. Taylor and Francis Group, New York. [ Links ]
YOSHIOKA I., SÁNCHEZ DE P.M., BOLAÑOS M. 2006. Actividad de fosfatasas ácida y alcalina en suelo cultivado con plátano en tres sistemas de manejo. Acta Agronómica 55. Consultado 3 de febrero de 2015 Disponible en http://www.redalyc.org/articulo. oa?id=169920417001 [ Links ]
ZAGAL E., CÓRDOVA C. 2005. Indicadores de calidad de la materia orgánica del suelo en un Andisol cultivado. Agricultura Técnica 65:186-197. [ Links ]
1. Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP) de Ecuador y Becario de la Secretaría Nacional de Educación Superior, Ciencia, Tecnología e Innovación del Ecuador (SENESCYT). Correo electrónico: lidieth.uribe@ucr.ac.cr
2. Centro de Investigaciones Agronómicas (CIA) Universidad de Costa Rica, San José. Costa Rica.
3. Centro de Investigaciones Agronómicas (CIA) Universidad de Costa Rica, San José. Costa Rica.
4. Centro de Investigaciones Agronómicas (CIA) Universidad de Costa Rica, San José. Costa Rica.
Recibido: 26/09/14 Aceptado: 06/12/14


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}