











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
In 1934, R. Lee Collins named a new taxon of sloth, Prepotherium venezuelanum, based on an isolated femur recovered from an outcrop of the Miocene Río Yuca Formation on the Río Tucupido, 11 km west-southwest of the town of Guanare, State of Portuguesa, on the eastern side of the Venezuelan Andes (Fig. 1A; 1B; 1E). This femur was later interpreted as a new genus by Hoffstetter (1961) and named Pseudoprepotherium with a new combination Pseudoprepotherium venezuelanum. Hoffstetter (1961) also recognized that it was from a mylodont and not a prepothere. While Collins was on the faculty at Johns Hopkins University, he did not provide the name of the institutional repository or catalog number for the femur, and the location of the type specimen of P. venezuelanum was lost for many years. Consequently, neither Hoffstetter (1961), when he established the new genus for the species, nor Hirschfeld (1985), when she described Pseudoprepotherium confusum based on the abundant material from La Venta, Colombia, were able to examine the original P. venezuelanum specimen. Therefore, its taxonomic status and phylogenetic relationships to other sloths has not been critically examined based on examination of the type specimen since its original description.
In addition to the type skull (UCMP 39957) of Pseudoprepotherium confusum, Hirschfeld (1985) also described the postcranial skeleton of the species based on both partial skeletons and isolated bones. The detailed descriptions of most of the major bones, including eight femora from six localities of the large sample of P. confusum from La Venta, Colombia made by Hirschfeld (1985), have been the basis for the characters and their state in subsequent phylogenetic studies of sloths that have included Pseudoprepotherium, so our understanding of the genus has been primarily based on this species and not the genotypic specimen. The goal of this paper is to evaluate the two species assigned to this genus and their relationship to each other and other mylodont sloths that have been described from the Socorro and Urumaco formations of Venezuela.
As previously noted, when Collins (1934) described Pseudoprepotherium venezuelanum he did not identify a repository nor provide a catalog number for the specimen. Consequently, it was not known whether the type still existed or if it did where it was located by subsequent researchers. Therefore, the only available information on the anatomy of the type femur was the illustrations included with the original description, so subsequent taxonomic interpretations of the genus and species were restricted to the illustrations and not first-hand examination of the specimen. We discovered that the holotype of Pseudoprepotherium venezuelanum had been transferred to the National Museum of Natural History, Smithsonian Institution in Washington (USNM) (USA) from Johns Hopkins University. It is cataloged as USNM V 13860 (Holotype) with the associated description: Appendicular element, right femur, published by Collins, 1934. Johns Hopkins Univ. Studies Geol. (n.11): 235, pl.15, 16, collected by Love, W. M., in the Miocene of Venezuela.
Recently there has been an increased interest in the vertebrate fauna of the middle and late Miocene Socorro and Urumaco formations of Venezuela (Fig. 1A; 1B; 1D). Sánchez-Villagra and Aguilera (2006) and Sánchez-Villagra, Aguilera, and Carlini eds. (2010) have provided an overview of these formations and their paleontology. This research has included the description of multiple new sloth taxa, and has greatly expanded our understanding of the sloth diversity beyond the only previously described sloth, Urumacotherium garciai (Bocquentin-Villanueva, 1984), based on a partial skeleton including the femur (CIAAP443). Interestingly, like Pseudoprepotherium, this taxon was also originally included in the Prepotheriinae but is now also recognized as a mylodont. Other groups for which new sloth taxa have been described beside the mylodonts (Carlini, Scillato-Yané & Sánchez et al., 2006a; Rincón, McDonald, Solórzano, Núñez & Ruiz-Ramoni, 2015a, 2015b; Rincón, Solorzano, Macsotay, McDonald & Núñez-Flores, 2016a; Rincón, Solorzano, McDonald & Nuñez-Flores, 2016b;) are megatheres (Carlini, Brandoni & Sanchez, 2006b, 2008) and megalonychids (Rincón, Solórzano, McDonald & Montellano-Ballesteros, 2019).
In recent studies of sloths from Urumaco, Falcón State, Venezuela (Rincón et al., 2015a; Rincón et al., 2015b) it was observed that Mirandabradys spp. described by Carlini et al. (2006a), and Pseudoprepotherium venezuelanum (Collins, 1934), from Río Yuca, Portuguesa State, were closely related, while Pseudoprepotherium confusumHirschfeld (1985), was more closely related to Bolivartherium urumaquensis (Carlini et al., 2006a). This analysis resulted in a paraphyletic relationship of these taxa and the recognition of a distinct clade comprised of Mirandabradys spp./ Pseudoprepotherium venezuelanum. Another interpretation is that P. venezuelanum and P. confusum form a monophyletic clade based on cranial and postcranial features (Boscaini, Pujos & Gaudin, 2019). These authors hypothesized this close relationship in their data matrix based on cranial characters (3-5, 14-16, 19, 24, 25, 30, 32, 34, 36-40, 43, 53, 56, 61-76, of the character matrix in Gaudin, 2004), and the humerus (301-303, 306-308, and 313 with new characters proposed by Boscaini et al., 2019). Based on these characters they proposed the taxonomic hypothesis that Pseudoprepotherium confusum and Pseudoprepotherium venezuelanum belong to the same genus, a relationship that has been accepted since Hirschfeld (1985). However, as P. venezuelanum is only known from one femur, and does not have any associated cranial material, these previous results are based on an unfounded assumption that the cranium and femur of P. venezuelanum are morphologically similar to those of Pseudoprepotherium confusum.
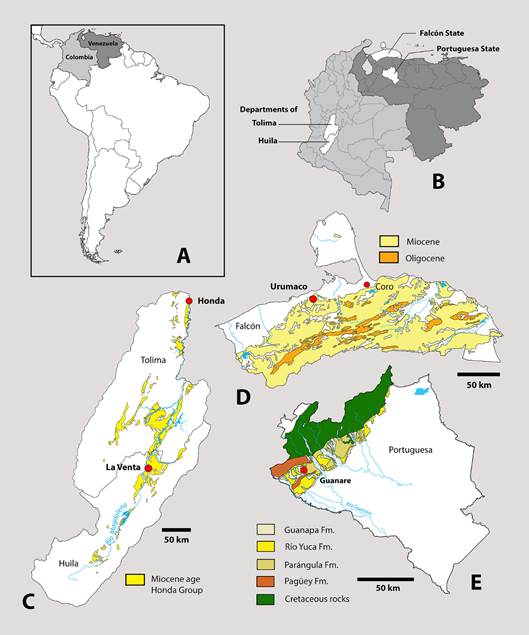
Fig. 1: Location maps, showing the relative position of: A, geographic position of Colombia and Venezuela in to South America; B, geographic position of Huila and Tolima Departments (Colombia), and Falcón and Portuguesa States (Venezuela); C, type locality of Magdalenabradys confusum in La Venta Miocene Age of Honda Group outcrops in Huila and Tolima Departments (Rodriguez Garcia, and Nuñez Tello, 1999; Marquínez and Velandia, 2001); D, Type locality of Magdalenabradys kolossiaia in Urumaco Formation, Falcón State (Rincón, et al., 2019).; E, relative position of locality of Rio Yuca type locality of Pseudoprepotherium venezuelanum, in Portuguesa State (Rincón, et al., 2016a).
Based on our examination of the type of Pseudoprepotherium venezuelanum the purpose of this paper is to refine these relationships by re-examining the morphological features of the femur upon which the various taxa are based and revise the taxonomic relationships of these taxa.
Materials and methods
Abbreviations:
AMU-CURS - Collection of Vertebrate Paleontology of the Alcaldía de Urumaco, Estado Falcón, Venezuela
CIAAP - Centro de Investigaciones Antropológicas, Arqueológicas y Paleontológicas, Coro, Venezuela
MCNC - Museo de Ciencias, Caracas, Venezuela
UCMP - Museum of Paleontology, University of California, Berkeley
USNM - National Museum of Natural History, Smithsonian Institution, Washington D.C.
Dataset
In order to estimate the broader phylogenetic context of the taxa analyzed in this study, we used the dataset of Rincón et al. (2015b). That includes 25 characters based on the femur and tibia. We included 19 members of several lineages of South and North American sloths within the Mylodontidae, Megalonychidae, Megatheriidae and Nothrotheriidae that range in age from the Oligocene to Pleistocene. The character state assignments for the postcranial skeleton of the 19 taxa used in this study are based on direct observation of specimens and information obtained from the primary literature. All of the femora examined have fully fused epiphyses with no visible epiphyseal lines. Consequently, we consider them to all be fully grown adults and at similar ontogenetic stages so that any observed differences are indicative of differences in the genera or species.
Phylogenetic Analysis
We modified the original Character 10 fovea capitis of Rincón et al. (2015b): Present and not open to the border of the femur head (0) (as Urumacocnus); Present and open posteriorly (1) (as Mylodon); Present and open posteromedially (2) (as in Lestodon); or completely absent (3) (as Eionaletherium). Also, we included two more taxa: Lestodon armatus, and Thinobadistes segnis, with data for the latter taken from Webb (1989).
The dataset (Appendix 1) was analyzed using the TNT 1.1 software (Goloboff, Farris & Nixon, 2008). All characters were treated as non-additive (unordered); gaps were treated as missing. The characters were analyzed using the ‘implied weights’ methodology with k = 3. The heuristic parsimony analysis of 1000 replicates was performed using the ‘traditional search option’ (Goloboff et al., 2008). The swapping algorithm used was tree bisection reconnection (TBR), with 10 trees saved per replication, collapsing the trees after each search. To measure node stability, we used the frequency differences (GC) arising from symmetric resampling (p = 33) based on 1000 replicates. The out-group taxon is the North American megalonychid, Megalonyx jeffersonii Desmarest, 1822, as the postcranial skeleton of this taxon is well known and possesses a distinct morphology of the femur and tibia compared to that of mylodonts. In order to elaborate an illustrative final tree, the obtained consensus tree was optimized with the results of the common synapomorphies, the common character modules and the support values.
Systematic Paleontology
Order Xenarthra Cope, 1889
Superfamily Mylodontoidea Gill, 1872
Family Mylodontidae Gill, 1872
PseudoprepotheriumHoffstetter, 1961
SYNONYMY:
MirandabradysCarlini, Scillato-Yané & Sánchez (2006a, p. 256, Fig 1, 3).
TYPE SPECIES. - Pseudoprepotherium venezuelanumCollins (1934).
INCLUDED SPECIES:
Pseudoprepotherium socorrensis nov. comb. (= Mirandabradys socorrensisCarlini et al., 2006a)
Pseudoprepotherium urumaquensis nov. comb. (= Mirandabradys urumaquensisCarlini et al., 2006a)
DIAGNOSIS (Original diagnosis from Hoffstetter, 1961). - Femur 35 to 40% greater than the length of the Santacrucian planopsines from Patagonia (Planops martini and Prepotherium potens); distal end enlarged, relatively narrow and very thick in comparison with the femur length, unlike the wide and depressed form found in Planops martini and Prepotherium potens; intercondylar width is 25% of the femur length in Pseudoprepotherium venezuelanum, but less broad than in Planops martini (35.6%) or Prepotherium potens (34.9%); lateral condyle narrower than the internal which is wider and larger; patellar facet with two prominent anterior crests , the medial l one is prominent as in Prepotherium and Planops and the lateral one less prominent than the medial one, but differs from the Planopsids where it is much less prominent; the medial and lateral condylar facets form a continuous surface with the patellar facet, where the union with the lateral condyle is narrower than the medial , while in Pr. potens the medial union is narrower than the lateral , and in Planops martini they are the same width; the femur head is strong, with scarcely marked neck, more similar to Prepotherium than to Planops; the greater trochanter is thicker, and the third trochanter has a more pronounced protrusion from the diaphysis.
EMENDED DIAGNOSIS. - Medium to large ground sloth with the following unique character combination that distinguishes it from other members of the Mylodontoidea: femur diaphysis curved in shape; valley between the femur head and greater trochanter is shallow; lesser trochanter is poorly developed, and aligned with the diaphysis; proximal end of the femur broader than the distal end; greater trochanter size is larger or closely equal in size to the head; ectepicondyle and entepicondyle are small and only project slightly; connection between patellar and condylar surfaces connected; fovea capitis is present and open posteromedially; femur neck is not well demarcated; femur head angle with respect to the diaphysis is around 80-120º; greater trochanter is almost at same level as the femur head; femur diaphysis transverse shape anteroposteriorly flattened; fossa trochanteric is deep and short; medial condyle is larger than lateral; femur borders on both sides are curved.
REMARKS. - Hirschfeld’s (1985) diagnosis of Pseudoprepotherium is based on the skull of her new taxon, Pseudoprepotherium confusum, and the femur of Pseudoprepotherium venezuelanum. Consequently, her original diagnosis is based on two different genera, Magdalenabradys and Pseudoprepotherium as recognized here. The diagnosis of the genus Mirandabradys by Carlini et al. (2006a) with the type species M. socorrensis was based on the skull, femur, tibia and fibula. The three species, M. socorrensis, M. urumaquensis and M. zabasi, were distinguished by differences in the femur. Characters of the femur used in the diagnosis of the genus include: diaphysis only slightly narrower distally than proximally, widening slightly up to the level of the trochanter minor and anteroposteriorly compressed; trochanter major large and massive; third trochanter major developed on the lateral margin, slightly below the midpoint of the diaphysis and continued distally as a laminar area that merges with the ectepicondyle.
As the femur was utilized by Carlini et al. (2006a) in the diagnosis of both the genus and each species, and is the type specimen of Pseudoprepotherium venezuelanum, we therefore focus on the femur as the basis for the proposed synonymies, due to the lack of cranial material for most of the described taxa. The resulting phylogenetic tree (Fig. 7) clusters Mirandabradys spp. with Pseudoprepotherium venezuelanum based on morphological affinities of the femur of the two taxa, so we consider them to represent the same taxon with Mirandabradys a junior synonym of Pseudoprepotherium.
Features shared by the femora of Magdalenabradys confusum (= Pseudoprepotherium confusum) and Magdalenabradys kolossiaia (= Bolivartherium urumaquensis, in partim) and differ from those for Pseudoprepotherium venezuelanum include the expansion of the lateral margin of the proximal end so that is has a more pronounced convex surface than in P. venezuelanum. The valley between the head and the greater trochanter in P. venezuelanum is shallow, but deeper and more pronounced in the other two taxa. The lesser trochanter is not as developed in P. venezuelanum as in the other two taxa, and the medial margin of the diaphysis is straight between the lesser trochanter and the medial epicondyle, while in M. confusum and M. kolossiaia this area is concave. The muscle scar for the medial vastus muscle is more pronounced in P. venezuelanum, but neither of the muscle scars for the vastus muscle is developed in the other taxa. The width across the distal epicondyles is less than the width of the proximal end in P. venezuelanum, so the femur has a tapered appearance distally, while in M. confusum and M. kolossiaia, the width across the epicondyles and the proximal end are the same. The area distal to the medial epicondyle is laterally expanded in M. confusum and M. kolossiaia while in P. venezuelanum it is narrow, so the patellar surface is centered, whereas in the other two taxa it appears positioned more laterally. The third trochanter in P. venezuelanum has a prominent lateral projection, while in the other two taxa it projects more posteriorly. The lateral epicondyle of P. venezuelanum is less developed and lacks the pronounced lateral projection present in the other two taxa. (Fig. 1A, 1B, 4C’).
Pseudoprepotherium venezuelanum (Collins) 1934
FIGURES 2A; 2B; 5C; 5C’; TABLE 1
EMENDED DIAGNOSIS. - The smallest species of Pseudoprepotherium. Third trochanter is more prominent than any other species of Pseudoprepotherium, and does not project from the diaphysis of the femur relative to the lateral margin of the greater trochanter. Third trochanter position at the middle of the diaphysis. Trochanter minor smaller and less developed than other species of Pseudoprepotherium.
TYPE. - USNM V 13860 (Holotype) right femur, Collins, 1934. Johns Hopkins Univ. Studies Geol. (n.11): 235, pl.15, 16, collected by W.M. Love, Miocene of Venezuela.
TYPE LOCALITY. - Along Río Tucupido, about 7 km NW of the village of Tucupido and about 11 km W-SW of Guanare, approximately 9ºN latitude and 70ºW longitude, Portuguesa state, Venezuela (Fig. 1B; 1E).
AGE AND STRATIGRAPHICAL CONTEXT. - The type specimen of Pseudoprepotherium venezuelanum was collected from an outcrop of the Río Yuca Formation on the Río Tucupido in the southwestern part of Portuguesa State, located about 7 kilometers northwest of the village of Tucupido, and about 11 kilometers west-southwest of the town of Guanare. The locality today is apparently under water resulting from the creation of the Virgen de Coromoto reservoir. Recently, a new vertebrate locality with multiple taxa close to the original locality of Pseudoprepotherium venezuelanum was described (Rincón et al., 2016a). The small fauna consists of at least seven taxa and includes a toxodont cf. Adinotherium, a Peltephilidae armadillo, probably a new genus and species, two freshwaters fishes Platysilurus and Phractocephalus, the caiman Purussaurus, an indeterminate dolphin, and turtles. This vertebrate assemblage is consistent with the proposed Middle to Late Miocene age of the Río Yuca Formation and is considered contemporaneous with the Urumaco Formation.
Pseudoprepotherium socorrensis (Carlini et al., 2006a) nov. comb.
SYNONYMY:
2006a Mirandabradys socorrensis Carlini, Scillato-Yané & Sánchez: 257, Fig. 1.
TYPE. - AMC-CURS 29 partial skull, thoracic vertebra, 3 caudal vertebrae, proximal left ulna, right femur, tibia, and partial fibula, 2 unguals, partial pelvis from one individual.
TYPE LOCALITY. - No specifics provided in original description other than Urumaco, Falcón State, Venezuela (Fig. 1B; 1D).
AGE AND STRATIGRAPHICAL CONTEXT. - Middle Miocene, Socorro Formation.
EMENDED DIAGNOSIS. -Femur has a shorter neck and the trochanter major is positioned lower of the femur head than in Pseudoprepotherium urumaquensis, or P. venezuelesis. Trochanter minor differentiated but not prominent, but more prominent than P. venezuelensis. Patellar facet is shorter proximodistally than in P. urumaquensis.

Fig. 2: Right femur of Pseudoprepotherium venezuelanum (type specimen USNM V 13860). A, anterior view, and B, posterior view.
Pseudoprepotherium urumaquensis (Carlini, Scillato-Yané & Sánchez, 2006a) nov. comb.
SYNONYMY:
2006a Mirandabradys urumaquensis Carlini, Scillato-Yané & Sánchez: 260, Fig. 3, A-F.
TYPE. - AMU-CURS 157 left femur and 10 thoracic vertebrae
TYPE LOCALITY. - No specific locality provided in type description other than Urumaco, Falcón State, Venezuela (Fig. 1B; 1D).
AGE AND STRATIGRAPHICAL CONTEXT. - Late Miocene, Urumaco Formation.
EMENDED DIAGNOSIS. - Larger than P. socorrensis, and P. venezuelensis Proximal half of femur more expanded mediolaterally than in P. socorrensis or P. venezuelensis, and with the distal half of the femur more rotated toward the sagittal plane. Trochanteric fossa well developed. Trochanter minor more developed than in P. socorrensis, or P. venezuelensis.
Third trochanter more prominent than P. socorrensis or P. venezuelensis. Entepicondyle prominent and projecting medially as in P. socorrensis. Patellar facet more developed and straighter than in the other two species (see Carlini et al., 2006a). Patellar surface longer than wide, than any other Pseudoprepotherium species. The lesser trochanter is well developed and positioned slightly caudally but more medially directed.
Pseudoprepotherium zabasi (Carlini, Scillato-Yané & Sánchez, 2006a) nomen dubium.
SYNONYMY:
2006a Mirandabradys zabasi Carlini, Scillato-Yané & Sánchez: 260, Fig. 3, G-N.
Table 1: Femur length of some Mylodontidae: Eionaletherium, Pseudoprepotherium, rumacotherium and Magdalenabradys.
| Taxa | Femur Length |
| Pseudoprepotherium venezuelanum | 428 mm |
| Eionaletherium tanycnemius | 510 mm |
| Urumacotherium garciai | 560 mm |
| Pseudoprepotherium socorrensis nov. comb. | 560 mm |
| Pseudoprepotherium urumaquensis nov. comb. | 594 mm |
| Magdalenabradys kolossiaia sp. nov. | 630 mm |
| Magdalenabradys confusum new comb. | 451-504 mm |
TYPE. - AMU-CURS 27 left femur almost complete lacking the area between the trochanter major and the head and the patellar facet.
TYPE LOCALITY. - No specific locality provided in type description other than Urumaco, Falcón State, Venezuela (Fig. 1B; 1D).
AGE AND STRATIGRAPHICAL CONTEXT. - Early Pliocene, Codore Formation.
REMARKS. - The holotype of P. zabasi (AMU-CURS 127), was based on an incomplete left femur, lacking the area between the trochanter major and the head, and the patellar facet. In the accompanying illustration (Carlini et al., 2006a Fig, 3G-N) the trochanter major is represented by only its lateral posterior part, and it is very difficult to say how massive it would have been, based on what is preserved. Additional specimens referred to this taxon by Carlini et al. (2006a) includes AMU-CURS 128, a distal half of a left femur, which lacks the critical diagnostic features of the major and minor trochanters, so it is not possible to determine whether it had the diagnostic morphology that would permit its placement in this species. It is also not possible to determine whether this specimen (AMU-CURS 128) has a trochanter minor that is less developed than in P. socorrensis or P. urumaquensis, and if the trochanter major is more massive than in other species of Pseudoprepotherium. Additional material referred by Carlini et al. (2006a) to this taxon includes (right femur, MCN-USB 16-04 PB) to P. zabasi, which was first referred by Linares (2004: Fig. 7) to Lestodon urumaquensis. These specimens do not have a massive trochanter major considered to be a diagnostic feature of P. zabasi by Carlini et al. (2006a), and also the trochanter minor is lacking in the original figure by Linares (2004), so lacks the critical diagnostic features that would allow it to be assigned to P. zabasi. We therefore consider this taxon to be a nomen dubium.
Magdalenabradys gen. nov.
SYNONYMY:
1985 Pseudoprepotherium confusum Hirschfeld: 5, Fig. 1 2006a Bolivartherium urumaquensis Carlini, Scillato-Yané & Sánchez: 260, Fig.4, G-L (In partim, MCNC-85-72V, not Fig. 4 A-F).
TYPE SPECIES. -Magdalenabradys confusum (Hirschfeld, 1985) nov. comb.
INCLUDED SPECIES:
Magdalenabradys kolossiaia nov. sp.
TYPE SPECIMEN OF TYPE SPECIES. - UCMP 39957 crushed skull
TYPE LOCALITY. - About 9 km E of Villavieja and 2.5 km NE of Hacienda Argentina, Huila, Colombia (Fig. 1A; 1B; 1C).
AGE AND STRATIGRAPHICAL CONTEXT. - Ferruginous (Yellow) Sandstone locality, UCMP V-5045, Villavieja Formation, Honda Group, Miocene, Laventan SALMA.
DIAGNOSIS. - Large ground sloth with the following unique character combination that distinguishes it from other members of the Mylodontoidea: a deep valley between the femur head and the greater trochanter; third trochanter projects posteriorly; third trochanter at the middle of the diaphysis; ectepicondyle and entepicondyle: robust and prominently projected laterally and medially, respectively; greater trochanter size larger or closely equal in size to the head; greater trochanter extends above the femur head.
Magdalenabradys confusum (Hirschfeld) (1985)
FIGURES 3; 5B; 5B’; TABLE 1
SYNONYMY:
1985 Pseudoprepotherium confusum Hirschfeld: 5, Fig. 1
EMENDED DIAGNOSIS. - Mylodontid with the shorter femur (451 - 504 mm) about 25% smaller than Magdalenabradys kolossiaia, with the lesser trochanter smaller than M. kolossiaia; Skull long and narrow, nasal region not greatly expanded dorsally as in later mylodonts; dental formaula 5/4, molariform teeth simpler than those post-Miocene mylodonts, resembling Orophodon haploides Ameghino, 1895, of Deseadan, but wider medio-laterally with greater proportion of vasodentine; no diastema separating M1/M1 from remaining molariform series; preorbital region long, maxillary extends anterior to the M1 rather than terminating at level of M1; radius and ulna long and slender, ulna equal in length to humerus; Mc II large in proportion to Mc III, facets for Mc I and Mc III planar; Mc V with well developed carina; femur long and slender with well-developed third trochanter; greater trochanter extends above the femur head; femur diaphysis transverse shape anteroposteriorly flattened; patellar surface shorter than wide; longitudinal ridges of the vastus lateralis and medialis on anterior surface; external and internal condyles broadly connected to relatively narrow trochlea; intercondylar notch narrow; well-marked trochanteric fossa; tibia proportionally longer and not as “twisted”as in Glossotherium; fibula long ans slender; inner astragalar condyle not as developed to from a tibial knob, and outer condyle not extended posteriorly as in Glossotherium; two facets between calcaneum and astragalus; calcaneum lacks posterior extension (emendation in bold).
TYPE SPECIMEN. - Hirschfeld (1985) assigned as the type of M. confusum a crushed and distorted skull. Her description of the femur is based on eight specimens from six localities, including one femur from a partial skeleton that included a skull and mandible (UCMP 3800, 37999). Hirschfeld (1985) noted that the femur of M. confusum was larger than the type of P. venezuelanum but was similar in shape and proportions. Differences between the species were the more pointed proximal projection of the greater trochanter, shorter and more prominent third trochanter, and the medial condyle with a more prominent proximal projection in M. confusum (Hirschfeld, 1985, p. 25). The location of the type specimen of Pseudoprepotherium venezuelanum was unknown at the time of Hirschfeld’s research, so was not available for examination. Consequently, her comparison of the two taxa was limited to the illustrations in Collins (1934: Pl XV, XVI) which illustrated the femur in anterior, medial, posterior, proximal and distal views. It should be noted that the illustration in Collins (1934) shows part of the shaft is missing distal to the third trochanter. Also, the preserved portions of the distal end cannot be directly connected to the proximal two-thirds of the femur, making it impossible to obtain an accurate measurement of the length of the bone, although Collins (1934:240) did provide a length measurement of 419 mm compared to M. confusum which ranges in length from 460 to 485 mm (N=4) (Hirschfeld, 1985, Table 15).
Magdalenabradys kolossiaia nov. sp.
FIGURES 4; 5A; 5A’; TABLE 1
SYNONYMY:
2006a Bolivartherium urumaquensis Carlini, Scillato-Yané & Sánchez: 260, Fig.4, G-L (In partim, MCNC-85-72V, not Fig. 4 A-F).
TYPE SPECIMEN. - Left and right femur corresponding to a single individual (MCNC-85-72V).
TYPE LOCALITY. - 1.75 km NW of kilometer 159.4 on the oleoducto (oil pipeline). Linares (2004) published a map with coordinates for this locality, which is identified as locality number 78. This is correlative with the Bryan Patterson locality number 85-72V. The coordinates of the type locality for Magdalenabradys kolossiaia are 11°12’29.90”N, 70°18’57.00”W (Fig. 1A; 1B; 1D).
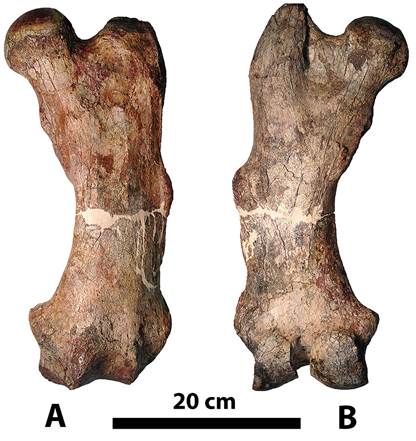
Fig. 3: Left femur of Magdalenabradys confusum (UCMP-41137). A, anterior view, and B, posterior view.
AGE AND STRATIGRAPHICAL CONTEXT. - 800 m below the Capa de Huesos, Middle Urumaco Formation, Huayquerian SALMA
ETYMOLOGY. - Kolossiaia (κολοσσιαία) from Greek form meaning colossal
DIAGNOSIS. - Mylodontid with femur length (650 mm), 25 % larger than M. confusum, with the lesser trochanter more prominent than in M. confusum; greater trochanter almost at same level as the femur head; femur diaphysis transverse shape cylindrical to oval; patellar surface length and width are similar, shape is roughly square.
REMARKS. - Linares (2004) described Lestodon urumaquensis based on a fragment of a right maxilla with the alveoli of the caniniform to fourth molariform, the posterior part of the cranium and a partial mandible with both rami joined at the symphysis (MCN 170-72v). The type location for Magdalenabradys kolossiaia (= Bolivartherium urumaquensis) is 11°15’35.35”N, 70°14’2.33”W Linares locality 26-FU (FU=fossil Urumaco).
The type specimen (MCNC-170-72V) lacks any parts of the postcranial skeleton. Carlini et al. (2006a) reexamined the specimen and determined it was different from Lestodon and established a new genus Bolivartherium urumaquensis. The original description of Bolivartherium urumaquensis also included additional material referred to the species. The second specimen, MCNC-85-72V, includes both femora, pelvis and 12 associated vertebrae, and an isolated right humerus (MCN 71-72). The two femora and vertebrae (MCNC-85-72V) were collected at kilometer 159.4 of the Urumaco oil pipeline, and 800 meters below (south) of “Capa de Huesos” which includes the contact between the Codore and Urumaco Formations. As the type and referred specimens lack any skeletal elements in common reference of the post-cranial skeleton to the associated skull and mandible is dubious. It should also be noted that the localities where the two specimens were collected are separated by 8 kilometers. For that reason we believe that the femora assigned to Bolivartherium (MCNC-85-72V) do not belong to that taxon, but the cranial and mandible remains (MCNC-170-72V) belong to Bolivartherium urumaquensis and remains as valid taxon.

Fig. 4: Femur of type specimen (85-72V) of Magdalenabradys kolossiaia. Right femur in A, anterior view, and A´ posterior view. Left femur in B anterior view, and B´posterior view
Morphologically, the type cranium of Bolivartherium urumaquensis (MCN 170-72v) with a distinct caniniform separated from the molariforms by a diastema and positioned lateral to the axis of the molariform tooth row permits its reference to the Lestodontinae.
Carlini et al. (2006a) described the femur as having a wide and massive proximal third with a conspicuous third trochanter. However, the referred femora do not resemble the femur of Lestodon. In Lestodon the head lacks a distinct neck and does not project proximally much above the low greater trochanter. The valley between the head and greater trochanter is very shallow. The proximal end of the femur does not have the relatively greater expansion of the proximal end relative to its length as in the femur referred to Magdalenabradys kolossiaia (=Bolivatherium urumaquensis, in partim). The fovea of the head consists of a prominent emargination located on the posteromedial margin, as in M. kolossiaia. The trochanteric fossa is large in Lestodon and its distal margin is at the level of the lesser trochanter while in M. kolossiaia its distal margin is above the lesser trochanter. The width of the shaft retains approximately the same width its entire length but while the lateral margin is relatively straight, the medial margin is concave from the lesser trochanter to the medial epicondyle. The third trochanter is distally elongated and positioned more distally on the shaft so most of it is distal to the midpoint of the shaft. The third trochanter lacks the posterior reflection present in the femur referred to Bolivartherium urumaquensis (now Magdalenabradys kolossiaia) by Carlini et al. (2006a). The area of insertion for the medial vastus muscle is formed by two short muscle scars, and the muscle scar for the lateral vastus is also short and all are confined to the proximal half of the shaft. The distal epicondyles are well developed but are not in the same plane as the lateral epicondyle, which is positioned more proximally on the shaft relative to the medial condyle, while in M. kolossiaia they are in the same plane. The patellar articular surface is broad, deeply concave mediolaterally and covers about three-quarters the width of the distal end of the femur. It forms a continuous articular surface with both of the distal condyles. In M. kolossiaia it is mediolaterally narrower, and not as concave and deep, although it also has a continuous articular surface with the distal condyles.

Fig. 5: Proximal view of: A, Magdalenabradys kolossiaia (85-72V); B, Magdalenabradys confusum (UCMP-41137); C, Pseudoprepotherium venezuelanum (type specimen USNM V 13860); and distal view of A’, Magdalenabradys kolossiaia (85-72V); B’, Magdalenabradys confusum (UCMP-41137); C’, Pseudoprepotherium venezuelanum (type specimen USNM V 13860).
Magdalenabradys has a deep valley between the femur head and the greater trochanter as an apomorphy, however that character is also seen in Pyramiodontherium (a megathere) and Analcimorphus.
In M. kolossiaia the lesser trochanter is well-developed, and slightly caudally but more medially directed. This character is shared by Eionaltherium, Glossotherium, and Magdalenabradys. However this character is also present in Analcimorphus and H. longiceps-H.ruetimeyeri, which differs from M. confusum as it has a poorly developed lesser trochanter, and is aligned with the diaphysis, as in G. wegneri, but in Eionaletherium the lesser trochanter is well-developed, and caudally and medially directed.
M. confusum and M. kolossiaia also share a third trochanter that projects posteriorly, as in Eionaltherium, Glossotherium ,Magdalenabradys, P. urumaquensis and P. socorrensis.
M. confusum and M. kolossiaia share a third trochanter positioned at the middle of the diaphysis, which is also present in Eionaltherium, Glossotherium, and Magdalenabradys, but again this character is present in Analcimorphus and P. venezuelanum. In Glossotherium wegneri it is distal to the middle of the diaphysis, and in Eionaletherium it is proximal to the middle of the diaphysis.
M. confusum and M. kolossiaia share a greater trochanter larger or closely equal in size to the head, as in Glossotherium wegneri and Pseudoprepotherium spp.
Ectepicondyle and entepicondyle are robust and prominently project laterally and medially, respectively in Eionaltherium, Glossotherium, and Magdalenabradys, which share the character only with the megathere, Eremotherium laurillardi.
In M. confusum and M. kolossiaia the fovea capitis is present and open on the posteromedial margin of the head, as in Lestodon, Thinobadistes, Pseudoprepotherium and Paramylodon.
The greater trochanter extends proximally above the femur head, and is an apomorphy in M. confusum, and in M. kolossiaia the greater trochanter is at the same level as the femur head, as in Glossotherium wegneri, H. longiceps-H.ruetimeyeri and Pseudoprepotherium spp.
In M. confusum the transverse cross section of the diaphysis of the femur is anteroposteriorly flattened, as in Eionaletherium, Glossotherium wegneri, Paramylodon, Chubutherium and Pseudoprepotherium, but in M. kolossiaia the femur diaphysis is cylindrical to oval in transverse shape, so is characteristic of the Eionaltherium, Glossotherium, and Magdalenabradys, but is also shared with Urumacotherium (Fig. 6; Table 1), Analcimorphus and H. longiceps-H.ruetimeyeri.
In M. confusum the patellar surface is shorter than wide, as in G. wegneri, Chubutherium, Analcimorphus, H. longiceps-H.ruetimeyeri and Pseudoprepotherium spp, except P. urumaquensis. In M. kolossiaia the length and width of patellar surface is similar as in Urumacotherium garciai.
Morphologically the femur of P. venezuelanum is more similar to Mirandabradys (=Pseudoprepotherium) than to M. confusum. Characters (from Rincón et al., 2015b) that these two taxa (Pseudoprepotherium and Mirandabradys) share in common include 1(1), 2(1), 3(2), 6(0), 7(1), 11(1), 13(2), 14(1), 16(0) and a femur diaphysis that is curved [character 1(1) ]. However, that character is also present in many other Mylodontidae including Paramylodon, Eionaletherium, G. wegneri, Magdalenabradys spp., and Chubutherium (Fig. 7).
Valley between the femur head and the greater trochanter is deep [character 2(1) ], and is shared also with Paramylodon, Eionaletherium, G. wegneri, Urumacotherium, and H. longiceps-H.ruetimeyeri
Lesser trochanter is poorly developed, and aligned with the diaphysis [character 3(2) shared with many Megatheriidae, Eremotherium laurillardi, Pyramiodontherium brevirostrum, Pyramiodontherium scillatoyanei, Analcimorphus giganteus), a nothrothere, Nothrotheriops shastensis and some Mylodontidae, Paramylodon, G. wegneri, M. confusum, Chubutherium, and Urumacotherium.
Proximal end of the femur is broader than the distal end [character 6(0) ] shared with Chubutherium, Magdalenabradys spp., G. wegneri, and Paramylodon.
Greater trochanter size is larger or closely equal in size to the head [character 7(1) ] shared with G. wegneri and Magdalenabradys.
Femur neck is not well-demarcated [character 11(1) ] shared with Chubutherium and Urumacotherium.
Greater trochanter is almost at same level as the femur head 13(2) and shared with G. wegneri, M. kolossiaia and H. longiceps - H. ruetimeyeri
Femur diaphysis transverse shape is anteroposteriorly flattened 14(1) shared with Chubutherium, M. confusum, G. wegneri, Eionaletherium and Paramylodon.

Fig. 6: Right femur of Urumacotherium garciai (UNEFM-CIAAP-433). A, anterior view, and B, posterior view.
Trochanteric fossa is deep and short 16(0) shared with Chubutherium, Eionaletherium and Paramylodon.
Discussion
Prepotherium was first described by Ameghino (1891) from the early Miocene Santacrucian SALMA. There are three recognized species of the genus, the genotypic species, Prepotherium potens, together with P. filholi and P. moyani. Specimens referred to P. potens include YPM-VPPU 15345, an incomplete post-cranial skeleton including a right femur, which permits comparison with the type of P. venezuelanum. The genus Prepotherium is known from the Miocene Santa Cruz, and younger Collon-Cura Formation of Argentina. Prepotherium and the closely related Planops are regarded as members of the Megatheriidae by De Iuliis (1994) while taxa referred to Pseudoprepotherium and the other genera discussed here are considered members of the Mylodontidae.
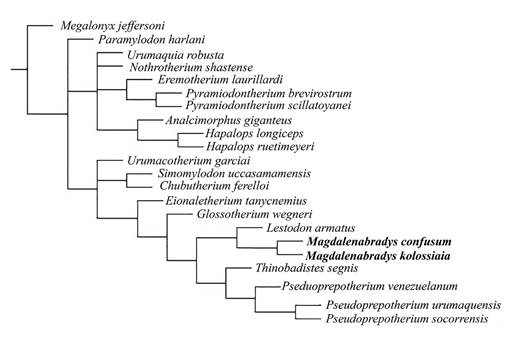
Fig. 7: Majority rule tree using the prior weights (k = 3) methodology tree bisection reconnection (TBR) score = 88, consistency index (CI) = 0.352 and retention index (RI) = 0.637. Character/taxon matrix modified after Rincón et al. (2015a, b).
The femoral head of Prepotherium is hemispheric, as in extant xenarthrans, relatively large, inclined anteriorly, and the fovea capitis is very well developed suggesting a robust teres femoris ligament. The greater trochanter is robust in Prepotherium and large, as in Myrmecophaga, but is laterally directed and does not reach as far proximally as the femoral head, as in extant sloths. The femoral diaphysis is wide transversely, especially in Prepotherium, the largest Santacrucian sloth, due to the large greater trochanter, expanded third trochanter and the wide distal epiphysis. The large and prominent third trochanter is positioned approximately at midshaft or very slightly distal to midshaft and protrudes more in Prepotherium than in extant sloths and anteaters, so more closely resembles the extant armadillos, Priodontes, Cabassous and Chaetophractus. In Prepotherium the medial lip of the patellar groove is more pronounced than the lateral one (Toledo, Bargo & Vizcaíno, 2015).
The type material of Bolivartherium urumaquensis is represented by cranial fragments, and remains as a valid taxon until more detailed studies. Carlini et al. (2006a, p. 264) stated “The Lestodontinae (Mylodontidae) is represented in the Urumaco Formation by the two species of Bolivartherium. These are characterized by their low, long and straight skull, large caniniform, divergent molariform series and a femur that has a wide and massive proximal third with a conspicuous third trochanter”. The two associated femora (MCN 85-72V) were referred to the genus by Carlini et al. (2006a), but they did not provide any morphological comparisons of the femur of Bolivartherium with any other Lestodontinae or Mylodontidae except Mirandabradys. As discussed previously, there are significant morphological differences between the femora referred to Bolivartherium and Lestodon, suggesting the femora are not from that taxon, while the femora of Bolivartherium and Magdalenabradys confusum have multiple features in common. Likewise, as they are morphologically distinct from the type of Pseudoprepotherium, so both femora previously assigned to Bolivartherium are assigned here to a new genus, Magdalenabradys.
Based on the cranium there are significant differences between the types of Bolivartherium urumaquensis (MCN 170-72v) and Magdalenabradys confusum (UCMP 39957). The primary difference is the modification of the first tooth into a caniniform, which is separated from the molariforms by a large diastema in Bolivartherium, while in Magdalenabradys the first upper tooth is not modified into a caniniform and there is not a diastema between it and the following molariform. The tooth rows in Magdalenabradys are parallel while in Bolivartherium they diverge anteriorly and the first tooth (caniniform) is lateral to the axis of the molariform tooth series.
Carlini et al. (2006a) state the femur (MCN 85-72V) belongs to a Lestodontinae (Mylodontidae) because it has a wide and massive proximal third with a conspicuous third trochanter” a description that also applies to Magdalenabradys confusum.
The femur of Pseudoprepotherium venezuelanum is more columnar than M. kolossiaia or M. confusum. Additionally, the medial side in P. venezuelanum is less curved than M. kolossiaia or M. confusum. The third trochanter in P. venezuelanum is completely restricted to the lateral margin of the shaft, but in M. kolossiaia or M. confusum it is shifted posteriorly on the shaft. The trochanter minor is greatly reduced and close to absent in P. venezuelanum, and while it is reduced in M. kolossiaia or M. confusum it is still a distinctive process. The medial epicondyle in M. kolossiaia and M. confusum is more projected than in P. venezuelanum.
There are numerous features that distinguish Pseudoprepotherium and Magdalenabradys from Urumacotherium garciai, another mylodont from the Urumaco Formation. In Urumacotherium garciai the head extends proximally well above the greater trochanter, which is not well developed, and there is essentially no valley between the greater trochanter and head. There is no trochanteric fossa. The femur maintains an essentially consistent width from proximal to distal end with straight and parallel lateral and medial margins. Urumacotherium also lacks a third trochanter. The epicondyles are not well developed so the width across the epicondyles is only slightly greater than that of the condylar width.
Pseudoprepotherium venezuelanum was described from the middle to late Miocene of Portuguesa state, in Venezuela (Collins, 1934; Rincón et al., 2016a). This places it within the chronological range of Mirandabradys with M. socorrensis from the Middle Miocene (Socorro Formation), and M. urumaquensis from the Late Miocene (Urumaco Formation).
Summary and Conclusions
The recent phylogenetic hypothesis of the relationships of the northern South America Mylodontidae from the Miocene proposed by Rincón et al. (2015a) resulted in a paraphyletic relationship of P. confusum and P. venezuelanum. In that analysis P. confusum, nested with B. urumaquensis based on the referred material (left and right femur, pelvis and some vertebrae associated, of a single individual), assigned to that taxon by Carlini et al. (2006a) and not with P. venezuelanum. While the femur is from a mylodont, since the remains do not have any common element comparable with the type of Bolivartherium urumaquensis, an associated skull and mandible, there is no a priori reason to refer this specimen to Bolivartherium. Boscaini et al. (2019, Figure 4) recovered a close relationship between Pseudoprepotherium venezuelanum, P. confusum and Mirandabradys spp., with Urumacotherium as the sister taxon, but that analysis is based on the assumption that the cranium of P. venezuelanum is similar to that of P. confusum of Hirschfeld (1985) which cannot yet be confirmed because cranial remains for P. venezuelanum are not yet known, resulting in an artificial relationship with the other taxa for which cranial remains are known. Our analysis presented here confirms this relationship and additionally that Pseudoprepotherium confusum, is distinct from P. venezuelanum and cannot be referred to that genus. Based on the analysis presented here the type specimens of Pseudoprepotherium venezuelanum nests with Mirandabradys from Urumaco. Therefore, we consider Mirandabradys of Carlini et al. (2006a) to be a junior subjective synonym of PseudoprepotheriumHoffstetter (1961). We therefore propose a new genus for the species as a new combination. Magdalenabradys confusum along with the post-cranial material from Venezuela previously included in B. urumaquensis to the new genus Magdalenabradys.
We propose also that two of the three species of Mirandabradys be included in Pseudoprepotherium as: Pseudoprepotherium socorrensis nov. comb. (Carlini et al., 2006a); and Pseudoprepotherium urumaquensis nov. comb. (Carlini et al., 2006a) but consider Pseudoprepotherium zabasiCarlini et al., 2006a) a nomen dubium.
Finally, we propose a new species of Magdalenabradys, M. kolossiaia based on the left and right femur with associated pelvis and vertebrae, corresponding to a single individual (MCNC-85-72V) originally referred to Bolivartherium urumaquensis by Carlini et al. (2006a) based on the femur’s morphological similarity to M. confusum from La Venta, Colombia.
Currently we recognize the following diversity of mylodonts from the Socorro and Urumaco Formations, (Table 2) although not are all contemporaries and some species occur at different levels within the formation: Urumacotherium garciai, Pseudoprepotherium socorrensis, Pseudoprepotherium urumaquensis, Bolivartherium urumaquensis, Bolivatherium codorensis, Eionaletherium tanycnemius, and Magdalenabradys kolossiaia (Table 2).
Table 2: Sloth taxa currently recognized from the Urumaco Region. (Bocquentin-Villanueva, 1984; Carlini, et al., 2006a; Carlini, et al., 2006b; Rincón, et al., 2015a; Rincón, et al., 2016a; Rincón, et al., 2019).
| - | Socorro Formation | Urumaco Formation | Codore Formation |
| Family Megalonychidae | - | - | - |
| Urumacocnus urbanii | - | X | - |
| Pattersonocnus diazgameroi | - | X | - |
| Family Megatheridae | - | - | - |
| Urumaquia robusta | - | X | X? |
| Family Mylodontidae | - | - | - |
| Eionaletherium tanycnemius | - | X | - |
| Urumacotherium garciai | X | ||
| Pseudoprepotherium socorrensis | X | - | - |
| Pseudoprepotherium urumaquensis | - | X | - |
| Magdalenabradys kolossiaia gen. et sp. nov. | - | X | - |
| Bolivartherium urumaquensis | - | X | - |
| Bolivartherium codorensis | - | - | X |
| Number of Taxa by formation | 1 | 8 | 2 |

