











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Los gonfoterios (Mammalia, Proboscidea, Gomphotheriidae) migraron de Asía a América del Norte durante el Mioceno medio (~15 Ma; Tedford et al., 2004) y posteriormente incursionaron en América Central durante el Mioceno tardío (Lucas y Alvarado, 2010), con la consecuente diversificación y dispersión hacia América del Sur al final del Plioceno tardío y Pleistoceno temprano durante el Gran Intercambio Biótico Americano (Webb, 1991; Mothé y Avilla, 2013; Mothé y Avilla, 2015). Aunque en algún momento la presencia de Amahuacatherium peruvium en el Mioceno tardío de la Amazonia peruana, sugería que los gonfoterios arribaron a ese subcontinente antes de lo que se ha planteado (Campbell et al., 2000; 2009). Mothé y Avilla (2015) revisaron una muestra importante de ejemplares bien conservados, en la cual consideraron inválido a A. peruvium, debido a que las características morfológicas están dentro del rango de variabilidad de Notiomastodon platensis,argumentando que los registros más antiguos de gonfoterios en América del Sur sugieren que el arribo de esta familia probablemente ocurrió durante el Plio-Pleistoceno, después del cierre del istmo de Panamá.
Tradicionalmente, los gonfoterios se dividen en brevirostrinos y longirostrinos. Los gonfoterios brevirostrinos se caracterizan por tener una sínfisis corta; incisivos superiores rectos o recurvados, con o sin esmalte, e incisivos inferiores ausentes o muy reducidos (Frick, 1933). Por su parte, los gonfoterios longirrostrinos, considerados un linaje más primitivo, se caracterizan por tener una mandíbula larga que se extiende anteriormente debido a la presencia de incisivos inferiores; premolares (deciduos y definitivos) y primeros molares de tipo bunodontes-bilofodontes; molares permanentes intermedios trilofodontes y terceros molares tetralofodontes (Frick, 1933; Ferrusquía-Villafranca, 1990a; Lucas y Alvarado, 1995; Aguilar y Laurito, 2011; Valerio y Laurito, 2008). El patrón de la morfología oclusal en toda la serie dental es simple (como en la mayoría de las formas primitivas) y los molares son de menor talla que en las formas brevirostrinas del Pleistoceno, hasta un 50% (Valerio y Laurito, 2008). En este grupo se incluyen Rhynchotherium y Gomphotherium (Shoshani y Tassy, 2005; Ferreti 2008a). Rhynchotherium tiene un amplio registro en América del Norte y es considerado endémico de la región (Lambert, 1996; Lucas y Morgan, 2008). Por su parte, Gomphotherium tuvo una distribución más amplia, desde América del Norte hasta América Central (Webb y Perrigo, 1984; Lucas y Alvarado, 1995; Laurito y Valerio, 2007; Valerio y Laurito, 2008; Aguilar y Laurito, 2011; MacFadden et al., 2015) y no se tiene evidencia de su existencia en América del Sur.
Gomphotherium es considerado el primer gonfoterio bunodonte en arribar a América y algunos autores creen que las formas posteriores se originaron a partir de este género (Tobien, 1976; Lambert y Shoshani, 1998; Arroyo-Cabrales et al., 2007).
En las faunas de México, principalmente las pleistocénicas, se encuentran representados los gonfoterios brevirrostrinos Stegomastodon y Cuvieronius, cuya distribución en el país está bien documentada (Alberdi y Corona, 2005; Arroyo-Cabrales et al., 2007). Por el contrario, los registros de gonfoterios longirrostros son escasos y provienen de las localidades que datan de los NALMA Barstoviano (Mioceno medio) al Blancano (Plioceno), que se encuentran distribuidas desde el norte al sur de México (ver Cuadro 1).
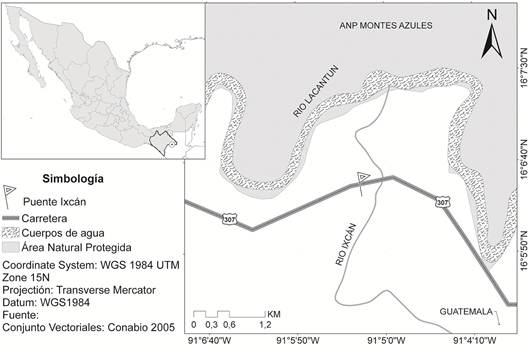
En este trabajo describimos un P4 permanente, proveniente de la localidad Puente Ixcán, municipio de Maravilla Tenejapa, Chiapas, sureste de México (Fig. 1), el cual es referido al género Gomphotherium. El único registro previo de Gomphotherium para Chiapas se conoce para la Fauna Ixtapa-Soyaló, de edad Mioceno medio (NALMA Barstoviano temprano) y está basado en un segundo molar superior derecho y un fragmento de un premolar deciduo (FerrusquíaVillafranca, 1990a). Por tal motivo, el ejemplar reportado en este trabajo es significativo ya que documenta por primera vez la presencia de Gomphotherium en el Mioceno tardío de Chiapas, y por su cercanía con Centroamérica contribuye al conocimiento de la distribución de los gonfoterios longirrostrinos en esta región.
Marco geológico y paleontológico
En la localidad Puente Ixcán (Fig. 1) el levantamiento de la sección es preliminar. Allí aflora una secuencia de sedimentos detríticos no claramente diferenciados de 7-8 m de espesor, dispuestos en posición horizontal, que es cruzada por una falla vertical. La litología se compone principalmente por arena de grano medio a grueso bien compactado, de color gris oscuro a gris claro con gravas subangulares, en la que se entremezclan lentes de arena de grano grueso que llegan a medir hasta 1,5 metros de amplitud y 1 metro de espesor, que contienen abundantes restos de moluscos marinos articulados (gasterópodos y bivalvos), aún sin determinar taxonómicamente. Asociado a los restos de Gomphotherium se encuentran restos de tortugas trioníquidas y dermatémidas, un Caimaninae identerminado, cf. Crocodylus sp., un Equidae indeterminado y el rinoceronte Teleoceras cf. T. hicksi (Carbot-Chanona, 2011; Brochu y CarbotChanona, 2015; Carbot-Chanona, en prensa). Los restos óseos presentan marcas de arrastre lo que indica que estos fueron transportados y depositados posteriormente a los gasteropodos. La presencia de Teleoceras cf. T. hicksi indica una edad del Mioceno tardío (NALMA Henfiliano) para la asociación faunística (Prothero, 2005).
Método
El ejemplar se extrajo de forma manual usando un jacket para su protección. En laboratorio se retiró el sedimento adherido usando puntas odontológicas y un cepillo de cerdas suaves y agua corriente. La pieza se consolidó usando pegamento a base de cianocrilato disuelto en acetona al 100%. El molar se comparó morfológica y merísticamente con molariformes bilofodontes deciduos y permanentes de ejemplares de Gomphotherium, Rhynchotherium, Stegomastodon y Cuviernonius reportados en la bibliografía (Cuadro 2), así como con dos ejemplares de Cuviernonius hyodon juveniles (IHNFG-2691, mandíbula, e IHNFG-4864, cráneo), alojados en la colección paleontológica de la Secretaría de Medio Ambiente e Historia Natural de Chiapas, México.
Cuadro 1 Localidades en México con presencia de gonfoterios longirrostros.
| Localidad | Estado | Edad | NALMA | Referencia |
|---|---|---|---|---|
| Rhynchotherium | ||||
| Santa Ana | Baja California Sur | Plioceno temprano | Blancano | Miller (1980) |
| Incierta | Chihuahua | Plioceno | Blancano | Carranza-Castañeda y Miller (2004) |
| El Ocote | Guanajuato | Mioceno tardío | Henfiliano | Miller y Carranza-Castañeda (1984) |
| San Juan Unión, Taxco | Guerrero | Plioceno temprano | Blancano | Alberdi y Corona-M. (2005); Corona-M y Alberdi (2006) |
| Tula | Hidalgo | Plioceno temprano | Blancano | Carranza-Castañeda (2006) |
| La Suerte, Lagunillas | Hidalgo | Plioceno temprano | Blancano | Carranza-Castañeda (2006) |
| Santa María Amajac | Hidalgo | Plioceno temprano | Blancano | Carranza-Castañeda (2006) |
| Ajijic | Jalisco | Incierta | Incierto | Alberdi y Corona-M. (2005) |
| Ameca | Jalisco | Incierta | Incierto | Alberdi y Corona-M. (2005) |
| Zona Chapala-Zacoalco | Jalisco | Incierta | Incierto | Alberdi y Corona-M. (2005) |
| Tecolotlán | Jalisco | Mioceno tardío | Henfiliano | Carranza-Castañeda y Miller (2004) |
| Charo | Michoacán | Plioceno temprano? | Blancano? | Alberdi y Corona (2005) |
| La Goleta | Michoacán | Plioceno temprano | Blancano | Miller y Carranza-Castañeda (1984) |
| El Huichol | Nayarit | Mioceno tardío | Henfiliano | Carranza-Castañeda (2006) |
| Huajapán | Oaxaca | Incierta | Incierto | Felix y Lenk (1891) |
| Tlaxiaco | Oaxaca | Incierta | Incierto | Felix y Lenk (1891) |
| Tecamachalco | Puebla | Incierta | Incierto | Felix y Lenk (1891) |
| Minas Prietas | Sonora | Incierta | Incierta | Frick (1933) |
| San José de Pimas | Sonora | Mioceno tardío-Plioceno temprano | Henfiliano- Blancano | Lindsay (1984) |
| Localidad incierta | Tlaxcala | Plioceno tardío | Blancano | Osborn (1921), Miller y Carranza-Castañeda (1984) |
| Yerbaniz, Jalpan | Zacatecas | Plioceno temprano | Blancano | Carranza-Castañeda (2006) |
| Gomphotherium | ||||
| Formación Ixtapa, Ixtapa | Chiapas | Mioceno medio | Barstoviano | Ferrusquía-Villafranca (1990a) |
| Landa de Matamoros | Querétaro | Mioceno tardío | Henfiliano | Carranza-Castañeda y Miller (2004) |
| Formación El Camarón, Nejapa | Oaxaca | Mioceno medio | Barstoviano | Ferrusquía-Villafranca (1990b); Ferrusquía-Villafranca (2003) |
| El Gramal | Oaxaca | Mioceno medio | Barstoviano | Ferrusquía-Villafranca (1990b) |
Cuadro 2 Medidas expresadas en mm de molares bilofodontes deciduos y permanentes de gonfoterios longirrostrinos y brevirrostrinos que se utilizaron para los análisis bivariantes.
| Posición | AP. | Tr. | Taxón | Localidad | Autor |
|---|---|---|---|---|---|
| p4 | 44,65 | 40,66 | Gomphotherium sp. | Puente Ixcán, Chiapas | |
| p4 | 46 | 40 | Gomphotherium hondurensis | Guatemala | Lucas y Alvarado (1995) |
| p4 | 48 | 44 | Gomphoterium angustidens | Somosagua, España | Mazo (2000) |
| p4 | 40 | 34 | Ghomphoterium angustidens | Hidroeléctrica, España | Mazo (2000) |
| p4 | 47 | 43 | Ghomphoterium angustidens | Puente de Toledo, España | Mazo (2000) |
| p4 | 41 | 39 | Ghomphoterium angustidens | Yuncos, España | Mazo (2000) |
| p4 | 48 | 44 | Ghomphoterium angustidens | Paracuellos 2, España | Mazo (2000) |
| p4 | 41,5 | 35,5 | Gomphotherium subtapiroideum | Sandelzhausen, Alemania | Göhlich (2010) |
| p4 | 46 | 42 | Gomphotherium subtapiroideum | Sandelzhausen, Alemania | Göhlich (2010) |
| p4 | 48 | 43 | Gomphotherium subtapiroideum | Sandelzhausen, Alemania | Göhlich (2010) |
| p4 | 40,5 | 37,16 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 41,5 | 37,9 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 41,14 | 35,48 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 49 | 41 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 47 | 40 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 39,5 | 29,5 | Ghomphoterium angustidens | Sandelzhausen, Alemania | Göhlich (2010) |
| p4 | 42 | 31 | Gomphotherium subtapiroideum | Sandelzhausen, Alemania | Göhlich (2010) |
| p4 | 40,5 | 30 | Gomphotherium subtapiroideum | Sandelzhausen, Alemania | Göhlich (2010) |
| p4 | 43 | 31 | Gomphotherium subtapiroideum | Sandelzhausen, Alemania | Göhlich (2010) |
| p4 | 40,8 | 34 | Gomphotherium subtapiroideum | Simorre, Francia (serie tipo) | Tassy y Göhlich (2011) |
| p4 | 42 | 31,16 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 48 | 33,98 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 47,12 | 34,74 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 47,94 | 34,92 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| p4 | 44,6 | 33 | Ghomphoterium angustidens | Péjouan, Gers, Francia | Tassy (2014) |
| dp2 | 31,84 | 25,52 | Gomphotherium hondurensis | Costa Rica | Valerio y Laurito (2008) |
| dp2 | 35,35 | 30,12 | Gomphotherium hondurensis | Costa Rica | Valerio y Laurito (2008) |
| dp2 | 25 | 21 | Rhynchotherium sp. | Arizona, EUA. | Miller (1990) |
| dp2 | 27,5 | 24 | Cuvieronius hyodon | Chapala, México | Aguilar (2011) |
| dp2 | 23 | 18 | Cuvieronius hyodon | Tarija, Bolivia | Boule y Thovelin (1920) |
| dp2 | 33,5 | 30,5 | Stegomastodon waringi | Brasil | Alberdi et al. (2002) |
| dp2 | 34 | 31 | Stegomastodon waringi | Brasil | Alberdi et al. (2002) |
| dp2 | 34 | 31 | Stegomastodon waringi | Brasil | Alberdi et al. (2002) |
| dp2 | 32 | 28,5 | Stegomastodon waringi | Brasil | Alberdi et al. (2002) |
Las medidas se expresan en milímetros y se obtuvieron utilizando un calibrador digital con precisión de 0,1 mm. Las fotografías fueron realizadas con una cámara Cannon XS usando un objetivo 18-55 mm y posteriormente se procesaron en Adobe Photoshop. Los análisis bivariantes se obtuvieron mediante el programa estadístico JMP 8.0, las gráficas resultantes se editaron en Corel Draw X7. Los términos morfológicos están basados en Corona y Alberdi (2006). La taxonomía es siguiendo a Shoshani y Tassy (2005).
Paleontología sistemática
Clase Mammalia Linnaeus, 1758
Orden Proboscidea Illiger,1811
Superfamilia Gomphotherioidea Hay, 1922
Familia Gomphotheriidae, Hay, 1922
Subfamilia Gomphotheriinae Hay, 1922 Género Gomphotherium, Burmeister, 1837 Gomphotherium sp.
Material. IHNFG-4888, cuarto premolar permanente superior derecho (P4); IHNFG-4895, fragmento apical de incisivo superior (IHNFG, Instituto de Historia Natural, Fósil Geográfico; acrónimo histórico usado actualmente en la colección paleontológica de la Secretaría de Medio Ambiente e Historia Natural de Chiapas, México).
Localidad y edad. Puente Ixcán, Maravilla Tenejapa, Chiapas, México. Hemphiliano, Mioceno tardío (Carbot-Chanona, 2011).
IHNFG-4888 es un cuarto premolar permanente de tipo bunodonte-bilofodonte, en buen estado de conservación. Su forma general es cuadrangular, con bordes redondeados que le dan al molar aspecto globoso. La parte anterior carece de esmalte. Su morfología oclusal es simple; presenta únicamente dos lofos de ápice romo. El primer lofo presenta mayor desgate en relación al segundo lofo, este desgaste es más notorio sobre la región de los pretritos. Los pretritos forman una figura sencilla no trebolada, debido a que no existen conos centrales. Las cúspides externas e internas son de forma piramidal con el ápice romo; estas no se dividen y se encuentran ligeramente separadas. El talón es incipiente y exhibe una marcada crenulación. El molar conserva un fragmento de raíz; ésta es robusta, no bifurcada y presenta una sola rama que se inclina ligeramente hacia el extremo posterior. Posee un cíngulo en los bordes labial y posterior. El diente es pequeño, anteroposteriormente mide 44,65 mm y transversalmente 40,66 mm. El esmalte tiene un grosor de 1,5 mm.
IHNFG-4895 corresponde a un fragmento apical de defensa superior, con una longitud de 59 mm y 32 mm en su parte más ancha. El extremo anterior de la defensa es triangular. La parte ventral es plana y conserva una banda de esmalte de 2 mm de grosor. El margen dorsal es rectilíneo. Debido a que se trata de un fragmento de la región apical no se observa indicio de torsión sobre su eje, pero si una ligera curvatura sobre la región esmaltada. En la cara lateral y medial se presentan estrías que corren longitudinalmente. En sección transversal forma una figura triangular ovalada, formada por capas concéntricas de dentina.
Discusión
Asignar dientes aislados a algún taxón dentro de Gomphotheriidae, especialmente en individuos juveniles, es una tarea compleja debido a la variedad de especies que existieron, ya sea en su forma brevirrostrina o longirrostrina (Lucas y Alvarado, 2010). Sin embargo, al comparar el ejemplar IHNFG-4888 con los premolares y molares de los gonfoterios brevirostrinos y longirostrinos presentes en América del Norte y Centroamérica, pudimos observar que comparte la característica bilofodonte únicamente con el segundo premolar deciduo (dP2) de Cuvieronius (Aguilar, 2011), pero IHNFG-4888 es casi el doble de tamaño. Las dimensiones de IHNFG-4888 son equiparables al dP3 del ejemplar de Cuvieronius juvenil IHNFG-4864, pero el dP3 en este género es trilofondonte y presenta una morfología más compleja que el presente en los premolares y molares de los gonfoterios primitivos Gomphotherium y Rhynchotherium. Göhlich (2010) menciona que los premolares superiores en gonfoterios longirrostrinos son de mayor tamaño y de forma cuadrangular, a diferencia de los inferiores que tienden a ser pequeños y piriformes. La morfología de IHNFG-4888 es cuadrangular como en los premolares superiores.

Fig. 2 IHNFG-4888: a) Vista oclusal P4; b) Vista lingual P4. IHNFG-4895 c) Fragmento apical de incisivo superior vista ventral; d) Vista lateral. Escala: 20mm.
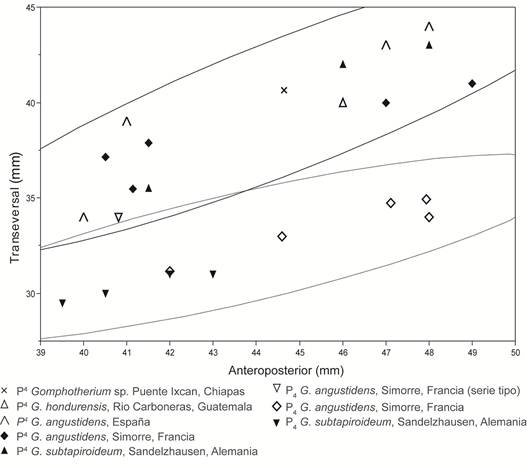
Por otra parte, las variables métricas permitieron agrupar a IHNFG-4888 con los P4 de gonfoterios longirrostrinos, separándose de los dP2 de los gonfoterios brevirrostrinos Cuvieronius y Stegomastodon (Fig. 3). Adicionalmente, el análisis bivariado que realizamos para discriminar los dientes inferiores de los superiores (Fig. 4), agrupa a IHNFG-4888 con los P4 de Gomphotherium. En consecuencia, basados en las características morfológicas y métricas de IHNFG-4888, determinamos que se trata de un cuarto premolar superior permanente de un gonfoterio longirrostrino.
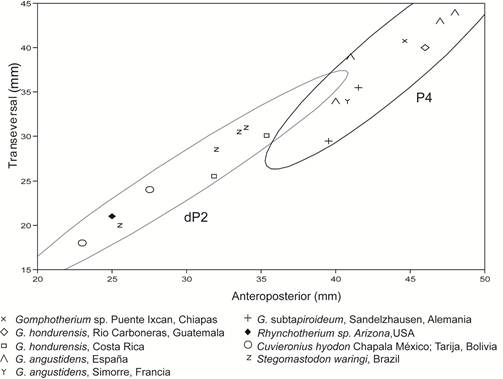
Fig. 3: Análisis bivariado en premolares bilofodontes de gonfoterios longirrostrinos y brevirrostrinos.
El reemplazamiento de molares en gonfoterios longirrostrinos y brevirrostrinos es diferente, incluso entre los gonfoterios primitivos (Fig. 5) (Ferreti, 2008b, 2010). En Ghomphotherium los premolares definitivos P3/3 y P4/4 sustituyen verticalmente a los deciduos dP3/3 y dP4/4, que a su vez son desplazados de forma horizontal por los primeros molares. Por lo tanto, la presencia del P3/3 y P4/4 es una característica morfológica que se presenta únicamente en las especies de Gomphotherium de América del Norte y Europa (Frick 1926; Tassy, 1990). Contrariamente, en Rhynchotherium los dP3/3 y dP4/4 son sustituidos por el M1/1 y M2/2, por lo que jamás presentan el P3/3 y P4/4, posiblemente porque poseía mandíbulas más angostas (Frick, 1926; Osborn, 1936).
Las características morfológicas y merísticas, permiten asignar el ejemplar IHNFG-4888 como Gomphotherium sp. La asignación específica no se realizó debido a lo escaso del material, aunque se asemeja en talla y morfología al P4 NMMNH P-14378 (Cuadro 1) del ejemplar de Guatemala asignado a G. hondurensis (Aguilar y Laurito, 2011), determinado primeramente como Rhynchotherium blicki por Lucas y Alvarado (1995). No obstante, el rango de tallas entre las especies de América y Europa no son significativas (Cuadro1; Fig. 3b).
Adicionalmente a las características morfológicas y métricas, usamos el rango geográfico conocido de Rhynchotherium y Gomphotherium como una variable más para sustentar nuestra asignación genérica, ya que Rhynchotherium se considera endémico para la región norte de América, siendo su límite geográfico conocido el norte de México (Aguilar y Laurito, 2011). Alberdi y Corona (2005) mencionan la presencia de Rhynchotherium para el centro de México, pero Lucas y Morgan (2008) consideran esto cuestionable, debido a que las determinaciones fueron basadas en material no diagnóstico. Por otro lado, material de América Central determinado originalmente como Rhynchotherium (e.g. Webb y Perrigo, 1984; Lucas y Alvarado, 1995; Laurito y Valerio, 2007; Valerio y Laurito, 2008), actualmente ha sido reasignado al género Gomphotherium (Lucas y Morgan, 2008; Lucas y Alvarado, 2010). Por su parte, Gomphotherium tiene un alcance geográfico más amplio y se conoce desde el norte de Estados Unidos hasta Panamá (MacFadden et al., 2015).
Se han propuesto ambientes semiacuáticos, sabanas arboladas y humedales como hábitat de Gomphotherium (Osborn, 1936; Webb y Perrigo, 1984; Laurito 2010; Arroyo-Cabrales et al., 2017). Carbot-Chanona (en prensa) con base en la presencia de tortugas, caimanes y cocodrilos en la localidad Puente Ixcán, infiere la existencia de afluentes de agua dulce cercanos a una zona costera y ambiente tropical para la zona, que corresponde con el ambiente de humedal como hábitat de Gomphotherium propuesto por Webb y Perrigo (1984).
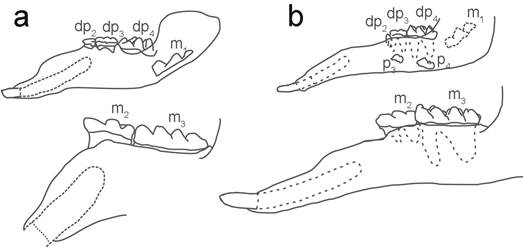
Fig. 5: a) Sustitución de molares en Rhynchotherium sp; b) Sustitución de dientes en Gomphotherium sp. Tomado y modificado de Frick, 1933
Conclusión
La morfología, medidas y alcance geográfico, permiten asignar los restos de gonfoterio rescatados en la localidad Puente Ixcán al género Gomphotherium; sin embargo, lo escaso del material no permitió hacer una determinación específica. Este género se documenta por primera vez para el Mioceno tardío de Chiapas. La presencia de Gomphotherium amplía notablemente su rango geográfico en México y viene a llenar el sesgo existente entre los registros del Mioceno tardío del centro de México y América Central.

