










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkAgronomía Costarricense
versão impressa ISSN 0377-9424
Agron. Costarricense vol.35 no.2 San Pedro de Montes de Oca Dez. 2011
Nota técnica
Influencia de coberturas en suelo sobre la selección del hospedante por Bemisia tabaci, en condiciones de invernadero
Influence of ground covers on hostplant selection by Bemisia tabaci, under greenhouse conditions
*Dirección de correspondencia:
Abstract
Even though several ground covers, either inert or living, can reduce B. tabaci adult numbers arriving to tomato plants, the specific mechanism by which they act remains unknown. Therefore, an experiment was conducted in a greenhouse in Turrialba, Costa Rica, to determine B. tabaci response to both inert (green metal, and silver and yellow plastics) and living covers: perennial peanuts (Arachis pintoi, Fabaceae), “cinquillo” (Drymaria cordata, Caryophyllaceae) and coriander (Coriandrum sativum, Umbelliferae). These were compared to a control (bare soil). There were 4 replicates of each treatment. Each cover was prepared in a large pot, surrounding a 15-cm tall tomato seedling. Pots were distributed on the greenhouse floor, 40 cm apart. Each experiment was replicated over 4 consecutive days, with treatments being randomized each time. A split-plot design was used, with ground covers as the main plot and dates as subplots. On the eve of each replicate, 1200 B. tabaci adults (A biotype) were released into the experimental area, and the next day adults on both the tomato plants and the respective covers were counted. Adult numbers were by far highest on tomato plants sown in bare soil (p<0.01). Furthermore, their numbers did not differ among ground covers, either living or inert (p>0.05). Despite all covers interfering with insect’s ability to locate tomato plants, the underlying specific mechanism differed depending on each cover’s nature, except among living covers, which seemed to share the same mechanism.
Keywords: Ground covers, host-plant selection, Bemisia tabaci, tomatoes, Costa Rica.
Resumen
Aunque varios tipos de coberturas en suelo (inertes o vivas) pueden reducir la afluencia de adultos de B. tabaci en plantas de tomate, se desconoce el mecanismo específico de acción de cada una de ellas. Por tanto, se realizó un experimento en un invernadero en Turrialba, Costa Rica, para determinar su respuesta tanto a coberturas inertes (metal verde y plástico plateado y amarillo) como vivas: maní forrajero (Arachis pintoi, Fabaceae), cinquillo (Drymaria cordata, Caryophyllaceae) y culantro (Coriandrum sativum, Umbelliferae). Se les comparó con un testigo (suelo desnudo). Hubo 4 repeticiones de cada tratamiento. Cada cobertura se sembró en una maceta grande, rodeado de una plántula de tomate de 15 cm de altura. Las macetas se colocaron sobre el piso del invernadero, espaciadas 40 cm entre sí. Cada experimento se repitió en 4 días consecutivos, para lo cual los tratamientos se aleatorizaron todos los días. Se utilizó un diseño de parcelas divididas, con las coberturas como la parcela principal y las fechas como las subparcelas. La víspera de cada repetición se liberaron 1200 adultos de B. tabaci (biotipo A) en el área experimental y al día siguiente se registraron sus datos tanto en la planta de tomate como en cada cobertura. Los números de adultos fueron mucho mayores (p<0,01) en las plantas de tomate del testigo que en las de todos los demás tratamientos. Asimismo, no difirieron entre los diferentes tipos de coberturas (p>0,05). Aunque todas interfieren con la habilidad de los adultos para localizar las plantas de tomate, el mecanismo específico de acción difirió según la naturaleza de cada una, excepto en las coberturas vivas, que parecen compartir el mismo mecanismo.
Palabras clave: Coberturas al suelo, selección del hospedante, Bemisia tabaci, tomate, Costa Rica.
Introducción
Hace casi medio siglo Kennedy et al. (1961) demostraron que algunas especies de áfidos emplean el contraste de color, es decir, las diferentes longitudes de onda emitidas por las plantas y el suelo desnudo, para localizar a sus hospedantes. Sus hallazgos serían confirmados y ampliados en años subsiguientes por otros autores (A’Brook
1968, Smith 1976).
No obstante, se desconocía que este fenómeno también podría ocurrir con otros homópteros, como la mosca blanca B. tabaci, hasta que se evaluaron con éxito varias coberturas inertes, de diferente color, en Israel (Cohen y Melamed-Madjar 1974, 1978, Cohen 1982, Cohen y Berlinger 1986). Más recientemente, gracias a evidencias que sugerían que en semilleros o campos de tomate con crecimiento espontáneo de vegetación silvestre, las plantas de tomate mostraban menor afección por parte de los virus transmitidos por dicho vector; el primer autor se propuso investigar el rol de algunas coberturas vivas en la reducción de la afluencia de adultos de B. tabaci y de la incidencia de los begomovirus (Geminiviridae) comúnmente transmitidos por este insecto.
La experimentación con dichas coberturas, desarrollada durante varios años y resumida por Hilje (2008), incluyó una mezcla de vegetación espontánea (plantas arvenses), las leguminosas maní forrajero (Arachis pintoi) y mucuna (Styzolobium deeringianum) (Fabaceae), el cinquillo (Drymaria cordata, Caryophyllaceae) y el culantro (Coriandrum sativum, Umbelliferae) (Amador y Hilje 1993, Blanco y Hilje 1995, Cubillo et al. 1999a, Hilje y Stansly 2008). Entre todas, sobresalió la última, tanto por su sencillo manejo agronómico como por el aporte de ingresos complementarios para los agricultores.
No obstante, permanece desconocido el mecanismo mediante el que actúan las coberturas vivas, lo cual ocurre también con varios tipos de coberturas inertes que han demostrado su eficacia en reducir la afluencia de adultos de B. tabaci y en la disminución de la incidencia de virosis en tomate (Cohen y Melamed-Madjar 1974, 1978, Cohen 1982, Cohen y Berlinger 1986). Por tanto, de manera paralela a un amplio experimento de campo (Hilje y Stansly 2008), la presente investigación se orientó a tratar de esclarecer el o los mecanismos implicados en la colonización del tomate por parte de los adultos de B. tabaci.
Materiales y Métodos
El experimento se efectuó en un invernadero de la Unidad de Fitoprotección, en el
Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), en Turrialba. El CATIE se ubica en la vertiente del Caribe de Costa Rica, a 09º52’N, 83º38’O y 590 m de altitud.
Se utilizó la var. Hayslip (Asgrow Seed Co., Michigan), y las plántulas se produjeron según procedimientos estándar (Cubillo et al. 1999b), en cartuchos de papel colocados bajo un túnel cubierto con la malla fina IN50 (Tildenet, EE. UU.).
Las plántulas de 22 días, 15 cm de altura y con 4 hojas totalmente desarrolladas se trasplantaron de manera individual en macetas plásticas negras, grandes (10,5 l) (Nº. 1200, V-J Growers Supply, Florida), con suelo esterilizado en una autoclave. En el sustrato se hizo un hueco (6 cm de diámetro), en el centro de cada maceta, para extraer y reemplazar fácilmente las plantas de tomate durante el experimento. Las macetas se colocaron sobre el piso del invernadero, espaciadas 40 cm entre sí (Figura 1).
Se utilizó un diseño completamente aleatorizado, dadas las condiciones homogéneas en el invernadero y el pequeño espacio ocupado por el experimento, con 4 repeticiones. (Little y Hills 1978). Cada experimento se repitió 4 veces, en días consecutivos; aleatorizando los tratamientos. Estos fueron: maní forrajero (MF), cinquillo (CQ), culantro (CO), lámina verde (LV), plástico plateado (PP), plástico amarillo (PA), y suelo desnudo (testigo) (T) (Figura 2).

Las coberturas vivas se establecieron al menos 2 semanas antes de trasplantar las plántulas de tomate, mientras que las coberturas inertes se colocaron la víspera del trasplante; las plásticas se sujetaron a la maceta mediante una banda de hule. La cobertura verde se confeccionó con una lámina de metal delgado, el cual se pintó con una tonalidad semejante a la de una cobertura viva verde, como la Southern Pine, 90 GY 08/187 (The Master Palette, Glidden). Para la cobertura plástica amarilla (Pantone 116C) se utilizó un plástico empleado para fabricar bolsas para basura doméstica (Kanguro). La cobertura plástica plateada (plateado/negro, coextruido, de 1,25 Mls) es un material de uso agrícola (Olefinas S.A., Guatemala). En el tratamiento testigo, la maceta tenía suelo hasta el borde.
Los adultos de B. tabaci (pertenecientes al biotipo A) se recolectaron con un aspirador manual, en una colonia criada sobre plantas de tomate, en un invernadero vecino. Ellos (1200 individuos) se liberaron en el área experimental la víspera de cada experimento, a las 15-16 h, para que se habituaran al sitio. Fueron separados en 6 grupos (de 200 adultos), cada uno colocado en un frasco plástico transparente, el cual se sujetó con una banda de hule a un poste de metal de 1 m de altura. Los 6 postes se distribuyeron de manera uniforme en el área experimental (Figura 1), para evitar sesgos.
Se registraron los números de adultos tanto en el tomate como en cada cobertura, entre 8:30- 10:30 h, antes de su pico de vuelo diario (Jovel et al. 2000), el día de cada experimento. Para lo primero, se colocó una bolsa plástica grande sobre cada planta de tomate (Figura 3a), y ésta se cortó con cuidado (para no perturbar los adultos presentes en la respectiva cobertura), para así capturar los adultos; una vez anestesiados con éter, se contaron en el laboratorio.
Posteriormente, se muestreó cada cobertura mediante una trampa. Esta consistió en un balde negro grande de 34 cm de diámetro, invertido y colocado sobre cada maceta; éste tenía un frasquito en la parte superior y una manga lateral de manta (Figura 3b). Una vez colocado sobre la respectiva cobertura, a través de la manga se introducía la mano del observador para agitar las plantas de la cobertura viva o golpear suavemente la cobertura inerte, de modo que los adultos huyeran y se introdujeran en el frasquito, para ser contados, una vez anestesiados en el laboratorio. Al finalizar cada experimento, se recolectaron indistintos remanente adultos, para evitar sesgos en los recuentos de los experimentos de los días siguientes.
La temperatura y la humedad relativa dentro del invernadero se registraron diariamente, mediante un higrotermógrafo.
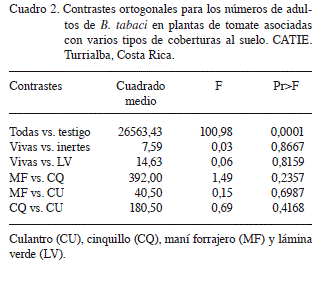
Los datos se sometieron a un análisis de varianza de parcelas divididas (Little y Hills 1978), donde los tratamientos (coberturas) se consideraron como la parcela principal y el tiempo (fechas) como las subparcelas. Las medias fueron comparadas mediante la prueba de Tukey (SAS 1985). Asimismo, se realizaron los siguientes contrastes ortogonales más pertinentes, entre tratamientos individuales o de grupos de tratamientos con ciertas afinidades entre sí (Little y Hills 1978): todas las coberturas vs. testigo, coberturas vivas vs. inertes, coberturas vivas vs. Cobertura verde, MF vs. CQ, MF vs. CU y CQ vs. CU.
Resultados y Discusión
Los números de adultos de B. tabaci fueron mucho mayores (p<0,01) en las plantas de tomate del testigo que en las de todos los demás tratamientos (Cuadro 1). Esto es congruente con todos los hallazgos de campo previos (Amador y Hilje 1993, Blanco y Hilje 1995, Cubillo et al. 1999a, Hilje y Stansly 2008), lo cual indica que aún en áreas muy pequeñas y espacios restringidos, los adultos utilizan el contraste del suelo para seleccionar su planta hospedante, como lo determinaron otros autores para áfidos (Kennedy et al. 1961, A’Brook 1968, Smith 1976).
Otro resultado llamativo es que los números de adultos no difirieron entre los diferentes tipos de coberturas (p>0,05), ya fueran vivas o inertes (Cuadro 2), lo cual sugiere que todas ellas interfieren con la habilidad del insecto para localizar las plantas de tomate.
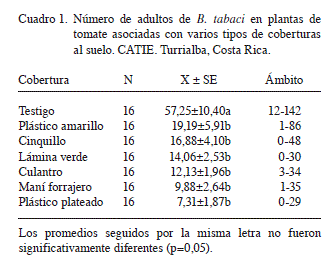
Asimismo, al comparar el número de adultos posados en cada cobertura viva con los posados en la planta de tomate asociada con cada una de ellas, hubo una gran diferencia en el valor promedio consolidado de las 3 coberturas vivas y el de la planta de tomate (2,52 vs. 12,95, p<0,01, N=48). Cabe indicar que en este aspecto las 3 coberturas vivas no difirieron entre sí (p>0,05), con valores promedio de 3,56 (maní forrajero), 2,43 (culantro) y 1,56 (cinquillo); estos datos no se agrupan en un cuadro, pues corresponden a cálculos sencillos.
La posible explicación de este fenómeno es que algunos adultos se refugiaron en las coberturas vivas para evadir las muy altas temperaturas (de hasta 39ºC) típicas de dicho invernadero, pero fueron muy pocos, realmente. Estos datos son bastante congruentes con los de Hilje y Stansly (2008), quienes en el campo registraron números de adultos sumamente bajos en las coberturas vivas, con un valor máximo de apenas 0,33 adultos/ balde, el promedio; además, no se hallaron huevos ni ninfas en ellas. En síntesis, es claro que ninguna de estas coberturas vivas atrae y retiene a los adultos de B. tabaci, lo cual fue aún más evidente en el campo, donde ellos no se estuvieron forzados a permanecer en un espacio pequeño y cerrado, como el invernadero.
En cuanto al mecanismo de acción de cada tipo de cobertura, es claro que difieren entre sí, como se discute a continuación.
En el caso de las 3 coberturas vivas (cinquillo, culantro y maní forrajero) es muy posible que compartan el mismo mecanismo. Al respecto, existen al menos 2 posibilidades. Una es que ellas enmascaren la planta de tomate (de manera visual, olfativa o ambas), tornándola imperceptible para los adultos de B. tabaci y dificultó, o incluso impidió su localización, donde el color es el principal factor determinante en la selección del hospedante, a distancia, pero la capacidad visual de ellos es limitada, por lo que no pueden reconocer a sus verdaderos hospedantes sino una vez que hayan hecho contacto, al insertar su estilete en el apoplasto del mesófilo (van Lenteren y Noldus 1990).
La otra posibilidad es la de la no preferencia. En este caso los adultos aterrizan en el área con vegetación, pero sin distinguir a priori su composición botánica y, una vez que insertan su estilete y se percatan de que la planta no corresponde a la de un hospedante satisfactorio, tienden a alejarse; no obstante, al hacerlo en cierta dirección, algunas quizás se topan con la planta de tomate sembrada en el centro de la maceta y permanecen en ésta.
Esta hipótesis coincide con la teoría de “aterrizajes apropiados/inapropiados”, postulada por Finch y Collier (2000) a partir de su experiencia con la mosca Delia radicum (Anthomyiidae) en un sistema mixto de repollo con trébol. Esta plantea que durante su proceso de búsqueda y en presencia de una mezcla de plantas, los herbívoros pueden aterrizar tanto en sus hospedantes (apropiados) como en los que no lo son (inapropiados), a la vez que evitan el suelo desnudo. Inicialmente, son las sustancias volátiles emitidas por la planta hospedante las que inducen al insecto a detenerse y aterrizar en el área con vegetación, pero sin detectar ni ubicar exactamente a su hospedante. Puesto que, en proporción por unidad de superficie hay mucho más plantas de la respectiva cobertura que de tomate, aumentaría la probabilidad de que el insecto aterrice en una planta de éstas, tras lo cual entrarían en juego otros estímulos (tanto visuales, como de sustancias no volátiles propias de la planta) para su reconocimiento y colonización.
Extrapolado a las condiciones de campo, este mecanismo explicaría en gran parte lo acontecido cuando las coberturas (cinquillo, culantro y maní forrajero) se evaluaron en parcelas de tomate, en Costa Rica (Amador y Hilje 1993, Blanco y Hilje 1995, Cubillo et al. 1999a, Hilje y Stansly 2008). Es decir, sería de esperar que tras varios intentos fallidos y reiterados (al probar con su estilete una y otra vez en varias plantas inapropiadas) los adultos de B. tabaci posiblemente, tiendan a abandonar la parcela de tomate, lo que quizás se vería favorecido por el pequeño tamaño de las plantas de tomate. No obstante, al crecer éstas, se incrementaría la probabilidad de contacto (olfativo, visual o físico) con ellas, por lo que progresivamente se acumularían adultos ahí. El retraso en todo este proceso conduciría a retardar notablemente el desarrollo de la epidemia viral.
Por su parte, el mecanismo de acción de las coberturas inertes es muy diferente de los anteriores, e incluso entre cada una de ellas.
En el caso de la cobertura verde, que simula ser una cobertura viva, actuaría ya sea por enmascaramiento visual de la planta de tomate, aunque no debe descartarse el olor de la pintura. En todo caso, al ser una estructura dura e imposible de penetrar con el estilete, pronto el insecto se percataría de que no le aporta alimento y se alejaría. Al respecto, en 2 experiencias de campo (Amador y Hilje 1993, Blanco y Hilje 1995) en una, el plástico era verde claro y en la otra era oscuro pero se decoloró rápidamente, por lo que la cantidad de adultos no difirió del testigo (tomate en suelo desnudo), quizás por su cercanía cromática con el amarillo. En realidad, tienen una fuerte preferencia por el verde-amarillento más que por el amarillo (van Lenteren y Noldus 1990).
En cambio, la cobertura de plástico plateado actuó como una superficie reflectora de luz ultravioleta (Csizinszky et al. 1999), la cual ha demostrado ser repelente de los adultos de B. tabaci y otros insectos (Swwuan et al. 1988, Csizinszky et al. 1995, 1997, 1999, Berlinger y Lebiush-Mordechi 1996, Cubillo et al. 1999a, Hilje y Stansly 2008), por lo que en la actualidad se utiliza a escala comercial en numerosos países. En el experimento se observó que repele a los adultos de manera muy eficiente, apenas hacen contacto con ella, lo cual explica que en dicha cobertura se presentaran los números más bajos de adultos en la planta de tomate.
Finalmente, aunque sin diferencias estadísticas, el plástico amarillo fue la cobertura con el mayor número de adultos, lo cual podría explicarse porque los atrae, por ser un color fuertemente preferido por B. tabaci, por lo que normalmente se utiliza para capturar los adultos en trampas (Berlinger y Lebiush-Mordechi 1996), pero una vez atraído el insecto al percatarse de que se trata de una estructura dura, inerte y sin valor nutritivo, pronto se alejaría; de hecho el tiempo de permanencia de los adultos en trampas amarillas es de apenas 2,16 min, en promedio, en el invernadero (Hilje, inédito).
Esto explica que en una experiencia de campo (Amador y Hilje 1993) la cantidad de adultos no difiriera del testigo (suelo desnudo), pues ellos pronto se desplazan hacia las plantas de tomate adyacentes. Esto sugiere que su uso en el campo sería contraproducente, pero en Israel las coberturas amarillas son ampliamente utilizadas en campos de tomate. No obstante, se trata de zonas sumamente calientes, donde el plástico acumula tanto calor que mata a los adultos cuando estos son atraídos y se posan sobre él (Cohen 1982, Cohen y Berlinger 1986). En el sitio donde se efectuó el experimento en Costa Rica la temperatura máxima diaria no superó los 23oC, en tanto que en el Valle del Jordán, Israel, ese valor puede alcanzar cerca de 50oC en la cobertura. En el presente experimento aunque las temperaturas del invernadero fueron altas, (de hasta 39ºC) posiblemente causaron el excesivo calentamiento de la cobertura amarilla y de las otras inertes, la metodología empleada no permitía determinar si hubo mortalidad directa causada por dichas coberturas.
En síntesis, es evidente que la respuesta de los adultos de B. tabaci a los distintos tipos de cobertura difiere según la naturaleza (color, olor, sabor y textura) de cada una de ellas. Pero, independientemente del mecanismo específico de cada una, que es interesante entender en términos científicos y académicos, lo más importante es que todas ellas tienen valor práctico para el manejo de B. tabaci como vector de virus en tomate y otros cultivos, en combinación con otras tácticas de manejo integrado de plagas.
Agradecimientos
A Douglas Cubillo, Guido Sanabria y Arturo Ramírez (CATIE), por su apoyo en el trabajo de campo o en los análisis estadísticos. Al CATIE y al Departamento de Agricultura de los EE. UU. (proyecto Development of crop associations for managing geminiviruses vectored by whiteflies in tomatoes, FG-CR-108, CS-AES-7), por el financiamiento de esta investigación.
Literatura Citada
A’BROOK J. 1968. The effect of plant spacing on the numbers of aphids trapped over the groundnut crop. Ann. Appl. Biol. 61: 289‑294. [ Links ]
AMADOR R., HILJE L. 1993. Efecto de coberturas vivas e inertes sobre la atracción de la mosca blanca, Bemisia tabaci (Gennadius), al tomate. Manejo Integrado de Plagas 29:14-21. [ Links ]
BERLINGER M.J., LEBIUSH-MORDECHI S. 1996. Physical methods for the control of Bemisia, pp. 617-634. In: D. Gerling y R.T. Mayer (eds.). Bemisia 1995: Taxonomy, biology, damage, control and management. Intercept, United Kingdom. [ Links ]
BLANCO J., HILJE L. 1995. Efecto de coberturas al suelo sobre la abundancia de Bemisia tabaci y la incidencia de virosis en tomate. Manejo Integrado de Plagas 35:1‑10. [ Links ]
COHEN S. 1982. Control of whitefly vectors of viruses by colour mulches, pp. 45-56. In: K.F. Harris y K. Maramorosch (eds.). Pathogens, vectors and plant disease: Approaches to control. Academic Press, New York. [ Links ]
COHEN S., BERLINGER M.J. 1986. Transmission and cultural control of whitefly-borne viruses. Agric. Ecosys. Environ. 17:89-97. [ Links ]
COHEN S., MELAMED-MADJAR V. 1974. Prevention of the spread of tomato yellow leaf curl virus transmitted by Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in Israel. Bull. Ent. Res. 64(2):193-197. [ Links ]
COHEN S., MELAMED-MADJAR V. 1978. Prevention by soil mulching of the spread of tomato yellow leaf curl virus transmitted by Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in Israel. Bull. Ent. Res. 68(3):465-470. [ Links ]
CSIZINSZKY A.A., SCHUSTER D.J., KRING J.B. 1995. Color mulches influence yield and insect pest populations in tomatoes. J. Amer. Soc. Hort. Sci. 120(5):778-784. [ Links ]
CSIZINSZKY A.A., SCHUSTER D.J., KRING J.B. 1997. Evaluation of color mulches and oils sprays for yield and for the control of silverleaf whitefly, Bemisia argentifolii (Bellows and Perring) on tomatoes. Crop Protection 16(5):475-481. [ Links ]
CSIZINSZKY A.A., SCHUSTER D.J., POLSTON J.E. 1999. Effects of ultraviolet-reflective mulches on tomato yields and on the silverleaf whitefly. HortScience 34:911-914. [ Links ]
CUBILLO D., SANABRIA G., HILJE L. 1999a. Eficacia de coberturas vivas para el manejo de Bemisia tabaco como vector de geminivirus, en tomate. Manejo Integrado de Plagas 51:10-20. [ Links ]
CUBILLO D., SANABRIA G., HILJE L. 1999b. Evaluación de recipientes y mallas para el manejo de Bemisia tabaci mediante semilleros cubiertos, en tomate. Manejo Integrado de Plagas 51:29-35. [ Links ]
FINCH S., COLLIER R.H. 2000. Host-plant selection by insects -a theory based on ‘appropriate/ inappropriate landings’ by pest insects of cruciferous plants. Entomol. Exp. Appl. 96:91-102. [ Links ]
HILJE L. 2008. En busca de coberturas vivas para el manejo de Bemisia tabaci como vector de virus en tomate. Manejo Integrado de Plagas y Agroecología 79-80:98-107. [ Links ]
HILJE L., STANSLY P.A. 2008. Living ground covers for management of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) and tomato yellow mottle virus (ToYMoV) in Costa Rica. Crop Protection 27(1):10-16. [ Links ]
JOVEL J., KLEINN C., CARTIN V., VALVERDE B., HILJE L. 2000. Movimientos diarios de Bemisia tabaci en parcelas de tomate, en Turrialba, Costa Rica. Manejo Integrado de Plagas 55:49-55. [ Links ]
KENNEDY J.S., BOOTH C.O., KERSHAW W.J.S. 1961. Host finding by aphids in the field. III. Visual attraction. Ann. Appl. Biol. 49:1‑21. [ Links ]
LITTLE T.M., HILLS F.J. 1978. Agricultural experimentation; design and analysis. John Wiley & Sons, New York. SAS. 1985. Guide for personal computers. Version 6 ed. SAS Institute Inc. Cary, North Carolina. [ Links ]
SMITH J.G. 1976. Influence of crop background on aphids and other phytophagous insects on Brussels sprouts. Ann. Appl. Biol. 83:1‑13. [ Links ]
SUWWAN M.A., AKKAWI M., AL-MUSA A.M., MANSOUR A. 1988. Tomato performance and incidence of tomato yellow leaf curl (TYLC) virus as affected by type of mulch. Scientia Horticulturae 37(1/2):39-45. [ Links ]
VAN LENTEREN J.C., NOLDUS L.P.J.J. 1990. Whiteflyplant relationships: Behavioural and ecological aspects, pp. 47-89. In: D. Gerling (ed). Whiteflies: Their bionomics, pest status and management. New Castle, UK. Athenaeum Press. [ Links ]
*Correspondencia: Luko Hilje: Departamento de Agricultura y Agroforestería, Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Turrialba, Costa Rica. Correo electrónico: luko@ice.co.cr
Phil A. Stansly: Southwest Florida Research & Education Center, University of Florida. Immokalee, FL 34142, EE.UU.
1/ Autor para correspondencia. Correo electrónico: luko@ice.co.cr
* Departamento de Agricultura y Agroforestería, Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Turrialba, Costa Rica.
** Southwest Florida Research & Education Center, University of Florida. Immokalee, FL 34142, EE.UU.
Recibido: 26/01/11 Aceptado: 18/08/11

{kind=link}
{kind=link}