











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La clase CHONDRICHTHYES Huxley 1880 constituye el clado hermano de todos los vertebrados con mandíbula, ésta clase se caracteriza por un esqueleto cartilaginoso que se puede calcificar a lo largo de la vida del organismo. La clase comprende la subclase Elasmobranchii que incluye los Batoidea, es decir a las rayas, rayas de espina o látigo, guitarras, mantas, mantas diablo, torpedos o rayas eléctricas y peces sierra, entre otros; en la actualidad se registran unas 560 especies de rayas que es más de la mitad de todos los peces cartilaginosos existentes (Dean et al., 2007).
El Superorden Batomorphii Cappetta 1980 o Batoidea Compagno 1973 es un grupo monofilético (Maisey, 1984, McEachran et al., 1996, Naylor et al., 2005, entre otros), presentes en hábitats tan diversos como ríos y lagos, ambientes desmersales, incluso mares abiertos y a grandes profundidades. Su hábitos alimentarios son complejos y variados, pudiendo ser filtradores, durófagos o aquellos especializados en consumir organismos con exoesqueletos duros; hasta aquellos especializados en cazar presas escondidas en fondos lodosos o arenosos, o presas elusivas.
En las últimas décadas, el conocimiento de los ictiolitos asociados a éste superorden, principalmente dientes orales y dérmicos de las especies actuales ha avanzado mucho y ha permitido realizar determinaciones taxonómicas más congruentes y precisas, siempre teniendo en cuenta el previo y basto conocimiento paleontológico del grupo.
El presente trabajo forma parte de las investigaciones sobre los peces fósiles de la Cuenca Limón Sur que la Sección de Geología del Museo Nacional de Costa Rica ha venido realizando en los últimos 20 años y el objetivo principal es actualizar el estatus taxonómico de los peces fósiles, en particular el de los Batomorphii.
Los Batoidea Compagno 1973 en la Cuenca Limón Sur, solo han sido registrados hasta el momento en la Formación Uscari y en sedimentos restrictos al límite Mioceno Superior - Plioceno Inferior, zonas N17 a N19 de foraminíferos, específicamente en la localidad de Alto Guayacán.
Se puede asegurar que 2 publicaciones fueron claves y relevantes para el conocimiento de los peces fósiles de la Cuenca Limón Sur; curiosamente las dos fueron publicadas en el año de 1999 casi de manera simultánea, ''Bathymetric distribution of Miocene to Pleistocene caribbean teleostean fishes from the coast of Panama and Costa Rica'' y la monografía ''Los seláceos fósiles de la localidad de Alto Guayacán'', la primera por Orangel Aguilera y Dione Rodrigues de Aguilera y la segunda por el primer autor del presente trabajo, ello fue un salto cualitativo y también cuantitativo, para el desarrollo y el conocimiento de la paleontología de vertebrados de Costa Rica y América Central, llegándose a registrar cerca de 65 especies de peces fósiles, tanto cartilaginosos como óseos. Desde entonces el conocimiento de los peces fósiles del sur del istmo se ha ampliado notablemente con los importantes aportes de los trabajos de Catalina Pimiento y Victor Perez en Panamá, entre otros, y de los autores del presente trabajo, sin olvidar el aporte del geólogo Cristian Calvo en Costa Rica (ver Aguilera y Rodrigues de Aguilera, 1999, Laurito, 1999; Laurito et al., 2014, Pimiento et al., 2013).
Contexto geológico
La Cuenca sedimentaria de Limón Sur, se ubica en la microplaca Costa Rica-Panamá, propiamente en el trasarco del Arco de Islas y está limitada al occidente por el arco magmático de la Cordillera de Talamanca y hacia el noroeste por el arco volcánico o Cordillera Volcánica Central, específicamente por el edificio volcánico del Turrialba. Hacia el oriente limita con el Mar Caribe y la cuenca de Colombia y su límite con la cuenca de Limón Norte está dado por un sistema de sobrecorrimientos y altos estructurales, estos últimos producto de la tectónica compresiva del Eoceno (Bottazzi et al., 1994). De allí que las secuencias sedimentarias de la Cuenca Limón Sur, han sido fuertemente afectadas por la tectónica asociada al levantamiento del arco de islas y su acoplamiento con América del Sur (ver fig. 1). Ello ha dado origen a una variedad de ambientes marinos, reflejados en múltiples facies litoestratigráficas y sucesiones faunísticas con valor bioestratigráfico, estas últimas conocidas principalmente a partir de las asociaciones de invertebrados fósiles como ostracoda (Borne et al., 1999), nannoplankton (Bybell, 1999), foraminíferos (Cassel, 1986, Cassel y Sen Gupta, 1989, Pizarro, 1985 y Betancur, 2014), briozoos (Cheetham et al., 1999), corales (Aguilar, 1997, Aguilar y Cortés, 2001, Budd et al., 1996, Cairns 1999) y moluscos (Gabb, 1881a y 1881b, Taylor, 1975, Woodring, 1957, 1973 y 1982 entre otros).
Los vertebrados fósiles asociados a sedimentos marinos siempre estuvieron relegadas a un segundo plano, no solo para la Cuenca Limón Sur, sino para todo el país, prestándosele muy poca atención durante la mayor parte del siglo XX o ninguna durante el siglo XIX. Los peces, en especial los ictiolitos de tiburones y rayas, eran recuperados con cierta regularidad en el campo de manera fortuita, lo que produjo escuetas referencias, a falta de técnicas de recuperación apropiadas, ejemplo de ello son los trabajos de Henningsen, 1966 y Kruckow, 1975.
La Localidad de Alto Guayacán, se ubica en una región donde afloran los sedimentos más jóvenes y septentrionales de la Formación Uscari, mismo que son portadores de gran cantidad de ictiolitos (Laurito, 1999). El nombre de Uscari fue originalmente utilizado por Berry (1921) como ''Uscari Shale''. Olsson (1922) utiliza el nombre de ''Uscari Shale Stage or Formation'', para referirse a ''los sedimentos suaves, principalmente lutitas de color oscuro, poco resistentes a la denudación, originando amplios valles y cuencas interiores'' y menciona que el nombre deriva de la quebrada Uscari en el Valle de Talamanca.
La Formación Uscari está descrita como arcillolitas y limolitas con intercalaciones de areniscas de grano fino, calcilutitas y lentes de conglomerados finos, interpretados como depósitos de plataforma silicoclástica interna y externa (Fernández, 1987; Bottazzi et al., 1994; Bottazzi, 2016). Bottazzi (2016), indica que el espesor de ésta formación es de aproximadamente 1000 m y su contacto infrayacente es transicional con la Formación Senosri y discordante con la Formación Fila de Cal. Es sobreyacida concordantemente por la Formación Río Banano; la localidad tipo es la quebrada Uscari y está bien expuesta en los ríos Banano y Peje, señalando una edad de Mioceno Inferior- Mioceno Superior para ésta formación.
La localidad de Alto Guayacán de donde provienen los fósiles de las rayas, acá analizados, es sobreyacida por la Unidad de lavas alcalinas (β en el mapa geológico, Fig. 1), en la margen sur del río Reventazón; en la localidad de Guayacán, esta unidad está constituida por basaltos con analcima con una edad radiométrica K/Ar de 4,6 Ma (sensuBellon y Tournon, 1978; Alvarado y Gans, 2012).
El material del presente estudio se encuentra depositado en la colección de fósiles y minerales, de la Sección de Geología, en el Museo Nacional de Costa Rica, bajo los acrónimos CFM-0000.
Paleontología
Phylum CHORDATA
Subphylum VERTEBRATA
Superclase GNATHOSTOMATA
Clase CHONDRICHTHYES Huxley 1880
Subclase ELASMOBRANCHII Bonaparte 1838
Corte EUSELACHII Hay 1922
Superorden BATOMORPHII Cappetta 1980
Orden RAJIFORMES Berg 1937
Suborden RHINOBATOIDEI Fowler 1941
Familia RHYNCHOBATIDAE Garman 1913
Género Rhynchobatus Müller y Henle 1837
Rhynchobatus cf. R. pristinus (Probst 1877)
Material examinado: figuras 2.1. CFM-4648, 2.2. CFM-4649, 2.3. CFM-4652, 2.4. CFM-4646, 2.5. CFM-4651.
Procedencia: Alto Guayacán, Fm. Uscari.
Características dentales: dientes con coronas elípticas seudo romboidales en vista oclusal, con ornamentación corrugada y una cresta transversal bien marcada, en vista lateral la corona es inflada y redonda. La cara lingual muestra una úvula relativamente alargada, a veces se observa facetamiento por desgaste en su parte media, y está limitada tanto mesial como distalmente por sendas depresiones alares carentes de ornamentación. La raíz es menos ancha que la corona y se proyecta lingualmente por fuera de la misma, presenta sendos lóbulos alargados ubicados por detrás de la úvula que limitan el canal mediano y contiene cada lóbulo un amplio foramen, además de otros mucho más pequeños. En vista ventral el canal es cilíndrico angosto y presenta un amplio foramen nutricio. En vista lingual los lóbulos radiculares están limitados por dos proyecciones alares.
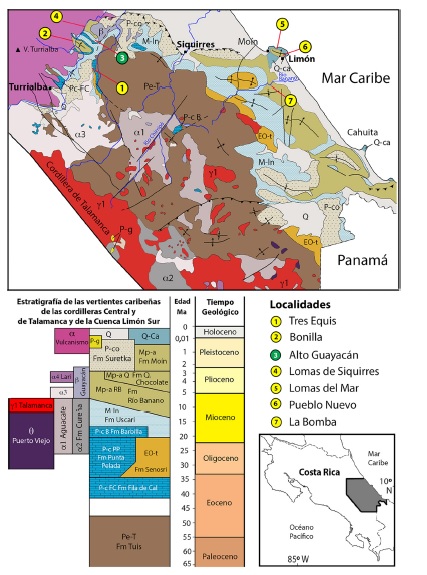
Fig. 1: mapa geológico simplificado que contiene la Cuenca Limón Sur y cuadro de correlación estratigráfico, redibujados de Denyer y Alvarado (2007). Se indican las localidades de 1 a la 7, donde se han registrados restos de peces fósiles en la cuenca, siendo la # 3, la localidad de Alto Guayacán, la única donde hasta el momento se han registrados restos fósiles de peces Batoideos, analizados en el presente escrito. Simbología de las formaciones magmáticas: γ1 plagiogranitos de 87,5 Ma; θ intrusivos gabroicos con diabasas y andesitas de 29 a 11,7 Ma; α1, α2 y α3 vulcanismo del Mioceno y β rocas volcánicas del Plioceno o unidad de lavas alcalinas. Simbología de las formaciones sedimentarias: Pe-T areniscas, lutitas y conglomerados turbidíticos de la Fm. Tuis; P-c rocas calizas de plataformas carbonatadas de las Formaciones Fila de Cal del Eoceno Superior, Punta Pelada de edad Oligoceno al Mioceno Inferior y Barbilla del Mioceno Inferior; EO-t lutitas de la Fm. Senosri de edad Oligoceno - Mioceno Inferior; M-ln limolitas, areniscas y arcillas de plataforma externa de la Fm. Uscari de edad Mioceno Inferior - Plioceno Inferior; Mp areniscas, lutitas y conglomerados asociados a ambientes neríticos a sublitorales de la Fm. Río Banano, Fm. Quebrada Chocolate y Fm. Moín de edad Mioceno Medio a Pleistoceno; P-co conglomerados fluviales de la Fm. Suretka de edad Pleistoceno; Q-ca calizas arrecifales del Plio-Pleistoceno; Q sedimentos cuaternarios de origen fluvial y Pg morrenas y sedimentos de origen glacial.
Rhynchobatus rudolffischeriLaurito 1999
Material examinado: figuras 2.6. CFM-4673, 2.7. CFM-4671, 2.8. CFM-4669 y 2.9. CFM-4668.
Procedencia: Alto Guayacán, Fm. Uscari.
Características dentales: dientes con coronas notablemente bajas de contorno trapezoidal, la cresta transversal divide la corona en dos secciones, la región lingual es lisa y la labial es ornamentada, la úvula es angosta y corta, pero se proyecta y sobresale ligeramente en sentido lingual por sobre las proyecciones del canal mediano de la raíz que es notablemente ancho.
Discusión sobre los registros del género Rhynchobatus: los dientes de Rhynchobatus cf. R. pristinus descritos para la Fm. Uscari son muy similares a los de la especie Rhynchobatus pristinus (Probst 1877) del Cenozoico de Europa, quizás la única diferencia es que la ornamentación de la corona de la especie de la Formación Uscari no es tan marcada. El problema es que la especie R. pristinus presenta una morfología que es más un morfotipo observable en muchas ejemplares del Oligoceno Superior y el Mioceno, lo que hace muy difícil la validación de esta especie fuera de Europa.
Por otra parte, solo tres especies de Rhynchobatus a nivel mundial son reconocidas en el registro fósil, estas son para el Neógeno: Rhynchobatus pristinus (Probst 1877) y Rhynchobatus rudolffischeriLaurito 1999 y para el Paleógeno: Rhynchobatus vincenti Jaekel 1894'' ( cf. Adnet et al., 2020 y Pollerspöck y Straube, 2017).
Distribución paleogeográfica y paleoecología del género Rhynchobatus: aparte del registro de la localidad de Alto Guayacán, en América Central el género Rhynchobatus ha sido registrado en el Mioceno Tardío de la Fm. Gatún en Panamá, asociada a la especie actual Rhynchobatus luebberti (Ehrenbaum 1914) y a una especie indeterminada (Pimiento et al., 2013). Las especies del género Rhynchobatus se asocian a ambientes costeros, como habitantes de los fondos arenosos y lodosos desde la parte superior del talud continental hasta bahías y desembocaduras de ríos, su dieta consiste principalmente de crustáceos y peces (Compagno, 1999).
Familia RHINOBATIDAE Müller y Henle 1838
Género Rhinobatos Linck 1790
Rhinobatos sp. indet.
Material examinado: figuras 3.1. CFM-4679, 3.2. CFM-4681 y 3.3. CFM-4680.
Procedencia: Alto Guayacán, Fm. Uscari.
Características dentales: dientes con coronas bajas con la cara labial corta y convexa, y repliegue basal redondeado; su cara lingual es alargada, con una prominente úvula alargada y cilíndrica, algunos ejemplares muestran facetamiento por desgaste y no se proyecta por delante de la raíz, en algunos casos es ligeramente más corta y se ubica bastante más debajo de la cresta transversal. La corona presenta dos talones a manera de úvulas laterales. La raíz es bilobulada de contorno triangular y de altura similar a la corona, presenta un amplio canal mediano y sendos forámenes a cada lado de la úvula.
Discusión: originalmente estos ejemplares habían sido asociados al género Pristis y sus especies por Laurito (1999), pero se diferencian de Pristis spp. porque en estos últimos la corona es globosa; las longitudes de las regiones labial y lingual de la corona con respecto a la cresta transversal son muy similares, la corona es inflada y más alta que la raíz, y la úvula es cilíndrica.
Distribución paleogeográfica y paleoecología: el registro de Rhinobatos sp. indet. en la localidad de Alto Guayacán, Fm. Uscari, es el único conocido para América Central. Las rayas del género Rhinobatos habitan los fondos lodosos y arenosos de todos los mares tropicales y templados, sobre las plataformas insulares y continentales desde las zonas sublitorales hasta profundidades de 370 m (Compagno, 2005).
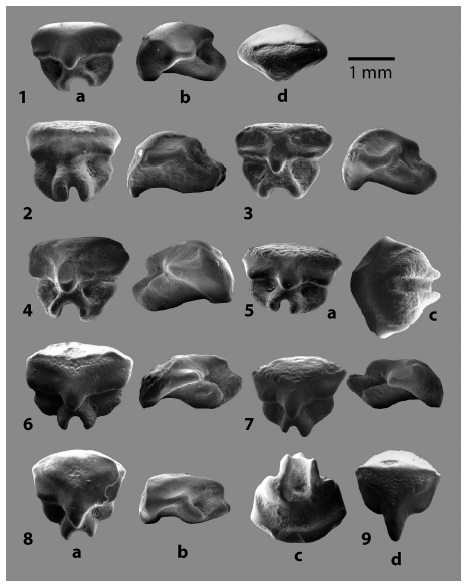
Fig. 2: Familia RHYNCHOBATIDAE Garman 1913, dientes orales de: 1 - 5. Rhynchobatus cf. R. pristinus (Probst 1877) y 6 - 9. Rhynchobatus rudolffischeriLaurito 1999. En vistas a. lingual, b. lateral, c. basal y d. oclusal.
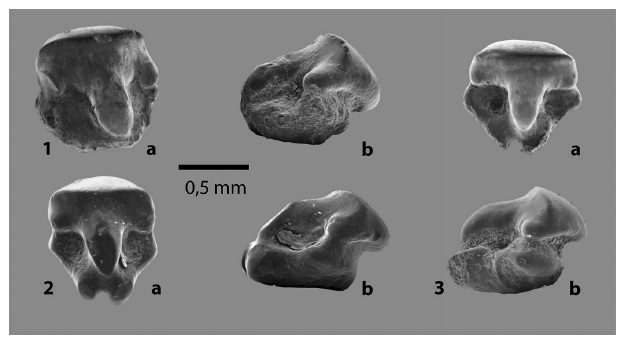
Fig. 3: Familia RHINOBATIDAE Müller y Henle 1838, 1 - 3. dientes orales de Rhinobatos sp. indet. En vistas a. lingual y b. lateral.
Suborden RAJOIDEI Garman1913
Familia ARHYNCHOBATIDAE Fowler 1934
Género Atlantoraja Menni 1972
Atlantoraja sp. indet
Material examinado: figuras: 4.1. CFM-4676 y 4.2. CFM-4677.
Procedencia: Alto Guayacán, Fm. Uscari.
Características dentales: se recuperaron dos dientes, el CFM-4676 es un diente correspondiente a un macho adulto, con la corona alta, cónica y volcada lingualmente con la base subcircular, formando una amplia visera. La corona, además, presenta sendos filos laterales que hacia la base forman un reborde anguloso (fig. 4.1b). La raíz es baja, constituida por dos lóbulos radiculares de similares dimensiones, de contorno elíptico alargado y no tan ensanchados en su parte media, juntándose en el extremo lingual y separándose marcadamente en el extremo labial, conteniendo el canal mediano; los lóbulos radiculares son basalmente planos.
El CFM-4677, un diente de tipo molariforme probablemente corresponde a un macho juvenil, con la corona baja rodeada por un reborde afilado y una ligera protuberancia en la superficie oclusal; la cara lingual presenta una úvula facetada en su extremo superior y la visera se proyecta hacia la raíz, dándole un aspecto triangular (fig. 4.2a), el reborde lingual de la corona es más angosto y de contorno semicircular. La raíz presenta dos lóbulos radiculares de aspecto lobulado y convergen en el extremo lingual y contienen el canal mediano donde se observa el foramen nutricio; en tanto que en el margen labial los lóbulos se proyectan por delante de la corona.
Discusión: el diente CFM- 4676 ilustrado en las figuras 4.1a-c son muy similares a los dientes centrales de los machos adultos de la especie actual Atlantoraja cyclophora Regan1903, aunque la presencia del filo lateral anguloso observable en la base de la corona es una característica que comparte con los dientes de los machos adultos de la especie actual Atlantoraja castelnaui (Miranda Ribeiro 1907), pero no así el formato de la corona tan cónica y redondeada que se observa en ejemplar de la Fm. Uscari. Las anteriores características son comparables con las de los ejemplares ilustrados por Rangel et al., 2016. El CFM-4677, un diente de aspecto molariforme, corresponde probablemente a un juvenil por sus pequeñas dimensiones y la carencia de una cúspide alta (ver figuras 4.2a, 4.2by 4.2d), muy similar a los ilustrados en la lámina 32 por Hovestadt et al., 1996, correspondientes a dientes inferiores de ejemplares juveniles de un macho de Atlantoraja cyclophora Regan1903. Pero en general, también comparte muchos detalles con los dientes inferiores laterales de adultos ilustrados en la lámina 34 por Hovestadt et al., 1996.
Distribución paleogeográfica y paleoecológica: en la actualidad el género Atlantoraja está representado por 3 especies Atlantoraja cyclophora Regan1903, Atlantoraja castelnaui (Miranda Ribeiro 1907) y Atlantoraja platana (Günter 1880) y todas se distribuyen en el Atlántico suroriental en las costas de Brasil, Uruguay y Argentina entre los 20°S - 40°S y a profundidades que varían desde los 24 m a los 150 m (cf. Bernardes et al., 2005). Sin embargo, como fósil el género Atlantoraja ha sido descrito para el Chattiense u Oligoceno tardío de Alemania sensuReinecke (2015), el Oligoceno ''medio'' de Bélgica (ReineckeSteurbaut y Herman, 1978) bajo otros géneros y especies de acuerdo con Reinecke (2015) y el Oligoceno inferior de Ucrania y Oligoceno de México como Raja aff. heinzelini Steurbaut y Herman 1978 de acuerdo con Müller y Rozemberg (2003) y Gonzáles-Barba según cita Marramà et al., 2018.
Por extrapolación y basándonos en los escasos datos biológicos observados para las especies actuales del género (Bernardes et al., 2005 y Oddone y Vooren, 2004), se puede asumir que Atlantoraja sp. indet. de la Formación Uscari, fue una raya desmersal, costero, visitante de la plataforma externa y el talud superior entre los 100 y 300 m, que solía alimentarse de peces, crustáceos y cefalópodos.
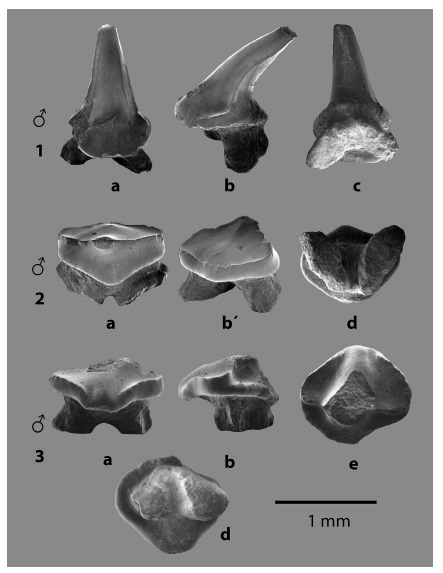
Fig. 4: Familia ARHYNCHOBATIDAE Fowler 1934, 1 - 2. dientes orales de Atlantoraja sp. indet. y Familia RAJIDAE Garman 1913, 3. dientes orales de Dipturus sp. indet. En vistas a. lingual, b. lateral, b. ocluso-labial, c. labio - basal, d. basal y e. oclusal.
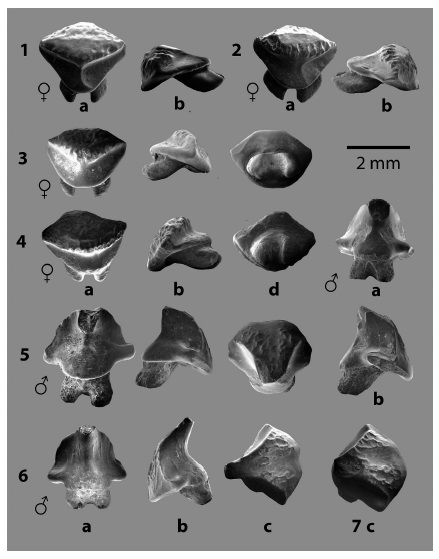
Fig. 5: Familia MYLIOBATIDAE Bonaparte 1838, dientes orales de Dasyatis dianapizarroaeLaurito 1999, 1 - 4. dientes ♀ y 5 - 7. dientes ♂. En vistas: a. lingual, b. lateral, c. oclusal y d. basal.
Familia RAJIDAE Garman 1913
Género Dipturus Rafinesque, 1810
Dipturus sp. indet
Material examinado: figura 4.3 correspondiente al ejemplar CFM-4678.
Procedencia: Alto Guayacán, Fm. Uscari.
Características dentales: se recuperó un único diente lateral de posición incierta, probablemente de un macho adulto. La corona tiene un contorno cuadrangular con los extremos redondeados y los bordes bien marcados y verticales. El ápice de la corona está roto, pero es posible ver que se ubica ligeramente en posición lingual y que el mismo fue corto y de contorno triangular con los extremos redondeados y la cara lingual semicircular. La corona presenta un filo liso y transversal que la divide en un área lingual más corta con pendiente fuerte, reborde angosto y redondeado, en tanto que el área labial es más amplia con un reborde más alto y sigmoideo que hacia el extremo lingual forma una corta visera, angosta e inclinada hacia la raíz; la superficie oclusal de esta visera es angosta y notablemente convexa producto de la proyección lingual del ápice de la corona. El filo hacia los extremos laterales, en vista oclusal (ver fig. 4.3e), es curva y se continua con el filo del reborde labial.
La raíz presenta lóbulos de contorno más o menos triangular, con los extremos ampliamente redondeados, disímiles en cuanto a tamaño y contienen un amplio canal mediano ligeramente inclinado.
Discusión: el diente presenta varias características observadas en algunos dientes laterales inferiores y superiores del género Dipturus, ilustrados por Herman et al., 1995 en las láminas 19 a 21, de allí que se indica que su posición es incierta; entre las características más notorias están el contorno cuadrangular de la corona, el reborde perimetral del filo, la proyección de la corona en una visera corta y volcada hacia abajo, así como el contorno y la asimetría de los lóbulos radiculares. Todas estas características se observan incluso en algunos de los dientes laterales de la especie fósil Dipturus casieri (Steurbaut y Herman 1978) descritos e ilustrados por Reinecke (2015). Sin embargo, el ejemplar CFM-4678 no se puede asociar a ninguna de las cerca de 68 especies actuales, por falta de material comparativo que ilustre las diferencias dentales entre las especies vivientes.
Distribución paleogeográfica y paleoecología: en la actualidad el género Dipturus está constituido por 68 especies de acuerdo con la Fish Base Consortion, con registro de varias especies en el Caribe de Costa Rica, la distribución del género es pantropical y en aguas templadas, con rangos de profundidad variando desde los 20 m a los 1000 m, pero es más frecuente encontrarlas entre los 100 m y los 300 m, por lo tanto se puede asumir una hábito de vida desmersal, habitante de la plataforma continental intermedia hasta el talud superior, depredador de peces, crustáceos y cefalópodos.
Orden MYLIOBATIFORMES Compagno 1973
Superfamilia DASYATOIDEA Whitley 1940
Familia DASYATIDAE Jordan 1888
Género Dasyatis Rafinesque 1810
Dasyatis dianapizarroaeLaurito 1999
Material examinado: figuras 5.1. CFM-4851, 5.2. CFM-4853, 5.3. CFM-4847, 5.4. CFM-4850, 5.5. CFM-4861, 5.6. CFM-4862 y 5.7. CFM-4860.
Procedencia: Alto Guayacán, Fm. Uscari.
Características dentales: los dientes ♀ presentan superficie oclusal de contorno romboide, alargado en sentido mesio-distal y marcadamente angular; en vista lateral la superficie oclusal es bastante plana. La corona está dividida en dos secciones por una cresta transversal apenas insinuada y en algunos casos ausente, el área lingual de la corona está ornamentada por corrugaciones que varían en relieve según el desgaste del diente. El área de la superficie oclusal propiamente dicha abarca un 50% de la superficie total de la corona.
La cara lingual de la corona presenta una cresta lingual mediana, ancha y vertical, con forma triangular hacia la base de la visera y borde basal redondeado. La raíz es bilobulada, sus lóbulos son relativamente anchos y cortos. Los dientes ♂ presentan coronas infladas, cónicas y acampanadas, con cúspides con ápices cilíndricos. La cara labial presenta una leve corrugación como ornamentación, dividida en la parte media por una zona hendida con la superficie relativamente lisa. El visor lingual es notablemente redondeado y está limitado por dos proyecciones alares que caracterizan a la especie.
Discusión: ésta especie se caracteriza por el contorno notablemente acampanado e inflado de la corona de los especímenes machos y por la ornamentación poco conspicua de la corona tanto en machos como en hembras.
Distribución paleogeográfica y paleoecología: Dasyatis dianapizarroaeLaurito 1999 se restringe de momento, al límite Mioceno Superior - Plioceno Inferior de la localidad de Alto Guayacán, Fm. Uscari. Por otra parte, en América Central, el género Dasyatis ha sido descrito para el Mioceno Superior de la Fm. Curré, Localidad de San Gerardo de Limoncito (Laurito y Valerio, 2008) y el Mioceno tardío de la Formación Chucunaque, localidad de Lago Bayano por Perez et al., 2017.
Las rayas del género Dasyatis se caracterizan por ser habitantes epibentónicos de las zonas costeras tropicales y templadas hasta los 200 m de profundidad, suelen alimentarse de peces de fondo, crustáceos y moluscos (McEachran y Capapé, 1984)
Género Taeniurops Garman 1913
Taeniurops sergiomorai sp. nov.
Límite Mioceno Superior - Plioceno Inferior.
Etimología: dedicada al geólogo costarricense, profesor Dr. Sergio Mora Castro por sus importantes aportes a la Geología y a la Ingeniería Geotécnica del país.
Material examinado: figuras correspondientes al Holotipo: 6.1. CFM-4686 ejemplar ♀ y los Paratipos: 6.2. CFM-4701, 6.3. CFM-4684, 6.4. CFM-4704, 6.5. CFM-4682, 6.6. CFM-4683.
Total de ejemplares analizados: 124 bajo los acrónimos consecutivos desde CFM-4682 a CFM-4806..
Procedencia: Alto Guayacán, Fm. Uscari, Provincia de Limón.
Edad: límite Messiniano - Piacenziano.
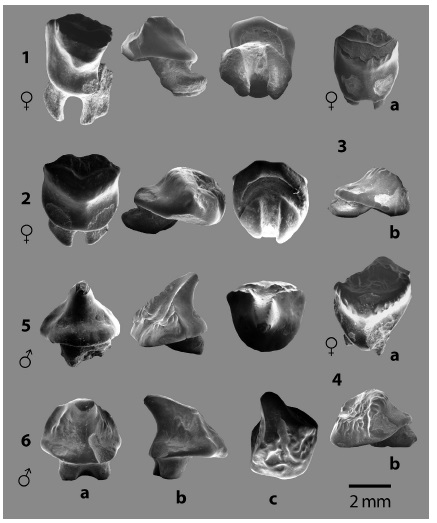
Fig. 6: Familia MYLIOBATIDAE Bonaparte 1838, Taeniurops sergiomorai sp. nov., dientes orales: 1. Holotipo con el código CFM-4686, ♀; Paratipos: 2. CFM-4701 ♀, 3. CFM-4684 ♀, 4. CFM-4682 ♀, 5. ejemplar CFM-4682 ♂ y 6. ejemplar CFM-4683, ♂.
Diagnosis: los dientes ♀ presentan coronas altas de contorno subcircular, con el borde basal constituido por un grueso pliegue redondeado, el borde basal de la margen labial muestra trazos más o menos rectos, angulosos. La superficie oclusal de la corona es alta, abombada y limita la región labial por una cresta transversal poco insinuada, semicircular irregular e incluso en forma de ''V'' muy abierta. La superficie oclusal tiene forma de ''chevron'' amplio y puede mostrar facetamiento por desgaste con proyección de su vértice mediano sobre la parte superior de la visera. La ornamentación de la superficie oclusal, presenta pliegues o corrugaciones cuasi verticales, visibles cuando no hay facetamiento por desgaste, siendo más evidente en la porción superior de la visera. La visera se caracteriza por ser muy amplia, con superficies lisas y está dividida por una amplia y muy poco prominente cresta mediana lingual que en su base puede ser redondeada o presentar una suave hendidura del margen basal de la corona. Es común observar a ambos lados de la cresta mediana, facetamiento de contorno irregular, producido por el desgaste ocasionado por el acoplamiento de los dientes que se ubicaban en la fila posterior, es por ello que también, se puede apreciar las correspondientes superficies de desgaste facetadas en la cara basal del pliegue labial de algunas coronas.
Las superficies labiales de las coronas, se caracterizan por presentar profundas cuencas más o menos ovaladas y alargadas en sentido mesio-comisural, bordeadas por fuertes corrugaciones. La raíz presenta dos lóbulos radiculares alargados y volcados lingualmente con el extremo lingual redondeado, por lo general los lóbulos son proporcionalmente simétricos con un canal central amplio.
Los dientes ♂, presentan coronas de contorno basal lingual notablemente redondeado, siendo el borde basal labial insinuadamente recto; el borde basal lingual, puede mostrar contornos angulosos laterales, pero la región central tiende a ser redondeada. La cúspide de la corona es cónica, lisa y redondeada en su cara lingual, además de curvarse suavemente en sentido lingual; en tanto la cara labial presenta un canal triangular que se expande desde el ápice hacia la base, el canal está limitado por bordes afilados en la mitad superior o pueden estar aplanados y engrosados por facetamiento por desgaste. El canal de la cúspide se ensancha ampliamente en la mitad inferior y está bordeado por amplias y profundas corrugaciones que no llegan a alcanzar el pliegue basal labial de la corona que muestra un contorno redondeado. Los filos que bordean el canal labial de la corona en la mitad inferior, se bifurcan a manera de crestas laterales que dividen la corona en dos secciones. La raíz presenta dos lóbulos muy cortos casi cilíndricos, con un canal mediano angosto.
Discusión: el género Taeniurops Garman 1913 fue resurrecto por Last y Stevens (2009) para la especie Taeniurops meyeni (Müller y Henle 1841), anteriormente incluida en el género Taeniura cf. Cappetta 2012, este último autor incluye también, dentro del género Taeniurops a la especie actual Taeniurops grabata (Geoffroy Saint-Hilaire 1809). En cuanto a la distribución geográfica actual de las especies actuales, T. meyeni es pantropical y se registra en la costa pacífica de nuestro país, no así en el Caribe ni en el Atlántico, la especie T. grabata se registra en las costas atlánticas de África, el Mediterráneo, el Mar Rojo y la costa del Índico africano, en las costas de Kenia, Tanzania, Malawi y Madagascar; así como en las plataformas insulares de Madeiras, Canarias, Cabo Verde y Santa Helena (cf. www.aquamaps.org, version 10/2019. Accessed 4 Dec. 2020), no existiendo registros de T. meyeni y T. grabata en el Caribe, ni Atlántico de América.
Al comparar los dientes ♀ de Taeniurops sergiomorai sp. nov. con dientes ♀ de Taenuriops grabata (Geoffroy Saint-Hilaire 1809), ilustrados por Herman et al., 1998, se ve que comparten muchas características como tener coronas altas, viseras muy amplias, superficie oclusal en forma de ''chevron'', presencia de una cresta mediana lingual, la ornamentación corrugada; pero existen notables diferencias, como: a. el contorno basal perimetral de la corona en la especie nueva es notablemente anguloso y los márgenes tienden a ser más rectos, mientras que en T. grabata tienden a ser curvos y el perímetro es más romboide; b. en T. grabata la superficie oclusal aunque tiene forma de ''chevron'' es relativamente angosta, mientras que en la especie nueva es muy amplia; c. la cresta mediana lingual en la especie actual es más conspícua y menos ancha que en Taeniuropss sergiomorai sp. nov.; d. el contorno de la visera en la especie nueva es cuadrangular mientras que en la especie actual tiende a ser más triangular y e. la ornamentación en la especie actual es mucho más marcada, constituida por corrugaciones del esmalte que se distribuyen más densamente que en la especie fósil.
En relación con la especie fósil Taeniurops cavernosus (Probst, 1877) ampliamente dispersa en el Mioceno del Paratehtys europeo (ver Villafaña et al., 2020, Cappetta, 1970, Antunes et al. 1981, Reinecke et al. 2011), las coronas de machos y hembras de esta especie son más bajas en comparación con el ancho labio-lingual mientras que en la especie nueva de la Formación Uscari las coronas son proporcionalmente más elevadas.
Los dientes de los machos Taeniurops sergiomorai sp. nov., recuerdan a los de los machos del género Dasyatis, pero no muestran ese volumen acampanado, presente en muchas especies de Dasyatis actuales, además, no presentan las proyecciones alares laterales.
Paleobiogeografía y paleoecología: el registro de Taeniurops sergiomorai sp. nov. de momento se restringe al Mioceno Superior - Plioceno Inferior de la Fm. Uscari en el Caribe Sur de Costa Rica. El género Taeniurops, se registra en el Mioceno Tardío de la Fm. Gatún por Pimiento et al., 2013 como Taeniura aff. grabata (Geoffroy Saint-Hilaire 1817). Fuera de América Central, Carrillo et al., 2016 registran cf. Taeniurops sp. en la Fm. Cantaure de edad Mioceno temprano (Burdigaliano a Langhiano). Se asume para Taeniurops sergiomorai sp. nov. una distribución batimétrica costera, desde el sublitoral hasta el talud superior. Pero teniendo el contexto paleoecológico de todo el conjunto de peces de la localidad de Alto Guayacán, tanto los peces óseos como los tiburones y demás rayas con el que la especie está asociada y su relativa alta abundancia, se puede asumir una variación batimétrica entre los 100 y 300 m de profundidad correspondientes con la plataforma externa y el talud superior. Por comparación con la especie actual Taenuriops meyeni (Müller y Henle 1841) se asume que su dieta consistió de peces de fondo, bivalvos, cangrejos y camarones (cf. Compagno et al., 1989).
Superfamilia MYLIOBATOIDEA Compagno 1973
Familia MYLIOBATIDAE Bonaparte 1838
Género Rhinoptera Cuvier 1829
Rhinoptera sp.
Material examinado: ejemplar CFM-4926, figuras 7a-d.
Procedencia: Alto Guayacán, Fm. Uscari, Provincia de Limón.
Características dentales: un fragmento de diente de corona angosta y elevada con la superficie oclusal de aspecto poroso, levemente convexa y contorno rectangular con ligera curvatura en sentido labial. La cara lingual de la corona presenta estriaciones verticales que se proyectan desde el filo superior hasta el cíngulo basal que limita con la parte superior de la raíz, el cíngulo es angosto pero evidente y está constituido por una serie de pequeñas protuberancias. Por debajo del cíngulo, la cara lingual de la raíz presenta poros alineados que se ubican entre las estrías relativamente gruesas que forman parte de las lamelas radiculares basales, y que en el ejemplar están muy erosionadas. La cara labial de la corona es vertical pero los extremos son redondeados, presentando evidente ornamentación por estrías gruesas y verticales en el tercio superior del diente, seguida por una ornamentación más fina compuesta de pequeñas hendiduras a manera de poros en la parte media; la raíz en su límite con la corona presenta forámenes relativamente grandes y alineados.
Discusión: el fragmento dental muestra algún grado de retrabajamiento probablemente por transporte, aunque la ornamentación no muestra un desgaste notable. La corona alta y la raíz muy baja con lamelas angostas y muy continuas permiten determinar que se trata de una especie del género Rhinoptera, aunque Laurito (1999) la asoció a la especie R. studeri, común en el Oligoceno Superior y Mioceno de Europa, en realidad el diente presenta características que son comunes a muchas especies del género, de allí que por lo fragmentario y escaso del material solo se puede concluir que se trata de una especie indeterminada de Rhinoptera sp.
Distribución paleogeográfica y paleoecología: en Costa Rica aparte del registro de la localidad de Alto Guayacán del límite Mioceno Superior- Plioceno Inferior de la Fm. Uscari, el género se ha registrado en el Mioceno Medio de la Fm. Punta Judas (Laurito, 2004) en el Pacífico Central y el Mioceno Superior de la Fm. Curré, en la localidad de San Gerardo de Limoncito en el sur del país (Laurito y Valerio, 2008). En América Central, además de nuestro país, se registra la especie Rhinoptera cf. R. steindachneri (Evermann y Jenkins 1891) en el Mioceno tardío de la Fm. Gatún (Pimiento et al., 2013) y Mioceno Medio cuspidal de la Fm. Alajuela sensuMacFadden et al., 2017, ambos registros de Panamá.
Las especies del género Rhinoptera están constituidos por individuos de hábitos bentopelágicos presentes en las plataformas continentales e insulares internas, en bahías, estuarios y zonas sublitorales. Son migratorios y se alimentan principalmente de invertebrados y moluscos bénticos (Bigelow y Schroeder, 1953).

Fig. 7: Familia MYLIOBATIDAE Bonaparte 1838, fragmento dental de Rhinoptera sp. en vistas: a. oclusal, b. lateral, c. labial y d. lingual.
Superfamilia MOBULOIDEA Whitley 1936
Familia MOBULIDAE Gill 1893
Género Mobula Rafinesque 1810
Mobula thurstoni (Lloyd 1908)
Material examinado: figuras 8.1. CFM-4923, 8.2. CFM-4924, 8.3. CFM-4925, 8.4. CFM-4913 y 8.5. CFM-4880.
Procedencia: Alto Guayacán, Fm. Uscari, Provincia de Limón.
Sinonimia: Mobula lorenzolizanoiLaurito 1999 como sinónimo más reciente sensuAdnet et al., 2012.
Características dentales: dientes ♂ con coronas muy altas e infladas, en vista oclusal las superficies presentan ornamentaciones con rugosidades y diminutos tubérculos; contornos ovalados, romboides o cardiodes, con lóbulos laterales que en algunos casos son muy conspícuos y redondeados; con úvulas centro linguales anchas y a veces son la estructura predominante, ocupando casi toda la superficie oclusal, que suele ser de contorno triangular, otras veces se observan úvulas bífidas angostas y pequeñas protuberancias espiniformes en los extremos lobulares.
Mobula cf. M. japanica (Müller y Henle 1841)
Material examinado: figuras 8.6. (a-c) CFM-4875
Características dentales: diente ♂ de corona muy alta, cilíndrica, ligeramente inflado en forma de clava con el ápice aplanado y volcado lingualmente. La raíz es bilobulada, muy baja y más ancha que la corona, muy similar a los dientes ilustrados en la figura 23 por Notarbartolo-Di-Sciara (1987).
Mobula hypostoma (Bancroft 1831)
Material examinado: figuras 9.1. CFM-4879, 9.2. CFM-4882, 9.3. CFM-4881 y 9.4. CFM-4883.
Características dentales: dientes ♂ con coronas altas, con superficies oclusales con contornos que recuerdan una ''T'', con úvulas centro linguales alargadas y una serie de depresiones y pliegues dispuestos labio-lingualmente, siendo más notorios en el borde labial. Adicionalmente, en cada extremo de la cara lingual en el margen de la superficie oclusal, se observa sendas proyecciones espiniformes, aunque en algunos casos estas proyecciones son redondeadas. La raíz es poliaulacorhizada relativamente baja con 2 o 3 lóbulos.
Discusión sobre el género Mobula
Las especies de Mobula en la localidad de Alto Guayacán, presentan un registro fósil muy particular, ya que todos los ejemplares colectados de Mobula thurstoni (Lloyd 1908), Mobula cf. M. japanica (Müller y Henle 1841) y Mobula hypostoma (Bancroft 1831), que en conjunto suman unos 50 ejemplares, corresponden a ejemplares de machos. Un sesgo en el registro fósil que es difícil de explicar, simplemente apelando a una distribución conductual de los géneros o a fenómenos asociados a procesos tafonómicos o digenéticos. Algo que no se ha visto en otras localidades de América Central Meridional, como en la localidad de Lago Bayano, Formación Chucunaque y en la Formación Gatún, ambas del Mioceno Superior de Panama, donde suelen registrarse tanto machos y hembras de diferentes especies del género Mobula (ver Pimiento et al., 2013 y Perez et al., 2017).
Distribución paleogeográfica del género Mobula
Mobula thurstoni (Lloyd 1908), aparte de la localidad de Alto Guayacán del Mioceno Superior-Plioceno Inferior de la Fm. Uscari en Costa Rica (Laurito, 1999), se registra en el Mioceno Superior de la localidad de Lago Bayano de la Fm. Chucunaque (Perez et al., 2017) y el Mioceno Medio de la Fm. Gatún, ambas en Panamá (Pimiento et al., 2013). Mobula hypostoma (Bancroft 1831), se registra en la Fm. Uscari, en la localidad de Alto Guayacán (Laurito, 1999), y en la Fm. Gatún de edad Mioceno Medio de Panamá (Pimiento et al., 2013).
Género Manta Bancroft 1829
Manta cf. M. birostris (Donndorff, 1798)
Material examinado: figuras 9.5. CFM-4874 y 9.6. CFM-4877.
Procedencia: Alto Guayacán, Fm. Uscari, Provincia de Limón.
Características dentales: el género Manta solo consta de dientes inferiores (sensuHerman et al., 2000), los ejemplares de la Fm. Uscari presentan coronas altas en forma de clava cilíndrica simple o doble, con la superficie oclusal aplanada pero oblicua y ornamentada con corrugaciones y con uno o dos ápices principales, limitados por sendos ápices secundarios volcados hacia la región lingual. La raíz es baja, inflada con dos lóbulos laterales amplios y una especie de protuberancia lingual, angosta pero prominente.
Dicusión: al comparar los ejemplares de la Fm. Uscari con los ejemplares actuales Manta birostris (Donndorff, 1798) ilustrados por Herman et al., 2000, no cabe duda de su similitud pero no son exactamente iguales, las formas fósiles parecen ser más esbeltas en sus proporciones. Por otra parte, se puede establecer con algún grado de seguridad que se trata de dientes de ejemplares ♀, en vista de que son más altos que anchos.
Ecología de los mobúlidos
Los mobúlidos están presentes en los mares tropicales y templados entre los 40º de latitud Norte y 40º de latitud Sur, pero la mayoría de las especies tienen una distribución tropical y subtropical en aguas con temperaturas entre los 20 y 26 ºC. Las diferentes especies de Mobúlidos son consideradas pelágicas o epipelágicas, en aguas costeras, incluso es posible encontrarlas en aguas someras o en estuarios, pero también se pueden encontrar en aguas profundas del mar abierto a 1600 m. Suelen viajar y alimentarse en escuelas con número variable de especímenes, desde unos pocos a varios cientos, alimentándose de zooplancton y pequeños peces, durante el día en las áreas costeras y migrando a las aguas abiertas y profundas por las noches. (Notarbartolo-di-Sciara, 1988, Couturier et al., 2012).
Móbula japanica (Müller y Henle 1841) es un organismo filtrador de plancton, en el Golfo de México su dieta consiste casi exclusivamente de camarones eufausiáceos del género Nyctiphanes que forma parte del krill (Notarbartolo di-Sciara, 1988), por lo que realiza migraciones verticales aprovechando las concentraciones de sus presas, durante el día se suele encontrar a profundidades menores a 5 m en aguas con temperaturas mayores a 25°C pero pueden hacer incursiones hasta los 1400 a 1800 m de profundidad; en general hacen migraciones entre los 50 y 200 m tanto de día como de noche (Francis y Jones, 2017, Croll et al., 2012).
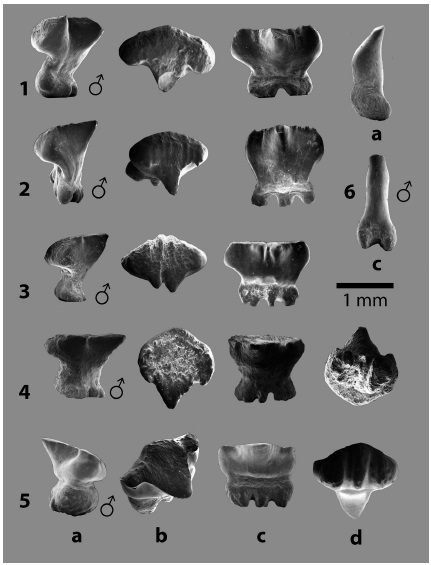
Fig. 8: Familia MOBULIDAE Gill 1893, dientes orales de: 1 - 5. Mobula thurstoni (Lloyd 1908) y 6. Mobula cf. M. japanica (Müller y Henle 1841). En vistas a. lateral, b. oclusal, c. lingual y d. basal.
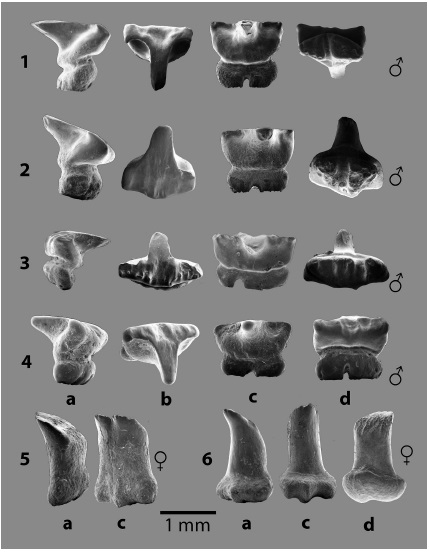
Fig. 9: Familia MOBULIDAE Gill 1893, dientes orales de: 1 - 4. Mobula hypostoma (Bancroft 1831) y 5 - 6. Manta cf. M. birostris (Donndorff, 1798). En vistas a. lateral, b. oclusal, c. lingual y d. basal.
Mobula thurstoni (Lloyd 1908) presenta similares patrones de alimentación y dieta que Móbula japanica (Müller y Henle 1841) según lo observado por Notarbartolo-di-Sciara (1988), básicamente krill de camarones de Nyctiphanes y Mysidium, copépodos, anfípodos, huevos de peces, larvas de estomatópodos y larvas de megalopa, en suma, una dieta muy especializada. No existen investigaciones sobre la distribución batimétrica y migraciones diarias de Mobula thurstoni, por lo general sus registros se restringen a no más de 100 m. De la especie Mobula hypostoma (Bancroft 1831) se sabe que es de habitos costero, pelágico y oceánico, pero no hay datos de su dieta, migraciones diarias o estacionales.
Manta birostris (Donndorff, 1798) es un gigante planctívoro que se alimenta de una variedad de zooplancton presente en las aguas superficiales de los mares tropicales y subtropicales entre los 35°N y 35°S, pero trabajos recientes sugieren que M. birostris prefiere presas de las regiones mesopelágicas (Burgess et al., al., 2016). Para esta especie, se han registrados inmersiones de hasta 600 m, relacionadas con la búsqueda de zooplancton en áreas de surgencia submarinas ricas en nutrientes (Stewart y Aburto-Oropeza, 2015).
Distribuición actual y paleogeográfica de los Mobulidae
Manta birrostris, Mobula japonica y Mobula thurstoni tienen una distibución mundial reportándose en los Océanos Indico, Pacífico y Atlántico, mientras que Mobula hypostoma se restringe al Atlántico de América y todo el Caribe (Couturier et al., 2012).
Los registros fósiles del género Manta son sumamente raros, el más antiguo procede del Oligoceno temprano de la Molassa Suiza (Picot et al., 2008) y el siguiente registro totalmente fiable, procede de la Localidad de Alto Guyacán, Mioceno Superior - Plioceno Inferior de la Fm. Uscari, en la provincia de Limón, el cual fue descrito inicialmente por Laurito (1999) y corroborado por Adnet et al., 2012. Espinas caudales atribuidas al género Manta, han sido descritas por Purdy et al., 2001 para el Plioceno de la Formación Yorktown en Lee Creek Mine en Carolina del Norte.
Ictiolitos asociados
Entre los ictiolitos asociados a Batoideos en la Localidad de Alto Guayacán, Laurito (1999) describió escamas placoides, basicamente 2 dientes dérmicos, uno de forma subcircular con corona central de esmaltoide y un patrón de cóstulas radiales, la raíz es mucho más amplia y alta que la corona. El segundo diente es de contorno elíptico, con la corona oval redondeada y lisa, su raíz es mucho más alta que la corona y presenta costulaciones radiales. No es posible asignarlas a un género específico.
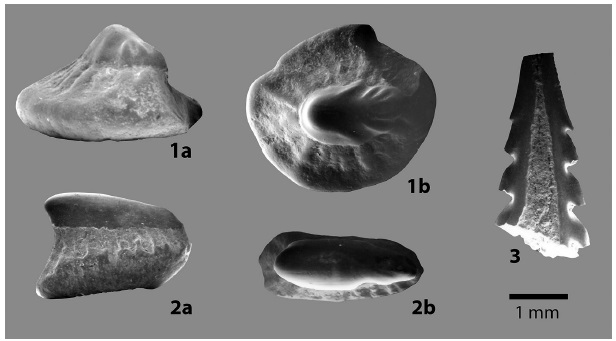
Otros elementos recuperados corresponden a 2 espinas caudales, de las cuales solo se ilustra una. En general presentan un patrón plano con filos aserrados y puntas volcadas hacia el extremo basal. Es posible que la espina corresponda a un Dasyatidae.
Conclusiones
La fauna fósil de Batoidea de la Fm. Uscari en la Cuenca de Limón Sur, es notable por su diversidad, registrando 12 taxones de rayas, repartidas en 5 familias y 8 géneros. La Familia MOBULIDAE Gill 1893 que comprende las especies Mobula hypostoma (Bancroft 1831), Manta cf. M. birostris (Donndorff, 1798), Mobula thurstoni (Lloyd 1908) y Mobula cf. M. japanica (Müller y Henle 1841), especies clave para interpretar el paleoambiente imperante durante la depositación de las secuencias sedimentarias de la Fm. Uscari, en el área de la Localidad de Alto Guayacán durante el límite Mioceno Superior - Plioceno inferior, ya que dichas especies están asociadas a corrientes de surgencia submarina ''upwelling'', ricas en nutrientes y ricas en zooplancton, ello explica la abundante representación de los mobúlidos filtradores de krill y su coexistencia con el tiburón ballena Rhiniodon typus Smith 1828, un tiburón filtrador planctívoro, asociado a dichos mobúlidos (cf. Laurito, 1999).
Por otro lado, la escasísima representación del Myliobatidea Rhinoptera entre la fauna de batoideos, puede explicarse apelando a un registro alotópico sesgado por transporte, en otras palabras, el fragmento de diente pudo ser transportado por corrientes de fondo, desde regiones más someras de la plataforma continental interna o ser un subproducto de depredación de un tiburón.
A diferencia del caso anterior, las rayas Dipturus y Atlantoraja a pesar de su escasa representatividad, se trata de taxones cuyo registro en los sedimentos de Alto Guayacán es acorde a su distribución actual, en sedimentos asociados a plataformas externas e incluso del talud continental, por lo que sus hallazgos deben ser considerados autóctonos y propios de especies epibénticas.
Por su parte, los Rhinobatidae o guitarras, representadas por Rhinobatos sp. en la fauna de Alto Guayacán son un reflejo de las litologías predominantes de la Fm. Uscari en dicha localidad, ya que son habitantes de los fondos lodosos y arenosos. Aunque también, suelen habitar bancos de macroalgas y arrecifes, su dieta está dominada por especies bentónicas como pequeños crustáceos, siendo los peces un componente complementario de su dieta (de la Rosa-Meza et al., 2013). Los Rhynchobatidae o guitarras están representadas en la Fm. Uscari por las especies Rhynchobatus cf. R. pristinus (Probst 1877) y Rhynchobatus rudolffischeriLaurito 1999, que se asumen era bentopelágicas y de habitos, ambientes y dietas similares a los del género Rhinobatos.
Los Dasyatidae en la localidad de Alto Guayacán están representados por 2 especies, Dasyatis dianapizarroaeLaurito 1999 y Taeniurops sergiomorai sp. nov., la gran abundancia de ictiolitos de estas dos especies presentes en los sedimentos en dicha localidad, sugiere una abundancia de alimento, probablemente de crustáceos como los decápodos que constituyen el 45% , seguido por los peces óseos con un 16%, bivalvos con un 2% y cefalópodos con algo menos del 2 % (sensuGilliam, 1991). Este tipo de dieta, para estas especies, se evidencia en la Fm. Uscari, por la abundancia en el registro de otolitos con el que se encontraron asociados, con especies como Brotula, Neobythites, Lepophidium, Paraconger y Ariosoma, entre otros; peces óseos epi-endobentónicos, habitantes comunes de la plataforma externa tropical. Lo anterior permite concluir que tanto D. dianapizarroae como Taeniurops sergiomorai eran organismos desmersales, epibentónicos y pelágicos que solían patrullar el fondo de la plataforma externa para capturar crustáceos y peces óseos.
Por último, la fauna de Batoideos confirma un ambiente de plataforma externa entre los 100 y 200 m de profundidad, zona de coincidencia de todas las especies a excepción de Rhinoptera sp. (fig. 11), zona además, que fue influenciada fuertemente por corrientes de surgencia submarina, aflorantes a lo largo del talud, ricas en nutrientes y zooplancton que dio soporte a un ecosistema con una alta diversidad ictiológica al final del Mioceno y principios del Plioceno. Corrientes de surgencia que se desarrollaron por la elevación del lecho oceánico y la progradación de los sedimentos marinos sobre los arcos insulares en la costa caribeña de América Central Meridional.
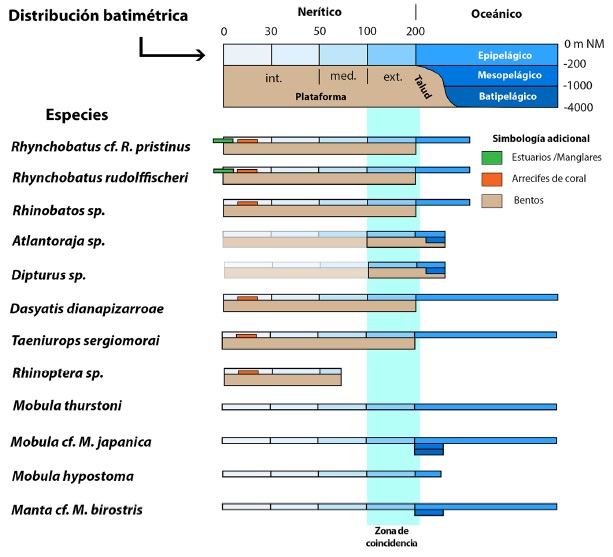
Fig. 11: Gráfico de distribución batimétrica de los Batoidea de la Localidad de Alto Guayacán. La zona de coincidencia corresponde a la plataforma continental externa entre los 100 y 200 m de profundidad. Las áreas difuminadas, corresponden a ambientes infrecuentes o donde raramente han sido observadas las especies indicadas.

