










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgronomía Costarricense
Print version ISSN 0377-9424
Agron. Costarricense vol.36 n.2 San Pedro de Montes de Oca Dec. 2012
Nota técnica
Herramientas para un programa de mejoramiento genético del Guayabo (Psidium guajava L.) en Cuba
Tools for a genetic breeding program for guava (Psidium guajava L.) in Cuba
Juliette Valdés-Infante1*/2*, Narciso Nerdo Rodríguez2, Josefa Bárbara Velásquez2, Darío Gaspar Sourd2, Gonzalo González2, Julio Alberto Rodríguez2, Wolfgang Rohde3*
*Dirección para corrrespondencia:
Resumen
El objetivo del presente trabajo es desarrollar un programa de mejoramiento genético del guayabo (Psidium guajava L.) en Cuba, que contribuya a un uso más eficiente de los recursos fitogenéticos del cultivo. El mismo comienza a partir de la colección, con la prospección de genotipos autóctonos, la introducción de accesiones y parientes silvestres con origen geográfico diverso. Dichas actividades son vitales para el posterior enriquecimiento y mantenimiento de la colección, unido a las diferentes formas de propagación sexual y asexual que garantizan la multiplicación del material y permiten crear nuevas fuentes de variabilidad y una vez establecida, se procede a su caracterización. Para esto se refieren un grupo de descriptores mínimos que pueden ser útiles para homogenizar estudios de este tipo a nivel internacional y para la confección de catálogos de cultivares, con gran demanda por parte de productores y especialistas. Para el manejo racional del germoplasma es necesario conocer la variabilidad existente; en este sentido, se recomiendan caracteres morfológicos altamente discriminativos así como marcadores moleculares AFLP y SSR, cuya combinación permite identificar accesiones y formar grupos de diversidad, no solo en guayabo sino también en otros representantes de Myrtaceae. Todo este análisis integral garantiza un proceso de selección más eficiente de genotipos promisorios para diferentes propósitos (mejoramiento, conservación y comercialización). Sobre esta base se recomiendan cruzamientos que pueden servir para la evaluación de híbridos con características comerciales deseables, además del desarrollo de un mapa de ligamiento genético y la asociación a QTLs, como primer paso para una futura selección asistida por marcadores. Se destaca también el papel del cultivo de tejidos, como método alternativo de conservación y para el mejoramiento del guayabo, a través de los principales resultados obtenidos en el país.
Palabras clave: Guayabo, fitomejoramiento, biodiversidad, marcadores moleculares.
Abstract
The objective of this work is to develop a genetic improvement program for guava (Psidium guajava L.) in Cuba, which may contribute to a more efficient use of its genetics resources. It begins with the establishment of the collection, taking into account the prospection of native genotypes, as well as the introduction of wild relatives and accessions with a diverse geographical origin from the main producer countries. These activities are vital for the posterior collection enrichment and maintenance, as well as the different sexual and asexual propagation procedures, which guarantee material multiplication and allow to create new variability sources. Once established, characterization can proceed. For this, a group of minimum descriptors that can be useful to homogenize such studies to an international level and for cultivar catalogues confection are referred to, the latter of great demand from producers and specialists. For a rational germplasm management it is necessary to know the existent variability. In this sense, highly discriminative morphological characters, as well as AFLP and SSR molecular markers, are recommended, whose combination allows to identify accessions and to form diversity groups, not only in guava but also in other Myrtaceae members. All this integral analysis guarantees a more efficient selection process of promissory genotypes for different purposes (breeding, conservation and commercialization). From them, crosses that can serve to evaluate hybrids with desirable commercial characteristics are recommended, as well as the development of a genetic linkage map and QTLs association, as the a step towards a future marker-assisted selection implementation. The role of tissue culture is also stressed, as an alternative conservation method and for guava improvement, considering the main results obtained in the country.
Keywords: Guava plant, breeding, biodiversity, molecular markers.
Introducción
El guayabo (Psidium guajava L.) constituye uno de los frutales tropicales y subtropicales de mayor valor nutricional, que unido a lo rentable de su cultivo, ha posibilitado que adquiera gran importancia económica en varias regiones del mundo (Coelho de Lima et al. 2002, Vasco et al. 2003, Sanabria et al. 2005).
En Cuba no existían plantaciones comerciales de guayaba, a pesar de la amplia variabilidad morfológica de los guayabales silvestres. No es hasta inicios de los años 60 del pasado siglo que se comienzan a desarrollar las primeras plantaciones con genotipos selectos introducidos (Cañizares 1981).
Unido a esto, se inicia en 1965 el establecimiento de un Banco de Germoplasma de Frutales Tropicales y Subtropicales, donde se incluyeron plantas introducidas prospectadas en diferentes localidades y procedentes de otras instituciones como la Estación Experimental Agronómica de Santiago de las Vegas, el Jardín Botánico Soledad (Cienfuegos) y el vivero de Niquero (Granma), entre otros (Cañizares 1981).
Una vez establecida esta colección incipiente, se enriqueció con materiales de características distintivas (Cañizares 1981) y se comienzan a caracterizar visualmente los mismos, lo que unido a la evaluación de los rendimientos y de algunas características del árbol (González y Sourd 1981), permitió que algunos ellos fueran posteriormente recomendados a escala productiva (González et al. 1995). De forma paralela, se desarrollaron experimentos dirigidos a evaluar la resistencia a estrés biótico y abiótico de un grupo de genotipos (González et al. 1996) así como de compatibilidad injerto/patrón (González y Sourd 1981, 1982). Todos estos trabajos constituyeron resultados preliminares que contribuyeron a sentar las bases para el desarrollo futuro de un programa de mejoramiento genético en el cultivo.
Dada la importancia que actualmente ha adquirido este frutal en Cuba (Farrés et al. 2000) y además del valor fitogenético incalculable y disponible para encontrar genotipos superiores a los que se cultivan actualmente (Lozano et al. 2002), se presenta este trabajo cuyo objetivo es desarrollar un programa de mejoramiento genético del guayabo en Cuba.
Programa de mejoramiento genético del guayabo (Psidium guajava L.)
Establecimiento de la colección de Myrtaceae
La prospección y colecta de materiales autóctonos son vías fundamentales para conservar ex situ la diversidad genética existente en una región o país, por lo que resultó una de las estrategias empleadas durante el establecimiento de la colección de Myrtaceae perteneciente hoy al Instituto de Investigaciones en Fruticultura Tropical (González et al. 1996). Algunas formas de ‘Cotorrera’ con características distintivas como alto contenido de vitamina C y de número de semillas, resistencia a estrés abiótico, las 2 últimas justifican su uso como patrón, así como las accesiones ‘Perú roja’ colectada en la Isla de la Juventud, presentan frutos grandes de forma aperada y con color de corteza verde y ‘Microguayaba’ colectada en San Antonio de los Baños, con alto contenido de vitamina C, bajo número de semillas, baja relación L/H de hoja y fruto, considerada una posible subespecie, constituyen ejemplos de estos materiales (Valdés-Infante et al. 2003).
Teniendo en cuenta además la posición que ocuparon los genotipos de guayabo introducidos al país en los dendrogramas generados por los diferentes marcadores genéticos tales como ‘Seychelles’ (Islas Seychelles, frutos redondeados de textura muy lisa, valores altos de acidez), ‘Indonesia blanca’ (Indonesia, frutos medianos con espesor de la pulpa grueso, bajo número de semillas, puede ser un buen sustituto de la pera en forma de cascos por su sabor y textura), ‘N6’ y ‘Suprema roja’ (frutos con forma homogénea y de buena calidad interna y externa), ambos procedentes de la Florida (Valdés-Infante et al. 2003, Rodríguez et al. 2004, Valdés-Infante 2009), se recomienda incluir accesiones con diferente origen geográfico durante el establecimiento de las colecciones. De hecho, algunas de ellas se emplearon en los primeros trabajos de mejoramiento genético en el país (Cañizares 1968, 1981).
De igual forma, la presencia de las especies afines es de vital importancia en la colección ya sea por el aprovechamiento de sus frutos confitados (Pomarrosa de Malasia: Zyzygium malaccense Merr., y Perr., y la Pitanga: Eugenia uniflora L.), para fabricar licores (Guayabita del Pinar: Psidium salutare (H. B. K.) Berg.), así como por disponer genes de resistencia para estrés biótico y abiótico que pudieran ser incorporados al guayabo. Tal es el caso de Psidium friedrichsthalianum Bert. y Psidium cattleianum Sabine, para los cuales se ha referido cierto grado de tolerancia a nemátodos del género Meloidogyne (Díaz-Silveira 1975, Cuadra y Quincosa 1982, Gonzálvez 1989, Pérez 2000), una de las principales plagas del cultivo (Rossi et al. 2002).
Respecto a estrés abiótico, P. cattleianum y P. guineense Sw. pueden soportar pequeños periodos de sequía o de inundación y se dan bien en suelos pobres (Morton 2000), por lo que representan materiales de interés para el país en las nuevas estrategias actuales de desarrollo agroecológico sostenible. A su vez, estas 2 especies son resistentes a las bajas temperaturas (-5,5ºC) y P. friedrichsthalianum es capaz de prosperar bien en regiones elevadas (Morton 2000), características estas que, aunque no tienen gran importancia para las condiciones de Cuba, pueden ser de gran utilidad para países como México, donde sus principales regiones productoras están establecidas por encima de los 1500 m, o para aquellos de clima templado interesados en cultivar este frutal, contribuyendo de esta forma a ampliar el rango de distribución geográfica de esta especie.
Enriquecimiento, mantenimiento y multiplicación de la colección de Myrtaceae
Debido a la influencia de factores bióticos y abióticos, la colección puede sufrir afectaciones que pueden conllevar a la erosión genética, y por tanto, a la pérdida de parte de la diversidad contenida en la misma. Teniendo en consideración la antigüedad de este cultivo en Cuba (Cañizares 1968) y a la alta variabilidad presente en las formas silvestres de guayabo (Cañizares 1981), deben retomarse de forma continua las prospecciones a lo largo y ancho de la Isla. Lo que permite recomendar la replicación de la colección en otras regiones del país y de esta forma minimizar los riesgos de su pérdida.
Por otra parte, la introducción continua de nuevos materiales reviste gran importancia. En este sentido, países como India y Brasil tienen gran diversidad de formas de guayabo obtenidas a través de muchos años de mejoramiento genético (Subramanyan e Iyer 1993, Silva et al. 2000), y son 2 de las regiones de mayor interés para la introducción de genotipos promisorios en la actualidad. También las colecciones de Venezuela (Aranguren y Fermín 2007, Aranguren et al. 2010a y 2010b) y México (Padilla et al. 2010) muestran gran diversidad, y pueden constituir fuentes importantes para importar nuevos tipos de guayabo.
Respecto a la multiplicación, esta especie puede propagarse por 2 vías fundamentales: sexual y asexual o vegetativa. Se recomienda el uso de la forma sexual o por semillas en el país para la obtención de patrones de formas más rústicas, como es el caso de la ‘Cotorrera’. También para la producción de nuevas formas de patrones o cultivares selectos, debido a la gran variabilidad que se genera, producto de la hibridación que presenta la misma a través de su polinización libre o inducida (Peña et al. 2007).
De igual forma, la reproducción asexual juega un papel importante, ya que permite perpetuar las características agronómicas de las plantas madres y se reduce la fase juvenil o periodo pre productivo.
Existen diferentes métodos, dentro de los que se pueden destacar la injertación, producción de posturas por enraizamiento de esquejes, estacas de brotes enraizados en el tronco, estacas de ramas lignificadas, acodos o magullos aéreos o en tierra y estacas de raíz.
También hay varios tipos de injertos que pueden utilizarse en el guayabo tales como: tangencial con patrón decapitado, de chapa o escudete de vástagos o ramas herbáceas y lignificadas. Para las condiciones de Cuba se recomienda este último ya que ha dado buenos resultados. Unido a esto está el hecho de que posibilita mantener por mucho más tiempo el almacenamiento del material vegetativo de propagación cortado, es de fácil ejecución y garantiza un buen prendimiento, aún cuando el portainjerto o patrón no dispone de suficiente savia y no requiere de condiciones especiales para su realización (Peña et al. 2007). Su uso fundamental está encaminado a la identificación de aquellos patrones que puedan producir efectos enanizantes, resistencia a estrés biótico (nemátodos) o abiótico (salinidad, sequía, pH), o que mejoren las características organolépticas del fruto (Bogantes y Mora 2010).
Sin embargo, la producción de posturas por enraizamiento de esquejes es la forma más utilizada en el país para la multiplicación de cultivares de guayabo, debido a que disminuye el tiempo de obtención de posturas listas para la siembra de 8-14 meses por injerto a 3,5-6,0 meses. A su vez, garantiza un ahorro de recursos financieros, por el hecho de que los envases empleados son de menor costo y necesitan menos sustrato; así como un ahorro de fuerza de trabajo al eliminar las labores de producción de los patrones y las relacionadas con el injerto y educación de los patrones (Peña et al. 2007).
El hecho de establecer, enriquecer, mantener y multiplicar una colección teniendo en consideración todos estos elementos, garantiza contar con una fuente de variabilidad adecuada para su uso racional por futuras generaciones.
Caracterización morfoagronómica del germoplasma de guayabo
Durante la selección de un grupo de descriptores para un cultivo se deben identificar aquellos que tienen algún valor o interés para cualquier disciplina científica relacionada con un banco de germoplasma. Los mismos deben corresponderse con caracteres de alta heredabilidad, alto valor taxonómico, alto valor agronómico, baja complejidad de medición y que posean pequeña variación dentro de las muestras (Engels 1985).
En este sentido, la Unión Internacional para la Protección de Nuevas Variedades de Plantas (UPOV, International Union for the Protection of New Varieties of Plants) estableció en 1987 los lineamientos para el desarrollo de la prueba de Distinción, Homogeneidad y Estabilidad, conocida por varios autores como DUS (Distinctness, Uniformity and Stability) (Mauria 2000).
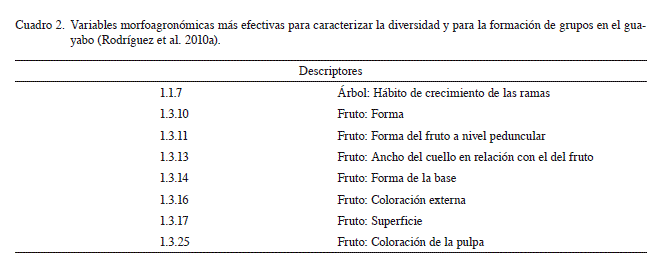
Teniendo en consideración las recomendaciones de la UPOV (UPOV 1987) y en aras de facilitar los estudios de este tipo y de homogenizar los resultados de caracterización y evaluación en la especie, tanto a nivel nacional como internacional, se confeccionó el primer descriptor ilustrado para este frutal en Cuba y en el mundo (Rodríguez et al. 2010a). El mismo representa una herramienta de gran utilidad ya que contiene imágenes que ayudan a la elección de los diferentes estados de los más de 70 caracteres incluidos. Además, se establecieron clases para los caracteres cuantitativos, de forma tal que éstos también puedan ser evaluados como cualitativos, que facilita los trabajos de procesamiento e interpretación de datos. Todos estos elementos justifican la recomendación de su uso para trabajos futuros de caracterización en el cultivo. El mismo incluye los caracteres mínimos referidos en el Cuadro 1 (Rodríguez et al. 2010a).
La evaluación de estos caracteres puede ser de utilidad también en la elaboración de catálogos de cultivares. Los mismos constituyen materiales de consulta de gran valor para productores, investigadores y estudiantes de los diferentes niveles de enseñanza. En estos momentos se cuenta con un catálogo que incluye las 20 accesiones más importantes del banco de germoplasma cubano de guayabo, acompañado de ilustraciones de las mismas, y se trabaja en la preparación de otros genotipos para ser anexados a este material (Rodríguez et al. 2007).
Caracterización de la diversidad del germoplasma de guayabo
Los estudios de variabilidad genética resultan útiles para el manejo racional del material, tanto para su conservación como para el mejoramiento (Marcucci et al. 2001, Frankham et al. 2003). Los mismos permiten definir patrones de variabilidad dentro de la colección, grupos de accesiones con características comunes, caracteres más significativos, entre otros elementos, que posibilitan ordenar la información obtenida y establecer las líneas de acción a desarrollar (Badenes et al. 2000, Tapia et al. 2005).
Existe gran variedad de métodos para determinar la clasificación de las accesiones, los niveles de diversidad y las relaciones entre los individuos (Marcucci et al. 2001, Oliveira y Nascimento 2001). Basado en las experiencias alcanzadas en Cuba para el guayabo (Valdés- Infante 2009), y aunado a las tendencias actuales de complementar los métodos clásicos para evaluar diversidad con otros marcadores genéticos, se recomienda combinar tanto caracteres morfoagronómicos como marcadores moleculares para estos fines. Sin embargo, antes de realizar el estudio, resulta útil conocer las potencialidades de cada marcador, con el objetivo de determinar su aplicación más adecuada, para lo cual resultan de gran ayuda los índices relacionados con el nivel de polimorfismo, poder de discriminación e informatividad (Belaj et al. 2003, Rodríguez et al. 2009). En este sentido, se refiere la efectividad de los microsatélites mPgCIR07, mPgCIR09, mPgCIR11, mPgCIR15, desarrollados por Risterucci et al. (2005), para estimar la diversidad en la colección (Valdés-Infante 2009), así como de las variables morfoagronómicas sugeridas por Rodríguez et al. (2010a). Las variables anteriormente mencionadas (Cuadro 2) se exponen en el presente documento.
También, el contenido de vitamina C (descriptor 1.3.34), es una variable cuantitativa que muestra gran variabilidad entre accesiones y que puede contribuir a la formación de grupos de diversidad en este cultivo.
Por otra parte, caracteres como: forma del extremo peduncular del fruto (descriptor 1.3.11), color de la corteza (descriptor 1.3.16) y de la pulpa (descriptor 1.3.25) y superficie del fruto (descriptor 1.3.17), mostraron un alto poder de discriminación (Dj), lo que indica que pueden ser empleados en la elaboración de claves. Los mismos, de conjunto con el marcador molecular AFLP, son de gran utilidad para la identificación de accesiones en el cultivo (Valdés-Infante et al. 2010a). Los cebadores AFLP más eficientes para la discriminación, dado por los valores de Dj calculados, son: E32 x M33, E32 x M35, E32 x M32, E33 x M39, E32 x M36, E33 x M32, E32 x M34. Las secuencias de los mismos han sido referidas por Vos et al. (1995).
Tanto para los análisis de diversidad, de discriminación, de esclarecimiento de relaciones de parentesco, como de detección de duplicados, se recomienda el uso de estos marcadores individualmente, antes de proceder al análisis de datos combinados, por la información distintiva que brinda cada uno (Rodríguez et al. 2010b).
De forma adicional, se sugiere aplicar estos estudios a otras especies de Myrtaceae, con la finalidad de esclarecer los problemas de clasificación taxonómica que presentan algunos géneros, como por ejemplo Eugenia, así como trazar estrategias futuras de conservación y mejoramiento. La extensión del empleo de cebadores microsatélites diseñados para el guayabo que amplifiquen, tanto en otras especies de Psidium como de otros géneros, sería de gran utilidad para estos propósitos. Lo discutido anteriormente constituye un ahorro en tiempo y recursos, teniendo en cuenta lo caro y laborioso que resulta el desarrollo de marcadores SSR (Valadez y Günter 2000).
Muchos estudios iniciales con SSRs han sido confinados a un único taxa, o sea, al taxon por el cual fueron desarrollados. La razón de esto parece ser debido a la conocida incapacidad de los cebadores SSRs de amplificar DNA en especies diferentes a la cual ellos fueron diseñados (Valadez y Günter 2000). No obstante, existen referencias en la literatura acerca de la transferibilidad de estas secuencias en otros representantes del mismo género e incluso de la misma familia (Zucchi et al. 2003, dos Santos et al. 2007). La capacidad de usar el mismo cebador SSR en diferentes especies de plantas dependerá de cuán conservado esté el sitio flanqueador del SSR entre las taxas relacionadas y la estabilidad de los SSR en el tiempo (Valdés-Infante y Rodríguez 2003).
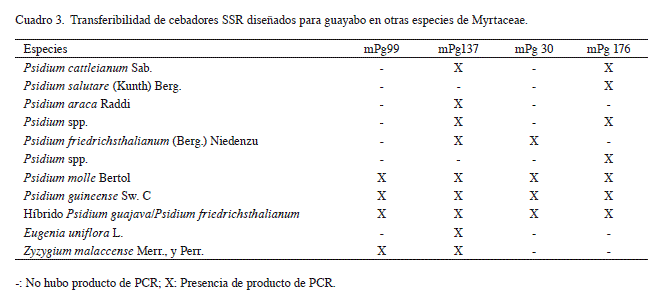
En este sentido, las combinaciones mPg99; mPg137; mPg30 y mPg176, de conjunto con las recomendadas por Risterucci et al. (2005), son marcadores potencialmente útiles para realizar estos estudios en los géneros Psidium, Zyzygium y Eugenia pertenecientes a Myrtaceae, teniendo en cuenta que muestran un 62,5%, un 50% y un 25% de transferibilidad respectivamente (Cuadro 3) (Valdés-Infante et al. 2009).
El número promedio de 13 alelos registrado (Valdés-Infante et al. 2009) supera al de 10,4 encontrado por Zucchi et al. (2003) en la transferencia de cebadores SSR de Eucalyptus spp., en genotipos de Eugenia dysenterica, también pertenecientes a la familia del guayabo. A su vez, las 4 combinaciones mencionadas anteriormente permitieron detectar bandas polimórficas en el 100% de los cebadores evaluados (Valdés-Infante et al. 2009), a diferencia del estudio referido por dos Santos et al. (2007) donde sólo el 26% de las combinaciones analizadas mostraron patrones polimórficos en la amplificación cruzada de cebadores de Eucalyptus spp., con Feijoa sellowiana. De igual forma, Zucchi et al. (2003) sólo pudieron detectar el 2% entre Eucalyptus spp., y Eugenia dysenteria.
La amplificación cruzada con otras especies de Myrtaceae reveló patrones SSR confiables, aunque para el caso de Eugenia uniflora, Psidium spp., procedente de la región Montesano del oriente cubano, Psidium araca y P. salutare, sólo se observó producto de PCR para una de las combinaciones evaluadas (Cuadro 3). Se registraron de 1 a 2 alelos por locus para Zyzygium malaccense, Psidium friedrichsthalianum, Psidium araca, Psidium spp., introducido de Islas Seychelles, Psidium salutare, Psidium spp., procedente de la región Montesano del oriente cubano y Eugenia uniflora (Valdés-Infante et al. 2009).
Por otra parte, Psidium cattleianum, Psidium molle, Psidium guineense y el posible híbrido de Psidium guajava con Psidium friedrichsthalianum mostraron desde 3 hasta 6 alelos por locus (Valdés-Infante et al. 2009). Risterucci et al. (2005) obtuvieron resultados similares para el caso de Psidium cattleianum, lo que coincide con lo referido en la literatura relacionado con el estatus poliploide de esta especie y de Psidium guineense (Nakasone y Paull 1998). No obstante, se hace necesario corroborar estos resultados por hibridación con sondas de estos microsatélites.
El número de alelos comunes detectado en las diferentes especies del género Psidium analizadas corrobora lo planteado por Risterucci et al. (2005) y Valdés-Infante et al. (2009) acerca de la transferibilidad de estos cebadores diseñados para guayabo en otros representantes del género. Marques et al. (2002) y Ochieng et al. (2007) obtuvieron resultados similares para diferentes representantes de Eucalyptus spp., los cuales observaron que estas secuencias SSR están bastante conservadas, lo que permite establecer homologías para trabajos futuros como la integración de mapas de ligamiento y de QTLs, así como estudios de filogenia.
Por otra parte, dos Santos et al. (2007) demostraron la transferibilidad de secuencias SSR entre géneros diferentes (Eucalyptus y Feijoa) pertenecientes a una misma familia (Myrtaceae), cuyo valor fue del 44,5%; a diferencia del referido por Zucchi et al. (2003) que solo obtuvo un 2,8% entre Eucalyptus y Eugenia. De igual forma, Steane et al. (2005) detectaron un 98% de transferibilidad de secuencias de SSR cloroplástico de Eucalyptus globulus en 4 representantes de este género y en otros de la familia Myrtaceae. Esta amplificación cruzada de secuencias conservadas de microsatélites también ha sido referida para otros frutales pertenecientes a los géneros Citrus (Kijas et al. 1995), Prunus (Wünsh y Hormaza 2002) y Malus (Coart et al. 2003).
Otra de las potencialidades de los marcadores SSR es que permiten demostrar que puede existir una correlación entre los resultados obtenidos por mediación de técnicas moleculares y la caracterización morfológica de las variedades (Valdés-Infante y Rodríguez 2003). Las relaciones de parentesco detectadas por el uso de estas combinaciones en los diferentes representantes de Myrtaceae en combinación con accesiones de guayabo de interés pertenecientes a la colección constituyen un reflejo de lo expresado anteriormente (Valdés-Infante et al. 2010b).
En este sentido, resulta interesante destacar la asociación de la accesión ‘Enana Roja Cubana’ con Psidium guineense, sin dejar de lado que es satisfactoria la hibridación entre P. guajava y P. guineense (Landrun et al. 1995), y que los híbridos muestran las mismas características de porte bajo y alta productividad por las que se destaca este cultivar del resto de las accesiones presentes en el país (Valdés-Infante et al. 2010b).
De igual forma, se pudo corroborar el parentesco entre el posible híbrido natural de P. guajava con P. friedrichsthalianum y sus 2 parentales, con la localización de este híbrido en un grupo que incluye ambas especies. El mismo se ubicó más cercano a P. friedrichsthalianum, por lo que se recomienda evaluarlo frente a la infestación con nemátodos del género Meloydogine (Valdés-Infante et al. 2010b).
A su vez, la accesión ‘Microguayaba’ se encontró asociada a una especie del género Zyzygium, con la cual muestra un valor de similitud del 50%. Aún cuando desde un punto de vista morfológico son bien diferentes, se hace necesario ampliar el número de combinaciones de cebadores a evaluar, para lograr una correcta clasificación de este genotipo y determinar si la asociación con este representante es casual o tiene una implicación taxonómica (Valdés-Infante et al. 2010b).
Este enfoque integrado, el cual es el resultado de todo lo comentado en el presente acápite, posibilita ordenar la información obtenida y establecer las líneas de acción necesarias para una conservación más eficaz y un aprovechamiento racional de los recursos genéticos del guayabo y de especies relacionadas.
Recomendación de genotipos para diferentes propósitos
La recomendación de genotipos para diversos fines debe realizarse a partir de la información generada por la caracterización de las accesiones y del estudio de diversidad, lo cual aporta tanto características morfológicas distintivas así como patrones de bandas moleculares que permiten ubicar a las mismas en grupos de diversidad diferentes. Esto garantiza un enfoque más integral, que posibilita una selección más eficiente de accesiones con potencialidades para el mejoramiento genético, así como identificar materiales importantes a conservar por representar una fuente peculiar de variabilidad en la colección. De esta forma se minimiza el riesgo de erosión genética en el cultivo, por el uso excesivo de genotipos estrechamente relacionados (Valdés-Infante 2009).
En este sentido, se destaca la necesidad de conservar las accesiones ‘Darío 19-2’ (bajo número de semillas); ‘Belic L-98’ (pulpa fina, textura lisa de la corteza del fruto); ‘BG 76-23’ (pulpa fina, textura lisa de la corteza del fruto, bajo contenido de vitamina C); ‘Belic L-205’ (bajo contenido de vitamina C y de semillas, forma aperada, hábito de crecimiento disperso del árbol) y ‘Microguayaba’, del banco de germoplasma cubano de guayabo, por presentar alelos raros específicos que las identifican como materiales distintivos. De igual forma, las diferentes especies y géneros de Myrtaceae que se encuentran en la colección cubana (Cuadro 3) mostraron este tipo de alelo. La combinación de alelo raro localizado se considera como una de las más difíciles de encontrar en cualquier estrategia de muestreo de diversidad, lo que realza la importancia de estos genotipos (Valdés-Infante et al. 2009, Valdés-Infante et al. 2010a).
Por otra parte, ‘N6’; ‘Dario 19-2’; ‘ENF 78-7’ (bajo número de semillas y de contenido de vitamina C, altos valores de SST); ‘Suprema roja’; ‘Belic L-100’ (bajo número de semillas y de contenido de vitamina C, altos valores de SST); ‘BG 76-8’ (bajo número de semillas, alto contenido de vitamina C, relación de espesor de la pulpa grueso); ‘BG 76-14’ (alto contenido de vitamina C y bajo de SST, relación de espesor de la pulpa fina); ‘Enana Roja Cubana’ o ‘EEA 18-40’; ‘EEA 1-23’ (porte bajo, productiva, altos valores de SST); ‘Ibarra’ (altos valores de SST, bajo número de semillas, forma redondeada del fruto, textura lisa de la corteza); ‘Microguayaba’; ‘Cotorrera’; ‘Seychelles’; ‘Indonesia blanca’; ‘EEA 6-19’ (relación de espesor de la pulpa grueso, bajo número de semillas, alto contenido de vitamina C); ‘Belic L-207’ (ausencia o muy baja presencia de células pétreas); ‘Belic L-205’; ‘Belic L-120’ (bajos valores de acidez y número de semillas, altos valores de SST), y ‘Perú roja’, representan materiales de interés para futuros trabajos de mejoramiento en el cultivo ya que presentan caracteres agronómicos deseables, así como ubicación diferente en los grupos de diversidad referidos por Rodríguez et al. (2010c) durante el estudio de diversidad de la colección. Esto garantiza una diversidad fenotípica y genotípica adecuada, lo que indica que pueden ser empleados en programas de cruzamientos encaminados a mejorar determinadas características del guayabo para su explotación a escala comercial.
Es importante resaltar también que las accesiones ‘Belic L-205’, ‘Belic L-100’, ‘Suprema Roja’, ‘EEA 18-40’ (‘Enana Roja Cubana’) y ‘N6’ presentan patrón heterocigoto para la mayoría de los loci SSR analizados. Estos resultados sugieren la utilidad de estas accesiones para futuros trabajos de mejoramiento en el cultivo, por ejemplo, para el establecimiento de poblaciones de mapeo, cuya factibilidad está, entre otros elementos, en dependencia de la presencia de altos niveles de heterocigosidad en los parentales escogidos para estos fines (Lanteri y Barcaccia 2005, Rodríguez et al. 2010c).
En contraste, las accesiones ‘BG 76-8’, ‘BG 76-14’, ‘Belic L-122’ (pulpa fina, alto contenido de SST, forma redondeada del fruto, buen contenido de vitamina C), ‘Microguayaba’, ‘ENF 78-7’ y ‘Seychelles’ se muestran homocigotas para todos los loci SSR analizados. Las mismas pueden ser de utilidad en el mejoramiento de este frutal si el objetivo de trabajo está relacionado con el empleo de líneas puras para fijar un determinado carácter.
Desarrollo de cruzamientos para el mejoramiento
La mayoría de los programas de mejoramiento genético en desarrollo están basados en la polinización artificial controlada. En la misma se realizan cruzamientos entre plantas que presentan características de interés con el objetivo de obtener nuevos cultivares. Las plantas que se emplean pueden ser de la misma especie (cruzamiento intraespecífico o intervarietal) o entre especies diferentes (cruzamiento interespecífico) (Pommer y Murakami 2009). Las progenies derivadas de hibridación controlada son de gran utilidad para la obtención de nuevos cultivares, debido a que permiten contar con poblaciones heterogicotas en las cuales hay una adecuada variación genética (Valdés-Infante 2009).
A continuación se refieren un grupo de características, sugeridas por diversos autores, para la búsqueda y selección de genotipos deseables comercialmente: a) frutos de gran tamaño (200-340g), con pocas semillas y pulpa gruesa; b) pulpa blanca para frutos de mesa y rosado intenso para la industrialización (aunque esto puede variar según las demandas de cada país); c) contenido de sólidos solubles totales superiores al 10º Brix; d) acidez de 1,25 hasta 1,50 en aquellos destinados al procesamiento y de 0,2 a 0,6 para el consumo en fresco; f) contenido de vitamina C igual o superior a los 300mp/100g de pulpa; g) número mínimo de células pétreas (responsables de consistencia arenosa de la pulpa); h) buena calidad poscosecha; i) resistencia al daño en los frutos causado por enfermedades e insectos; j) árboles vigorosos, de porte bajo, con hábito de crecimiento disperso; altos productores, resistentes a plagas y enfermedades (Mata y Rodríguez 1985, Pereira y Nachtigal 2002, Pommer y Murakami 2009). Algunos de estos parámetros pueden tener rangos de variación diferente, lo cual está determinado por la demanda del consumo local de cada país. No obstante, de forma general estas son las variables que suelen tenerse en cuenta a nivel internacional en un programa de mejoramiento encaminado a la búsqueda de nuevos cultivares para explotación comercial.
Cruzamientos intraespecíficos
A partir de la selección de progenitores, se debe considerar la variabilidad tanto fenotípica como genotípica, se determinaron programas de cruzamientos orientados a resolver las principales necesidades que tiene este cultivo en la actualidad. Para esto, se recomienda establecer los cruzamientos ‘Enana Roja Cubana’ x ‘N6’, ‘Enana Roja Cubana’ x ‘Suprema roja’ y ‘Enana Roja Cubana’ x ‘Belic L-207’. Para la selección de estos genotipos se tuvo en cuenta que ‘Enana Roja Cubana’ constituye el principal cultivar en explotación comercial y el patrón a mejorar por sus características morfoagronómicas distintivas de porte bajo y altos rendimientos. Respecto a los parentales masculinos, estos presentan forma homogénea y redondeada del fruto (‘N6’, ‘Suprema roja’), buena calidad interna (SST, acidez, contenido de semillas) y poca o ninguna presencia de células pétreas (‘Belic L-207), caracteres de interés a incorporar en el cultivar ‘Enana Roja Cubana’.
Se recomienda, además, realizar otros cruzamientos, como por ejemplo: ‘Enana Roja Cubana’ x ‘Seychelles’ y ‘Enana Roja Cubana’ x ‘Indonesia blanca’, con su origen geográfico diferente (Valdés-Infante 2009) y la presencia de características agronómicas de interés relacionadas con la calidad interna y externa del fruto (SST, acidez, contenido de semillas, forma y textura del fruto, color de la corteza y de la pulpa) (Cañizares 1968, 1981).
Cruzamientos interespecíficos
Las potencialidades de los parientes silvestres en el mejoramiento del guayabo es apreciable; sin embargo, los mismos se han empleado fundamentalmente como portainjerto para regular vigor, calidad del fruto y resistencia a plagas y enfermedades (Pathak y Ojha 1993, Subramanyan e Iyer 1993). De hecho, la hibridación interespecífica no ha brindado buenos resultados en el mejoramiento a escala internacional, debido a la incompatibilidad detectada entre diferentes especies del género Psidium (Subramanyan e Iyer 1993).
Sin embargo, respecto a la resistencia a plagas y enfermedades, es bien conocido que el guayabo es un hospedero de nematodos muy eficiente, ya que Meloidogyne el principal género dañino a este cultivo (Rossi et al. 2002). P. friedrichsthalianum ha sido reportado como una especie con resistencia elevada o moderada a la infestación con nematodos del tipo M. incognita (Díaz-Silveira 1975, González y Sourd 1982, Gonzálvez 1989). El cruzamiento entre P. friedrichsthalianum y P. guajava podría resultar en híbridos resistentes o tolerantes al nematodo, los cuales podrían tener mejor compatibilidad patrón-cultivar que sus parentales, debido a que se ha comprobado que ocurren serias afectaciones en el crecimiento y en el rendimiento de los árboles de P. guajava injertado sobre P. friedrichsthalianum (González y Sourd 1981). De hecho, en las condiciones de Cuba se ha referido un híbrido natural entre ambas especies de Psidium, el cual fue confirmado con el uso de marcadores moleculares SSR (Fernández 1991, Valdés-Infante et al. 2010b).
Por otra parte, P. guineense puede hibridar de forma natural con P. guajava, y los descendientes son generalmente de porte bajo, buenos productores y resistentes, además de que presentan un fruto que se mantiene firme aún después de haber madurado (Landrum et al. 1995); lo que pudiera aumentar la vida postcosecha del guayabo, que es limitada debido a su naturaleza climatérica, así como la obtención de genotipos élites con potencial para su explotación a escala comercial.
Preselección y evaluación de híbridos con características deseables para la comercialización
Comercialización del guayabo
El mercado mundial de la guayaba es aún restringido, sobre todo en comparación con el de otros productos frutales con producción menos dispersa como el mango. Esta circunstancia se refleja en una generalizada carencia de información estadística específica para la guayaba. Las fuentes internacionales, incluyendo la FAO, la consignan cuando lo hacen, en forma agregada con mango y mangostanes. No obstante, en los últimos años se registra un incremento de la producción de guayaba en el mundo (FAO 2002, Pommer y Murakami 2009). Dentro de los principales países productores podemos citar a la India, México, Brasil, Pakistán, Colombia, entre otros (Rodríguez 2007).
Debido a que el fruto del guayabo es altamente perecedero, su comercialización como fruta fresca presenta cierta dificultad en cuanto a su manejo ya que la guayaba madura es muy frágil y se deteriora muy fácilmente por daños mecánicos, sobre maduración, etc., siendo su industrialización la alternativa más viable. Esto, unido a la poca aceptación aroma-sabor en mercados como el europeo y a la falta de divulgación acerca de sus propiedades nutritivas y medicinales, son algunas de las razones por las cuales el aporte de este tipo de comercialización a la exportación es pequeño (ASERCA 1996, Prévez 2007, Rodríguez 2007). Sin embargo, se ha registrado en los últimos tiempos un ligero aumento; aunque la producción destinada al procesamiento agroindustrial aún representa el mayor volumen (Reveles et al. 2003).
Indudablemente el mayor valor nutritivo de la guayaba radica en su alto contenido de ácido ascórbico (vitamina C), alrededor de 300 mg/100g de pulpa. Esta situación hace que la fruta de la guayaba se utilice como aditivo para preparar otros jugos y purés a fin de fortificar su contenido de vitamina. Según los fabricantes, la guayaba rosada es la de más alta calidad por su contenido de azúcares y alto rendimiento en la producción de derivados. Dentro de los productos que se podrían elaborar industrialmente están la pulpa aséptica, pulpa estabilizada químicamente, pulpa refrigerada, concentrado congelado, mermelada, jalea, jugo, néctar, trozos en almíbar, pastas de guayaba, fruta deshidratada, entre otros (Prévez 2007).
El jugo de guayaba se exporta en forma de puré y puré concentrado. Los países exportadores utilizan diferentes variedades, pero por lo general se puede decir que el Brasil exporta jugo de guayaba roja, Malasia y Sudáfrica rosada, la India y Egipto blanca. El puré de guayaba contiene de 8° a 11° Brix y el puré concentrado 20° Brix. El jugo de guayaba y el puré se utilizan en la preparación de jugos con frutas múltiples y néctares y también existe demanda en la industria de alimentos para bebés (Rodríguez 2007).
El mercado mundial de puré de guayaba y puré concentrado es aproximadamente de 6000 toneladas métricas, importadas principalmente por importadores de América del Norte, Europa y el Medio Oriente. Los mercados europeos y norteamericanos por lo general prefieren la guayaba rosada sin mucha semilla. Los mercados árabes tienen preferencia por la guayaba blanca. La situación de precios para el puré de guayaba a 9°Brix ha sido durante los últimos años, alrededor de US$ 400–US$ 650 por tonelada métrica, FOB Brasil, y entre UD$ 700-800 tonelada métrica CIF puerto Europeo (Rotterdam). Adicionalmente, existe una demanda creciente a nivel internacional de frutas deshidratadas. El suministro abundante de frutas podría ser aprovechado para crear nuevos productos para este mercado, incluyendo cueros de fruta orgánica y frutas confitadas (Pommer y Murakami 2009, Prévez 2007).
En Cuba, el guayabo constituye un frutal de alta demanda dentro de la población, dedicándose grandes extensiones de tierra con vistas al consumo en fresco y a la industrialización. Resulta valioso no sólo por su demanda interna; sino también por sus potencialidades de exportación (Farres et al. 2000). Referente a la producción, en el país existe un déficit de cultivares comerciales de guayabo que contribuyan a diversificar los productos que se derivan de sus frutos, por lo que la búsqueda de nuevos individuos con potencialidades para la comercialización constituye una tarea de primer orden.
Las áreas existentes del cultivo de guayaba en Cuba representan el 11% del área total de frutales no cítricos, con 7745 ha plantadas. Las producciones están en el orden de las 3103 TM de guayaba. Los rendimientos se han visto favorecidos a partir de la siembra de variedades con alto potencial productivo como ‘Enana roja cubana’, con rendimientos promedios en todo el país de alrededor de 15 t.ha-1, pero en muchos lugares se han obtenido superiores a los 70 t.ha-1 con producción continua durante todo el año y 2 picos de producción localizados de julio-agosto y oct-nov (Rodríguez 2007).
El comportamiento de la producción en el país en el sector privado y el estatal se manifiesta de la siguiente manera: 33% para el sector estatal y 67% para el privado. A su vez, los rendimientos son de 8,8 t.ha-1 en el estatal y 24,1 t.ha-1 para el privado; mientras que para la siembra de nuevas plantaciones es de 51% para el privado y de 49% para el estatal. En todos los casos el sector privado alcanza mayor eficiencia. De forma general, en los últimos años se observa una tendencia al aumento de la producción y los rendimientos a partir de la aplicación de nuevas tecnologías y la siembra de variedades altamente productivas (Rodríguez 2007).
Aunque la piña continua en el primer lugar del comercio internacional de los frutales tropicales, se ha observado un crecimiento significativo tanto en el valor como en el volumen de otras frutas en los últimos años, particularmente para el caso del mango, y en menor extensión de aguatace, carambola, guayaba, lychee, mangostanes, maracuyá y rambután (FAO 2002, Pommer y Murakami 2009).
Selección de híbridos promisorios
La caracterización y la evaluación han sido las herramientas que usualmente se han empleado en la agricultura para la recomendación de materiales élites, al atender determinadas características importantes económicamente (Persson 2001). El enfoque integral que combina diversidad morfológica y molecular para la recomendación de progenitores a emplear en programas de cruzamientos, garantiza un proceso más eficiente de selección de accesiones promisorias para diversos fines.
Los 25 genotipos de porte bajo y con potencial productivo, preseleccionados de los 354 híbridos obtenidos a partir de los tres cruzamientos desarrollados sobre esta base (‘Enana Roja Cubana’ x ‘N6’, ‘Enana Roja Cubana’ x ‘Suprema roja’ y ‘Enana Roja Cubana’ x ‘Belic L-207’), constituyen un reflejo de la eficiencia de este proceso de selección. Estos híbridos mostraron gran variabilidad en caracteres cualitativos y cuantitativos de interés agronómico, por lo que representan un resultado promisorio para una posterior recomendación a escala productiva.
Muchos de ellos superan a la ‘Enana Roja Cubana’ en un grupo de características de interés comercial a mejorar en este cultivar, como lo son la forma homogénea del fruto, color de la corteza y de la pulpa, número de semillas, SST, vitamina C, acidez, relación de espesor de la pulpa, entre otras. A su vez, heredaron el porte bajo y la productividad de la ‘Enana Roja Cubana’, aspectos que la han mantenido durante todos estos años como el cultivar principal en explotación comercial y el patrón a mejorar por productores y especialistas del cultivo.
Estos resultados son similares a los obtenidos por Du Preez and Welgemoed (1990) en Sudáfrica, los cuales desarrollaron cruzamientos a partir de accesiones con amplia variabilidad en las características físicas y químicas de los frutos. El objetivo fundamental del trabajo estaba encaminado a la búsqueda de cultivares mejorados que garantizaran una mayor estabilidad en la industria, la cual está basada en un solo cultivar que es ‘Fan Retief’. La evaluación de estas progenies permitió detectar grandes diferencias para muchos de los caracteres de interés comentados anteriormente; los cuales, excepto para el contenido de ácido ascórbico, mostraron mejor comportamiento que el cultivar a mejorar.
Otro elemento importante en los programas de mejoramiento de este frutal es el periodo de producción. Esto se debe al hecho de que, debido a las condiciones climáticas, el uso de cultivares con una fecha de entrada en producción que no coincide con el pico normal de cosecha (precoses o tardíos) puede conllevar a que el producto adquiera mejores precios en el mercado de fruta fresca. No obstante, en algunos países se obtiene una producción durante todo el año con un manejo adecuado de las prácticas culturales (Pommer y Murakami 2009).
Tal es el caso de la ‘Enana Roja Cubana’, la cual en altas densidades de plantación presenta un potencial productivo de más de 100 t.ha-1, distribuido de la siguiente forma: 25% julio-agosto; 25% octubre-noviembre y el 50% en los meses restantes (Rodríguez 2007). En los híbridos evaluados se observó diferencias en cuanto a la fecha de entrada en producción, lo que permite hacer un escalonamiento de la cosecha, además de abastecer el mercado con frutas de diversa calidad y satisfacer de esta forma las preferencias de un grupo más amplio de consumidores.
En la actualidad, las instituciones de aquellos países donde la guayaba representa un cultivo de importancia, tales como Brasil, la India, México, Cuba, entre otros, dedican grandes esfuerzos a la obtención y liberación para explotación comercial de nuevas variedades mejoradas (Pommer y Murakami 2009, Valdés-Infante 2009).
Desarrollo del mapa de ligamiento genético y la asociación a QTLs
Muchos de los caracteres de mayor interés agronómico son de naturaleza cuantitativa y su control genético está determinado por múltiples loci con distintos efectos y con complejas interacciones genéticas y ambientales. Por ello se hace necesaria, como herramienta básica de trabajo, la generación de múltiples poblaciones segregantes para los caracteres de interés y la construcción de mapas genéticos (Sunil 1999).
Una aplicación directa de los mapas de ligamiento genético ha sido el marcaje de genes con importancia económica a través de marcadores moleculares, lo cual es conocido como selección asistida por marcadores (MAS, Marker Assisted Selection) (Ochieng et al. 2007).
Caracteres tales como productividad, calidad, maduración y resistencia a estrés biótico y abiótico están controlados por un número relativamente grande de loci, cada uno de los cuales tiene una pequeña contribución positiva o negativa al valor fenotípico final del carácter. Estos loci son llamados “loci de caracteres cuantitativos” (QTLs, Quantitative Trait Loci).
Estas técnicas pueden tener un gran impacto en los programa de mejora de especies frutales; los cuales presentan entre sus principales limitaciones el largo periodo juvenil, que extiende por un buen número de años la evaluación de caracteres importantes del fruto y del desarrollo de la planta (Clegg et al. 2004, Graham et al. 2004, Sanchez-Pérez et al. 2004).
Los 11 grupos detectados en el mapa de ligamiento genético construido, los cuales probablemente sean representativos en el número de cromosomas haploide de la especie, así como los 50 QTLs identificados para caracteres vegetativos y del fruto en el guayabo (Valdés-Infante et al. 2003, Rodríguez et al. 2005, Valdés-Infante 2009), pueden contribuir a la detección y clonaje de los mismos. Esto permite conocer, además, sobre la genética del cultivo, aspecto poco estudiado en la actualidad. Por otra parte, pudieran facilitar los trabajos de mejoramiento a través de la selección en generaciones tempranas de los materiales con características agronómicas de interés, si se reduce el número de genotipos que avanzan en cada ciclo de selección y los tamaños de los experimentos de campo. De esta forma se podría lograr un proceso de selección más eficiente, incluso desde el punto de vista económico (Clegg et al. 2004).
Una vez identificados y caracterizados las regiones del genoma donde se localizan los loci responsables de dichos caracteres, estos genes pueden emplearse en estrategias de ingeniería genética para introducir dichos caracteres en especies heterocigotas y mantener la combinación génica de interés (Georgiady et al. 2001). Por tanto, constituye una herramienta novedosa de gran utilidad para asistir los métodos clásicos de mejoramiento genético, así como sienta las bases para la implementación futura de la selección asistida por marcadores en el cultivo. Se recomienda, además, incluir en el mapa caracteres relacionados con la floración, como una vía de identificar genotipos precoces, medios y tardíos que permitan hacer un escalonamiento de la cosecha y la entrada en el mercado en épocas donde hay poco abastecimiento de fruta y mayores precios.
La ampliación de estos trabajos debe constituir una línea a seguir, que busque además, continuar la saturación del mapa con un mayor número de marcadores genéticos. De hecho, la detección de 117 secuencias candidatas a genes de resistencia (RGL) y de 59 involucradas en el desarrollo de la planta (37 MADS-box y 22 HOMEO-box) (González et al. 2010, Rodríguez et al. 2010c) pueden ser empleados en la saturación del mapa de ligamiento genético reportado para el cultivo. El estudio de las accesiones de la colección cubana de guayabo perteneciente al IIFT permitió identificar fenotipos asociados a posibles mutantes en hoja, flor y fruto para algunos de estos genes, lo cual debe ser corroborado posteriormente mediante la hibridación con sondas individuales y la secuenciación de los productos obtenidos.
Otro tema de vital importancia y que no ha sido explotado en Cuba es la introgresión de genes. El mapa de ligamiento genético y de QTLs construido, además de la detección de secuencias candidatas a genes RGL, MADS-box y HOMEObox, pueden servir de punto de partida para el desarrollo de las investigaciones vinculadas con esta línea de trabajo. En este sentido, se comienzan a dar los primeros pasos a nivel internacional con el clojane del gen CBF para la resistencia al frío en climas templados (Yadav 2008) y la puesta a punto de la metodología para el clonaje del gen que codifica para la enzima hidroperóxidoliasa, vinculada al olor de la fruta de guayaba (Valecillos y Fermín 2010).
Desarrollo de técnicas de cultivo de tejidos para la propagación, conservación y mejoramiento
El cultivo de tejidos tiene 2 papeles importantes en los frutales: facilitar la multiplicación de las plantas y como vía para la creación de nuevas fuentes de variabilidad genética en los cultivares existentes, considerar que la hibridación convencional puede presentar barreras naturales de compatibilidad sexual, lo cual limita su uso (Jiménez 2005, Pascual y Marín 2005, Valdés- Infante 2009).
Estas técnicas pueden constituir herramientas de gran utilidad para la propagación, conservación y mejoramiento del guayabo. Los primeros resultados en Cuba encaminados a lograr la propagación in vitro del guayabo cv., ‘Cotorrera’ fueron reportados por Rodríguez et al. (1994, 1995, 1998). Actualmente, se ha desarrollado una nueva tecnología de propagación in vitro para el cultivar ‘Enana Roja Cubana’ (Pérez et al. 2002, Concepción et al. 2004, 2005a, Concepción 2007). Además, se han realizado algunas investigaciones enfocadas en la embriogénesis somática con embriones cigóticos de este mismo cultivar (Concepción et al. 2005b, Concepción 2007). Estas 2 últimas metodologías son recomendadas para realizar trabajos de este tipo en el cultivo.
Por otra parte, Rodríguez y Velázquez (1999) determinaron los medios de cultivo más eficientes que garantizan una conservación a mediano plazo del guayabo, por un periodo de 6 meses a la temperatura de 21ºC. Esta puede constituir una técnica alternativa a la conservación en campo de genotipos de interés, así como para la introducción de nuevas accesiones al país.
También se ha logrado la propagación de otras especies de Myrtaceae, una de importancia económica como la Guayabita del Pinar (Psidium salutare) para la producción de licores (Sotolongo 2000), y otras endémicas en peligro de extinción, como Eugenia squarrosa Ekman y Urban y Eugenia subdisticha Urban (Montalvo et al. 2010). Todo ello puede contribuir a la conservación del germoplasma a mediano y largo plazo, a la producción de grandes cantidades de material de propagación y a la obtención de nuevas fuentes de variabilidad y de resistencia a estrés biótico y abiótico.
El desarrollo alcanzado en las técnicas de cultivo de tejidos por numerosos países es realmente asombroso, a tal punto, que no se puede pensar en la actualidad en el mejoramiento genético de un cultivo sin recurrir a algunas de sus expresiones, ya sea para acortar el ciclo de mejoramiento como para prevenir el aborto de embriones, vencer obstáculos de incompatibilidad, producir plantas haploides, obtener plantas libres de enfermedades, producir mutantes, obtener poliploides, permitir la clonación de plantas y crear variantes genéticas, entre otras aplicaciones. La implementación de técnicas como cultivo de células, anteras, óvulos y embriones y la fusión de protoplastos en el guayabo y en las especies afines de interés, son algunas de las líneas imprescindibles a desarrollar en el programa de mejoramiento genético del guayabo en Cuba.
Conclusiones
El programa de mejoramiento propuesto puede servir de guía para el desarrollo de este frutal en otras regiones del mundo, y su aplicación contribuirá a un uso más eficiente de los recursos fitogenéticos del guayabo a escala nacional e internacional.
Literatura citada
APOYOS Y SERVICIOS A LA COMERCIALIZACIÓN AGROPECUARIA (ASERCA). 1996. Estudio del Mercado Mundial de la guayaba. Aspra consultores S.C. 4 p. [ Links ]
ARANGUREN Y., BRICEÑO A., FERMÍN G. 2010a. Assessment of the variability of Venezuelan guava landraces by microsatellites. Proceedings of the Second International Guava Symposium, Mexico, Acta Horticulturae. Nº. 849:147-155. [ Links ]
ARANGUREN Y., FERMÍN G. 2007. Variabilidad de guayabas nativas en Venezuela por marcadores moleculares y UPOV. II Simposio Internacional Fruticultura Tropical y Subtropical 2007. ISBN 978-
959-296-001-5. 8 p. [ Links ]
ARANGUREN Y., VALECILLOS C., FERMÍN G. 2010b. Variability of Venezuelan guava geographic landraces employing phenotypic markers. Proceedings of the Second International Guava Symposium, Mexico, Acta Horticulturae. Nº. 849:87-95. [ Links ]
BADENES M.L., MARTÍNEZ- CALVO J., LACER G. 2000. Analysis of a germplasm collection of loquat (Eryobotria japonica Lindl.). Euphytica 114:187-194. [ Links ]
BELAJ A., ZATOVIC Z., CIPRIANI G., BALDONI L., TESTOLIN R., RALLO L., TRUJILLO I. 2003. Comparative study of the discriminating capacity of RAPD, AFLP and SSR markers and their effectiveness in establishing genetic relationship in olive. Theoretical and Applied Genetics.
107:736-744. [ Links ]
BOGANTES A., MORA E. 2010. Evaluación de cuatro patrones para injertos de guayaba (Psidium guajava L.)1. Agronomía Mesoamericana. 21(1):103-111. [ Links ]
CAÑIZARES J. 1968. La guayaba y otras frutas Myrtáceas. Ed. Revolucionaria. Instituto Cubano del Libro. 87 p. [ Links ]
CAÑIZARES J. 1981. Mejoramiento de la guayaba (Psidium guajava L.) por selección masal. Ciencia y Técnica en la Agricultura. 4(3-4):7-23. [ Links ]
CLEGG M., CHENG H., ASHWORTH V. 2004. Application of molecular markers to avocado improvement. Proceedings of the California Avocado Research Symposium. University of California, Riverside. California Avocado Commission, pp. 24-28. [ Links ]
COART E., VEKEMANS X., SMULDERS M.J.M., WAGN ER I., H U Y LEN BROECK J.V., BOCKSTAELE E.V., ROLDÁN-RUIZ I. 2003. Genetic variation in the endangered will apple (Malus sylvestris) in Belgium as revealed by amplified fragment length polymorphism and microsatellite markers. Mol. Ecol. 12:845-857. [ Links ]
COÊHLO DE LIMA M.A., SIMÂO DE ASSIS J., GONZAGA L. 2002. Caracterizacâo dos frutos de goiabeira e selecâo de cultivares na regiâo do submédio Sâo Francisco. Rev. Bras. Frutic. 24(1):273-276. [ Links ]
CONCEPCIÓN O. 2007. Organogénesis y embriogénesis somática in vitro en el guayabo (Psidium guajava L.) cv. Enana Roja Cubana. Tesis de doctorado en Ciencias Agrícolas. Universidad de Ciego de Ávila. Centro de Bioplantas. 114 p. [ Links ]
CONCEPCIÓN O., NÁPOLES L., PÉREZ A.T., HERNÁNDEZ M., PERALTA N., TRUJILLO R. 2005a. Efecto de tres antioxidantes en el cultivo in vitro de ápices de guayaba (Psidium guajava L.). Relación entre el origen del explante y el contenido de compuestos fenólicos. Cultivos Tropicales 26(1):33-39. [ Links ]
CONCEPCIÓN O., NÁPOLES L., PÉREZ A.T., HERNÁNDEZ M., PERALTA N., TRUJILLO R. 2005b. Embriogénesis somática de alta frecuencia en callos de guayaba (Psidium guajava L.) cv. Enana Roja Cubana EEA 18-40, cultivados en medio líquido. Memorias del Congreso Internacional de Biotecnología y Agricultura BIOVEG 2005. Ciego de Avila, Cuba. ISBN: 959-16-0300-2. 10 p. [ Links ]
CONCEPCIÓN O., NÁPOLES L., PÉREZ A.T., PERALTA N., TRUJILLO R. 2004. Regeneración de brotes adventicios en hojas de guayaba (Psidium guajava L.) cultivadas in vitro. Rev. Col. Biotecnología 6:126-135. [ Links ]
CUADRA R., QUINCOSA A. 1982. Comportamiento de diferentes especies de Psidium como patrones para guayabos resistentes a Meloidogyne (Nematoda: Heteroderidae). Ciencias de la Agricultura. 13:19-26. [ Links ]
DIAZ-SILVEIRA M.F. 1975. El Psidium friedrichsthalianum como patrón para guayabo, resistente a los nemátodos del género Meloydogyne. Revista de Agricultura, Habana. 3:80-85. [ Links ]
DOS SANTOS K.L., WELTER L., DE MESQUITA J.A., GUERRA M.P., DUCROQUET J.P.H.J., NODARI R. 2007. Transference of microsatellite markers from Eucalyptus spp., to Acca sellowiana and the successful use of this technique in genetic characterization. Genet. Mol. Biol. 30(1):30-40. [ Links ]
DU PREEZ R.J., WELGEMOED C.P. 1990. Variability in fruit characteristics of five guava selections. Acta Hort. (ISHS) 275:351-360. [ Links ]
ENGELS J. 1985. Caracterización y documentación. Lecturas sobre recursos fitogenéticos 6. Centro Internacional de Recursos Fitogenéticos. 20 p. [ Links ]
FAO. 2002. Commodity Market Review 2001-02. 114 p. [ Links ]
FARRÉS A.E., NODARSE W.G., MULÉN L., PLACERES J., PEÑA O., CASTRO T., DEL VALLÍN G., FRÓMETA E., FRÓMETA E., FUENTES V., GONZÁLEZ G., NORIEGA C., PEDRERA B., PARRA C., SOURD D., BORGES I. 2000. Curso sobre el cultivo de la guayaba. Ed. Instituto de Investigaciones de Cítricos y otros Frutales. 28 p. [ Links ]
FERNÁNDEZ E. 1991. Los nematodos del género Meloidogyne Goeldi en el cultivo de la guayaba (Psidium guajava L.) y su control. Tesis de doctorado en Ciencias Agrícolas, Universidad de la Habana. Cuba. 120 p. [ Links ]
FRANKHAM R., BALLOU J.D., BRISCOE D.A. 2003. Introduction to Conservation Genetics. Cambridge University Press, Cambridge, UK. 100 p. [ Links ]
GEORGIADY M., DESCENZO R., CAIN D.W., IRELAN N. 2001. Linkage mapping and QTL mapping in grape. Plant y Animal Genome Conference IX, San Diego. 15 p. [ Links ]
GONZÁLEZ G., FUENTES V., RODRÍGUEZ N., TORRES M., CAPOTE M., CAÑIZARES J., LIMA H., OROZCO P. 1995. Colecciones y recursos filogenéticos en la Estación Nacional de Frutales de Cuba. Primer Simposio Internacional sobre Fruticultura Tropical y Subtropical, La Habana, Cuba, pp. 71-72. [ Links ]
GONZÁLEZ G., FUENTES V., RODRÍGUEZ N.N., TORRES M., CAPOTE M., CAÑIZARES J., LIMA H., OROZCO P. 1996. Colecciones y recursos fitogenéticos en la Estación Nacional de Frutales de Cuba. Revista del Jardín Botánico Nacional 27-28:123-134. [ Links ]
GONZÁLEZ G., SOURD D. 1981. Ensayo de variedades clonales en Psidium guajava L. Primer Congreso Nacional de Cítricos y Otros Frutales. Palacio de las Convenciones, Ciudad de La Habana, Cuba. Tomo I, pp. 319-336. [ Links ]
GONZÁLEZ G., SOURD D. 1982. Ensayo de tres especies de Psidium y su tolerancia a los nemátodos. Ciencia y Técnica en la Agricultura. Cítricos y otros Frutales 5(2):13-25. [ Links ]
GONZÁLEZ L., BECKER D., RODRÍGUEZ N.N., VALDÉS-INFANTE J., SCHWARZ-SOMMER Z., ROHDE W. 2010. Detection of candidate genes for resistance and plant development in guava (Psidium guajava L.). Proceedings of the Second International Guava Symposium, Mexico, Acta Horticulturae. Nº. 849:241-245. [ Links ]
GONZALVEZ E.F. 1989. Los nemátodos del género Meloidogyne Goeldi en el cultivo de la guayaba y su control. Instituto de Sanidad Vegetal. Ciudad de la Habana. 31 p. [ Links ]
GRAHAM J., SMITH K., MACKENZIE K., JORGENSON L., HACKETT C., POWELL W. 2004. The construction of a genetic linkage map of red raspberry (Rubus idaeus subsp., idaeus) based on AFLPs, genomic-SSR and EST-SSR markers. Theoretical and Applied Genetics 109(4):740-749. [ Links ]
JIMÉNEZ V.M. 2005. Involvement of plant hormones and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regulation 47:91-110. [ Links ]
KIJAS J.M.H., FOWLER J.C.S., THOMAS M.R. 1995. An evaluation of sequence tagged microsatellite site markers for genetic analysis within Citrus and related species. Genome 38:349-355. [ Links ]
LANDRUM L., CLARK W., SHARP W., BRENDECKE J. 1995. Hybridization between Psidium guajava and P. guineense (Myrtaceae). Economic Botany 49(2):153-161. [ Links ]
LANTERI S., BARCACCIA G. 2005. Molecular markers based analysis for crop germplasm preservation. The role of biotechnology, pp. 1-12. [ Links ]
LOZANO J.C., TORO J.C., GARCÍA R., TAFUR R. 2002. Manual sobre el cultivo del guayabo en Colombia. Cali, Colombia. 278 p. [ Links ]
MARCUCCI S., ACUÑA C., TORALES S., ZELENER N., PATHAVER P., LÓPEZ G., HARRAND L., HOPP E. 2001. Evaluación de la Variabilidad Genética en huertos semilleros de especies de Eucalyptus. Consultada el 24 de marzo del 2010. http://www.inta.gov.ar/ediciones/idia/forest/genetica10.pdf 7 p. [ Links ]
MARQUES C.M., BRONDANI R.P.V., GRATTAPAGLIA D., SEDEROFF R. 2002. Conservation and synteny of SSR loci and GTLs for vegetative propagation in four Eucalyptus species. Theoretical and Applied Genetics. 105(2-3):474-478. [ Links ]
MATA I., RODRÍGUEZ A. 1985. Cultivo y producción del guayabo. Ed. Trillas. México. 163 p. [ Links ]
MAURIA S. 2000. DUS testing of crop varieties-a synthesis on the subject for new PVP-opting countries. Plant Varieties and Seeds 13:69-90. [ Links ]
MONTALVO G., QUIALA E., MATOS J., MORFFI H., DE FERIA M., CHÁVEZ M., LA O M., BALBÓN R., PÉREZ M. 2010. In vitro establishment and acclimatization of two threatened species of the genus Eugenia (Myrtaceae). Proceedings of the Second International Guava Symposium, Mexico. Acta Horticulturae. Nº. 849:235-241. [ Links ]
MORTON J.F. 2000. Guava. In: Fruits of warm climates. Miami. Consultada el 24 de marzo del 2010. http://newcrop.hort.purdue.edu/newcrop/morton/guava.html pp. 356-363. [ Links ]
NAKASONE Y.H., PAULL R.E. 1998. Tropical Fruits. Crop Production Science in Horticulture. ISBN 0-85199- 254-4. UK. 468 p. [ Links ]
OCHIENG J.W., STEANE D.A., LADIGES P.Y., BAVERSTOCK P.R., HENRY R.J., SHEPHERD M. 2007. Microsatellites retain phylogenetic signals across genera in Eucalypts (Myrtaceae), Genetics y Molecular Biology. 30(4):1125-1134. [ Links ]
OLIVEIRA M. DO S.P de., NASCIMENTO W.M.O. 2001. Caracterizacao morfológica de frutos em acessos de pupunheira (Bactris gasipaes var. Gasipaes Khunt.). In: Simposio de Recursos Genéticos para América Latina y el Caribe. Instituto Agronómico do Paraná, pp. 348-350. [ Links ]
PADILLA J.S., GONZÁLEZ E., OSUNA E.S. 2010. Collection and characterization of Mexican guava (Psidium guajava L.) germplasm. Proceedings of the Second International Guava Symposium, Mexico, Acta Horticulturae. Nº. 849:49-55. [ Links ]
PASCUAL L., MARIN J.A. 2005. A liquid 2,4-D pulse increased shoot and root regeneration from leaf explants of adult Prunus rootstocks. Scientia Horticulturae 106:582-592. [ Links ]
PATHAK R.K., OJHA C.M. 1993. Genetic Resources of Gueva. In: Advances in Horticulture. Vol. 1-Fruits Crops part 1. Malhotra Publishing House. New Delhi, pp. 143-147. [ Links ]
PEÑA O., GONZÁLEZ D., FARRÉS E., RODRÍGUEZ A., PLACERES J. 2007. Tecnología empleada para la producción de la Guayaba Enana Roja cubana. Curso internacional teórico- práctico en el cultivo del guayabo (Psidium guajava L.) y el papayo (Carica papaya L.). 23 p. [ Links ]
PEREIRA F.M., NACHTIGAL J.C. 2002. Goiabeira, pp. 267-289. In: C.H. Bruckner ed. Viçosa. Melhoramento de Fruteiras Tropicais. UFV. [ Links ]
PÉREZ A.T., NÁPOLES L., CONCEPCIÓN O., TRUJILLO R. 2002. Multiplicación in vitro de brotes de guayabo (Psidium guajava L.) cv. Enana Roja Cubana obtenidos a partir de semillas. Cultivos Tropicales 23(3):57-61. [ Links ]
PÉREZ J.M. 2000. Conferencia sobre fitonematología en el cultivo de la guayaba (Psidium guajava L.). Curso internacional sobre el cultivo de la guayaba. Ed. Instituto de Investigaciones de Cítricos y Otros Frutales. 6 p. [ Links ]
PERSSON H. 2001. Estimating Genetic Variability in Horticultural Crop Species at different Stages of Domestication. Doctoral Thesis, University of Miami, USA. ISSN 1401-6249. ISBN 91-576-5838-2. 30 p. [ Links ]
POMMER C.V., MURAKAMI K.R.N. 2009. Breeding Guava (Psidium guajava L.), pp. 83-120. In: S.M. Jain y P.M. Priyadarshan (eds.). Breeding Plantario Tree Crops: Tropical Species. Springer Science Business Media. Vol 1. [ Links ]
PRÉVEZ L. 2007. Procesamiento de la guayaba. Curso internacional teórico- práctico en el cultivo del guayabo (Psidium guajava L.) y el papayo (Carica papaya L.). 14 p. [ Links ]
REVELES L.R., SAENZ L.A., ESPARZA E., CABRAL F.J. 2003. Polimorfismo de ADN genómico en 12 selecciones de guayabo (Psidium guajava L.) del banco de germoplasma del campo experimental “Los Cañones”. Primer Simposio Internacional de la guayaba, pp. 248-252. [ Links ]
RISTERUCCI A.M., DUVAL M.F., ROHDE W., BILLOTE N. 2005. Isolation and characterization of microsatellite loci from Psidium guajava L. Molecular Ecology Notes 5:824-826. [ Links ]
RODRÍGUEZ A. 2007. El cultivo de la papaya (Carica papaya L.) y de la guayaba (Psidium guajava L.) en el contexto de la fruticultura cubana. Curso internacional teórico- práctico en el cultivo del guayabo (Psidium guajava L.) y el papayo (Carica papaya L.). 18 p. [ Links ]
RODRÍGUEZ N.N., FERMÍN G.A., VALDÉS-INFANTE J., VELÁSQUEZ B., RIVERO D., MARTÍNEZ F., RODRÍGUEZ J., ROHDE W. 2010a. Illustrated descriptors for guava (Psidium guajava L.). Proceedings of the Second International Guava Symposium, Mexico, Acta Horticulturae. Nº.849:103-113. [ Links ]
RODRÍGUEZ N.N., FUENTES J.L., COTO O., FUENTES V.R, RAMÍREZ I.M., BECKER D., RODRÍGUEZ I., GONZÁLEZ C., XIQUÉZ X., ROMÁN M.I., VELÁZQUEZ B., ROHDE W. 2009. Agromorphologic traits, isoenzymes and DNA markers for estimating the polymorphism levels, discriminating capacity and informativeness in avocado. Revista CENIC Ciencias Biológicas 40(1):63-74. [ Links ]
RODRÍGUEZ N.N., FUENTES V.R., HERNÁNDEZ M.R., VALDÉS-INFANTE J., VELÁZQUEZ J.B., RIVERO D., SOURD D.G., RODRÍGUEZ J.A., GONZÁLEZ G., MARTÍNEZ F. 2007. Catálogo de cultivares de guayabo (Psidium guajava L.) en Cuba. Memorias del II Simposio Internacional de Fruticultura Tropical y Subtropical 2007. ISBN 978-959-296-001-5. 10 p. [ Links ]
RODRÍGUEZ N.N., MAS O., CAPOTE M. 1998. Avances en la propagación in vitro de frutales cultivados bajo condiciones tropicales. III Encuentro Latinoamericano de Biotecnología Vegetal. La Habana, Cuba, pp. 107-108. [ Links ]
RODRÍGUEZ N.N., PÉREZ L.O., BLANCO M. 1994. Avances en la propagación in vitro de Psidium guajava L. VII Jornada Científica INIFAT-MINAG. Santiago de las Vegas, Cuba, pp. 94-95. [ Links ]
RODRÍGUEZ N.N., RODRÍGUEZ O.L., BLANCO M., SILVA J. 1995. Sobre la propagación in vitro de Psidium guajava L. I Simposio Internacional sobre Fruticultura Tropical y Subtropical. La Habana, Cuba, pp. 143-144. [ Links ]
RODRIGUEZ N.N., VALDÉS-INFANTE J., BECKER D., VELÁZQUEZ B., COTO O., RITTER E., ROHDE W. 2004. Morphological, agronomic and molecular characterization of Cuban accessions of guava (Psidium guajava L.). J. Genet. y Breed 58:79-90. [ Links ]
RODRÍGUEZ N.N., VALDÉS-INFANTE J., BECKER D., VELÁZQUEZ B., GONZÁLEZ G., SOURD D., RODRÍGUEZ J., BILLOTTE N., RISTERUCCI A.M., RITTER E., ROHDE W. 2005. Characterization of guava accessions by SSR markers, extension of the molecular linkage map, and mapping of QTLs for vegetative and reproductive characters, pp. 201- 215. In: R.K. Pathak, G. Singh, R. Kishun and E. Chandra ed. Proceedings of the First International Guava Symposium. Lucknow, India. December 5-8, 2005. Louvain: ISHS [Belgique]. International Guava Symposium. 1, 2005-12-05/2005-12-08, Lucknow, Inde. [ Links ]
RODRÍGUEZ N.N., VALDÉS-INFANTE J., GONZÁLEZ G., FUENTES V., CAÑIZARES J. 2010c. Genetic resources of guava (Psidiun guajava L.) in Cuba: Germplasm characterization and breeding. Proceedings of the Second International Guava Symposium, Mexico, Acta Horticulturae. Nº. 849:341-349. [ Links ]
RODRÍGUEZ N.N., VALDÉS-INFANTE J., VELÁSQUEZ B., RIVERO D., MARTÍNEZ F., RISTERUCCI A.M., BILLOTE N., BECKER D., RITTER E., ROHDE W. 2010b. Individual versus combined data set for molecular characterization of Cuban guava (Psidium guajava L.) germplasm. Proceedings of the Second International Guava Symposium, Mexico, Acta Horticulturae. Nº. 849:163-175. [ Links ]
RODRÍGUEZ N.N., VELÁSQUEZ B. 1999. Propagación del guayabo. Informe final de proyecto. Propagación, mejoramiento y conservación de frutales cultivados bajo condiciones tropicales. CITMA. Ciudad de la Habana. 70 p. [ Links ]
ROSSI C.E., FERRAZ L.C.C.B., MONTALDI P.T. 2002. Resistencia de Frutíferas de Clima Subtropical e Temperado a Meloidogyne incognita raca 2 e M. javanica. Arq. Ints. Biol. Sao Paulo 69(2):43-49. [ Links ]
SANABRIA H., GARCÍA M., DÍAZ H.A., MUÑOZ J. 2005. Caracterización morfológica en árboles nativos de guayaba del Valle del Cauca. Acta Agronómica 54(4):1-8. [ Links ]
SANCHEZ-PÉREZ R., DICENTA F., MARTINEZ-GOMEZ P., HOWAD W., ARUS P. 2004. Construction of a linkage map and QTL analysis of agronomic traits in almond using SSR markers. ISHS. Acta Horticulturae 726: IV International Symposium on Pistachios and Almonds. 10 p. [ Links ]
SILVA D., EDUINO M., MENDES C., FERNÁNDEZ M., DE LUCENAS J.J., FERNÁNDEZ F., DA SILVA A.V. 2000. Goiabeira (Psidium guajava L.): cultivo sob condicho Irrigada. 2 ed. Recife. SEBRAE/PE. 40 p. [ Links ]
SOTOLONGO R. 2000. Micropropagación de Psidium salutare (H.B.K) Berg. Tesis de doctorado en Ciencias Forestales. Universidad de Pinar del Río, Cuba 100 p. [ Links ]
STEANE D.A., JONES R.C., VAILLANCOURT R.E. 2005. A set of chloroplast microsatellite primers for Eucalyptus (Myrtaceae). Molecular Ecology Notes. 5(3):538–541. [ Links ]
SUBRAMANYAN M.D., IYER C.P.A. 1993. Improvement of Guava, pp. 337-347. In: K.L. Chadha and O.P. Pareek (ed.). Advances in Horticulture. Fruits Crops. Vol. 1-part 1. Malhotra Publishing House. New Delhi. [ Links ]
SUNIL K.L. 1999. DNA markers in plant improvement: An overview. Biotechnology Advances 17:143-182. [ Links ]
TAPIA E., GUTIÉRREZ M.A., WARBURTON M., SANTACRUZ A., VILLEGAS Á. 2005. Characterization of madarin (Citrus spp.) using morphological and AFLP markers. INCI 30(11):1-23. [ Links ]
UPOV. 1987. Guidelines for the conduct of tests for distinctness, homogeneity and stability. TG/110/3. Geneve. 29 p. [ Links ]
VALADEZ M.E., GüNTER K. 2000. Huellas de AND en genomas de plantas. Teoría y protocolos de Laboratorio. Mundi-Prensa México, S.A de C.V. ISBN 968-7462-22-1. 60 p. [ Links ]
VALDÉS-INFANTE J. 2009. Utilización de caracteres morfoagronómicos y de marcadores de ADN para el desarrollo de una metodología que contribuya al mejoramiento genético del guayabo (Psidium guajava L.) en Cuba. Tesis de doctorado en Ciencias Biológicas. Facultad de Biología. UH. Cuba. 100 p. [ Links ]
VALDÉS-INFANTE J., BECKER D., RODRÍGUEZ N.N., VELÁZQUEZ B., GONZÁLEZ G., SOURD D., RODRÍGUEZ J., RITTER E., ROHDE W. 2003. Molecular characterization of Cuban accessions of guava (Psidium guajava L.), establishment of a first molecular linkage map and mapping of QTLs for vegetative characters. J. Genet., y Breed 57: 349-358. [ Links ]
VALDÉS-INFANTE J., RODRÍGUEZ N.N. 2003. Marcadores moleculares de ADN y su aplicación en frutales tropicales., http// fao.cubasi.cu (Sitio virtual de la FAO). 25 p. [ Links ]
VALDÉS-INFANTE J., RODRÍGUEZ N.N., BAUTISTA M., ORTÍZ M.M., QUIROZ A., SÁNCHEZ L.F., RISTERUCCI A.M., ROHDE W. 2010b. Utilización de microsatélites desarrollados en guayabo (Psidium guajava L.) para el estudio de la diversidad en otros representantes de Myrtaceae. Revista Colombiana de Biotecnología. XII (1):64-76. [ Links ]
VALDÉS-INFANTE J., RODRÍGUEZ N.N., BAUTISTA M., ORTÍZ M.M., QUIROZ A., SÁNCHEZ L.F., RISTERUCCI A.M., ROHDE W. 2009. Amplificación cruzada de secuencias simples repetidas de guayabo (Psidium guajava L.) en otros representantes de Myrtaceae. Revista CITRIFRUT. 26(1):15-21. [ Links ]
VALDÉS-INFANTE J., RODRÍGUEZ N.N., VELÁZQUEZ B., RIVERO D., MARTÍNEZ F., RISTERUCCI A.M., BILLOTTE N., BECKER D., ROHDE W. 2010a. Comparison of the polymorphism level, discriminating capacity and informativeness of morph-agronomic traits and molecular markers in guava (Psidium guajava L.). Proceedings of the Second International Guava Symposium, Mexico. Acta Horticulturae. Nº. 849:121-133. [ Links ]
VALECILLOS C., FERMIN G. 2010. Cloning and sequencing of the hydroxyperoxide lyase gene and genetic transformation of guava. Proceedings of the Second International Guava Symposium, Mexico. Acta Horticulturae. Nº. 849:245-251. [ Links ]
VASCO N.L., PADILLA J.S., TORO J. 2003. Composición nutrimental de la guayaba y sus semillas. Primer Simposio Internacional de la guayaba, pp. 116-123. [ Links ]
VOS P., HOGERS R., BLECKER M., REIJANS M., VAN DE LEE T., HORNES M., FRITJERS A., POT J., PELEMAN J., KUIPER M., ZABEAU M. 1995. AFLP: a new technique for DNA fingerprinting. Nucleid Acids Research 23:4407-4414. [ Links ]
WÜNSCH A., HORMAZA J. I. 2002. Molecular characterization of sweet cherry (Prunus avium L.) genotypes using peach (Prunus persica (L.) Batsch) SSR sequences. Heredity 89: 56-63. [ Links ]
YADAV A.K. 2008. Guava biotechnology: Road toward enhancing plant cold hardiness and nutraceutical values of fruit. Oral presentation. Second International Guava Symposium, Mérida-Aguas Calientes. November 10-18. Mexico. [ Links ]
ZUCCHI M.I., BRONDANI R.P.V., PINHEIRO J.B., CHAVES L.J., COELHO A.S.G., VENCOVSKY R. 2003. Genetic structure and gene flow in Eugenia dysenterica DC. In the Brazilian Cerrado utilizing SSR markers. Genet. Mol. Biol. 26:449-457. [ Links ]
*Correspondencia.
Juliette Valdés-Infante: Autor para correspondencia. Correo electrónico: mejoramiento@iift.cu; Instituto de Investigaciones en Fruticultura Tropical. Ave. 7ma. Nº. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana. Cuba, C.P. 11300.
Narciso Nerdo Rodríguez: Instituto de Investigaciones en Fruticultura Tropical. Ave. 7ma. Nº. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana. Cuba, C.P. 11300.
Josefa Bárbara Velásquez: Instituto de Investigaciones en Fruticultura Tropical. Ave. 7ma. Nº. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana. Cuba, C.P. 11300.
Darío Gaspar Sourd: Instituto de Investigaciones en Fruticultura Tropical. Ave. 7ma. Nº. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana. Cuba, C.P. 11300.
Gonzalo González: Instituto de Investigaciones en Fruticultura Tropical. Ave. 7ma. Nº. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana. Cuba, C.P. 11300.
Julio Alberto Rodríguez: Instituto de Investigaciones en Fruticultura Tropical. Ave. 7ma. Nº. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana. Cuba, C.P. 11300.
Wolfgang Rohde: Max-Planck Institut für Züchtungsorschung (MPIZ), Carl-von-Linné-Weg 10, 50829, Köln, Germany.
1 Autor para correspondencia. Correo electrónico: mejoramiento@iift.cu
2 Instituto de Investigaciones en Fruticultura Tropical. Ave. 7ma. Nº. 3005 entre 30 y 32, Miramar, Playa. Ciudad de La Habana. Cuba, C.P. 11300.
3 Max-Planck Institut für Züchtungsorschung (MPIZ), Carl-von-Linné-Weg 10, 50829, Köln, Germany.
Recibido: 10/04/12 Aceptado: 20/08/12

{kind=link}
{kind=link}
{kind=link}