











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
Interest in coral restoration has increased in the last two decades due to the rapid global decline of coral reefs (Hoegh-Guldberg et al., 2007; Hughes et al., 2018). More recently, a special interest in the use of coral species or genotypes more tolerant to fluctuations in environmental conditions has emerged, as thermal anomalies associated with ENSO events are expected to increase in both frequency and intensity (Hughes et al., 2018; Rinkevich, 2014), reducing the time that these ecosystems have to recover from such disturbances (Boström-Einarsson et al., 2020). Therefore, it is important to identify those coral ecosystems that harbor scleractinian species capable of withstanding future disturbances from climate change and human activities, as they represent future coral refugia and may become sources of coral fragments for coral restoration programs. Identifying resilient communities requires ecological, physiological, and molecular studies to ensure that restoration strategies maintain and improve current restored areas and at least maintain coral diversity at ecological and genetic scales (Boström-Einarsoon et al., 2020; Montoya-Maya et al., 2016).
Within this context, it is important to prioritize resilient coral communities impacted by anthropogenic disturbances and climate change to evaluate their suitability to be restored and, eventually, rehabilitated. Along the Eastern Tropical Pacific (ETP), local coral communities have developed rapid acclimatization capacity and ability to remain stable over several years, despite being under the influence of moderate and severe El Niño Southern Oscillation (ENSO) events that have triggered bleaching, but no mortality in recent years (Cruz-García et al., 2020; Nava et al., 2021; Romero-Torres et al., 2020). Within the ETP, Punta de Mita (located in the Central Mexican Pacific, CMP) harbors a coral community that historically lost over 90 % of its live coral cover during the 1997-98 El Niño massive bleaching and mortality event (Carriquiry et al., 2001), which caused researchers to consider its recovery uncertain. However, recent life-history traits of corals in the area have promoted an increase in their ability to cope and resist different stressful conditions: not only has been observed thermal tolerance threshold but also a natural recovery capacity as well (Martínez-Castillo, Rodríguez-Troncoso, Mayfield et al., 2022; Tortolero-Langarica et al., 2017), which results in an opportunity to implement restoration strategies that contribute to the already observed recovery (Tortolero-Langarica et al., 2014; Tortolero-Langarica et al., 2017), implementing protocols that allow it to be site-specific since coral reefs do not thrive under the same conditions, especially under the current climate change scenario. Here we evaluate coral reef recovery resulting from active restoration at Punta de Mita, a site that has shown a natural recovery capacity from natural and anthropogenic disturbances.
Materials and methods
Study site: Punta de Mita (20.7699 °N, 105.5412° W) is located within the CMP (Fig. 1), a region considered an oceanographic transition zone where three water masses converge: the cold California Current flowing southernly, the warm coastal Mexican current flowing northerly, and the warm and saline water from the Gulf of California flowing southernly (Pantoja et al., 2012). The convergence of these currents creates a dynamic region, in addition to seasonal upwellings, nutrient pulses, and internal waves, wide daily temperature fluctuations (± 5 °C) can be recorded (Plata & Filonov, 2007; Portela et al., 2016). Punta de Mita presents a well-formed coral reef patch that used to harbor ~ 38 % of live coral cover, which was impacted by the 1997-98 ENSO event resulting in a dramatic loss of this ecosystem (Carriquiry & Reyes-Bonilla, 1997; Carriquiry et al., 2001).
Fig. 1 A. The Central Mexican Pacific (CMP), B. Punta de Mita, North of Banderas Bay within the CMP, C. Coral restoration site at Punta de Mita.
Coral restoration and monitoring: Coral restoration was performed in two different phases at a coral reef community that is at three meters depth: 1) the first in 2013 in order to validate the protocol, and 2) the second phase three years later in 2016. In each phase, a total of 50 Pocillopora fragments of opportunity, defined as fragments detached from an adult colony through natural processes such as water-motion, or bioerosion (Monty et al., 2006), were collected. Each coral fragment was individually examined to avoid signs of bleaching, damage, or algae overgrowth (such as turf and macroalgae) and then fixed to the natural substrate using zip ties within the same reef site (Fig. 2). All coral fragments restored during 2013 were tagged and measured for a two-year period (from November 2013 to October 2015), while those restored in 2016 were measured for one year (from January 2016 to February 2017) to record coral growth, i.e., the increase of fragment size (cm) maximum length (ML) and maximum width (MW) using a caliper (0.05 mm of precision). Also, survival of fragments was recorded.

Fig. 2 Pocillopora spp. fragment of opportunity fixed to the natural substrata in Punta de Mita, México A. initial fragment size during the first restoration period (2013), and B. Second restoration period (2016). Final colonies size at the end of the C. first restoration (2015) and D. second restoration period (2017).
Data analysis: Overall mean values of coral growth (± standard error) were calculated using Statistica 8.0 software (StatSoft Inc., 2007) and expressed as cm year-1. Coral growth rates, defined as the average total increase in coral length and width, were calculated using the original fragment size (cm ± standard error) for each restoration period. Also, the average and cumulative size increase in length and width (cm) within each period was estimated.
Simple linear regressions were calculated to evaluate the effect of changes in SST on coral growth rates. Mean monthly sea surface temperatures (SST) were obtained from the GIOVANNI database of the National Aeronautics and Space Administration of the United States (Li & Hegde, 2022). Regressions were calculated using Statistica 8.0 software (StatSoft Inc., 2007).
Results
The overall mean coral growth rate was 4.25 ± 0.15 cm year-1 for maximum length and 3.71 ± 0.17 cm year-1 for maximum width. Further details of each restoration period are given below.
First restoration period: Coral fragments tripled their size after 100 weeks of being reattached with an accumulated growth in length and width of ~ 8-9 cm (Fig. 3A). During the first restoration phase, corals grew at a rate of ~ 3.5 cm year-1 (Table 1). While coral growth remained stable throughout the first restoration period, the highest coral growth in both axes occurred during the warmest season (August 2015). The increase in coral size remained at 3.79 ± 0.21 and 3.55 ± 0.40 cm year-1 for both ML and MW (Table 1, Fig. 3B). Coral fragments self-attached to the substrata one month after being restored, and the final proportion of surviving corals was 38 % (Fig. 4).
Table 1 Coral growth rates in length and width (cm year-1) in each restoration period. ML: maximum length; MW: maximum width.
| Restoration period | ML | MW | ||||
| Mean | Min | Max | Mean | Min | Max | |
| First (2013) | 3.79 ± 0.21 | 1.97 | 5.38 | 3.55 ± 0.40 | 0.90 | 7.14 |
| Second (2016) | 4.49 ± 0.19 | 1.94 | 7.33 | 3.78 ± 0.16 | 1.63 | 6.17 |
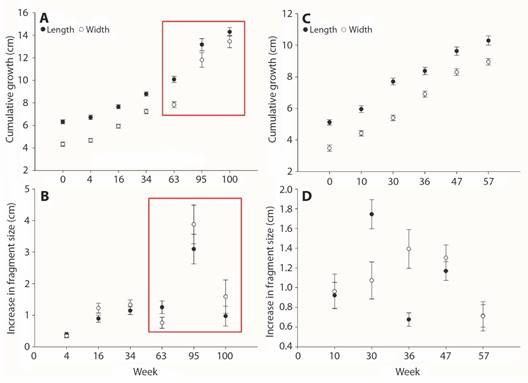
Fig. 3 A. Mean fragment size during the first restoration period (2013). B. Increase in coral size during the first period between measuring times. C. Mean fragment size during the second restoration period (2016). D. Increase in coral size during the second period between measuring times. Bars = SE. Red rectangles mark ENSO weeks during the first restoration phase.
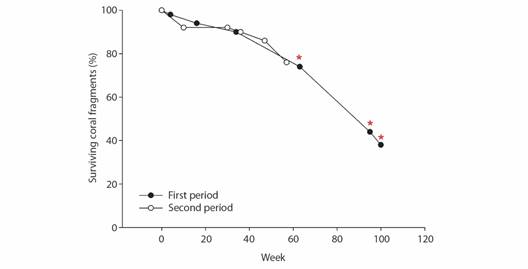
Fig. 4 Survival of restored Pocillopora spp. coral fragments during both restoration periods in Punta de Mita, México. Red asterisks signal ENSO weeks during the 2015-2016 El Niño event.
Second restoration period: Coral fragments almost doubled their size after 57 weeks following their re-attachment (Fig. 3C), with a maximum accumulated growth of ~ 5-6 cm in both length and width. The increase in coral size remained at 4.49 ± 0.19 and 3.78 ± 0.16 cm year-1 for both ML and MW (Table 1), peaking around week 30, during the warm season of 2016 (August; Fig. 3D). Corals self-attached to the substrate within the first two months after their restoration, and the proportion of surviving corals after 56 weeks was 76 % (Fig. 4).
Sea surface temperature and coral growth: During the first restoration period, the mean monthly sea surface temperature ranged from 25.7 to 32.3 °C, whereas during the second period, the mean monthly SST ranged from 24.1 to 31.3 °C, coinciding with an El Niño event, causing thermal anomalies ≥ 2.8 °C (Fig. 5). When assessing the effects of sea temperature on coral growth parameters, both regressions were not significant; hence, coral growth was not significantly related to sea surface temperature (R = 0.51, P = 0.10).

Fig. 5. Thermal anomalies (°C) caused by ENSO events across both restoration periods. The red frame signals positive thermal anomalies during the most recent and severe El Niño event. Thermal anomalies correspond to the Oceanic Niño Index (ONI) from the National Oceanic and Atmospheric Administration (NOAA, 2022).
Discussion
The capacity of any ecosystem to recover from environmental perturbations depends on the ability of its organisms to adapt to new conditions, which is determined by their physiological traits, including regulatory mechanisms, performance, and environmental tolerance (Cooke & Suski, 2008); specifically for coral ecosystems, these physiological traits include coral growth rates and reproductive strategies, their acclimatization capacity, and the type of endosymbiont they harbor, among others. The Central Mexican Pacific is a region considered sub-optimal for coral development due to its wide ranges in temperature, nutrient pulses, and elevated sedimentation rates (Glynn, 2017). It is also a region where in the last two decades, at least three severe thermal anomalies have affected coral communities such as Punta de Mita (Hughes et al., 2018). In spite of this, recent studies have documented that live coral cover in this region is slowly recovering (Martínez-Castillo, Rodríguez-Troncoso, Mayfield et al., 2022), evidencing the potential of this particular site to be rehabilitated using active restoration protocols.
Coral fragments in our study almost doubled their size one year following their re-attachment and increased three times their size after two years. Pocillopora has been characterized as a coral genus with one of the highest growth rates within the EP, a key physiological trait that has allowed this genus to remain as the most abundant species (Romero-Torres et al., 2020; Tortolero-Langarica et al., 2017). Mean extension rates of corals are ~ 3.5-4.5 cm y-1, which will not only increase the fragment length or width but in the development of the three-dimensional complexity that other coral colonies have exhibited elsewhere within the CMP (González-Pabón et al., 2021; Tortolero-Langarica et al., 2014; Tortolero-Langarica et al., 2017; Tortolero-Langarica et al., 2020). Indeed, corals not only continued to grow, but they also exhibited extension rates that are among the highest recorded in the Eastern Tropical Pacific (Martínez-Castillo, Rodríguez-Troncoso, Mayfield et al., 2022); therefore, corals within this region increase coral cover and carbonate production, contributing to the recovery of these reef sites (Tortolero-Langarica et al., 2019).
Restoration, in its broad sense, aims to assist the recovery of a damaged or degraded ecosystem, and particularly for coral reefs, their recovery relays on the increase of live coral cover both in its area and in its three-dimensional structure, which directly depends on sustained coral growth (Rinkevich, 2014). The fact that restored corals continued to grow even during high thermal anomalies caused by recent ENSO events (as seen in this study), brings up the relevance of their physiological traits such as the acclimatization capacity, which have allowed corals at Punta de Mita to recover and remain stable under stress events, highlighting the CMP as an important EP refuge, not only for corals, but also for the organisms associated with them (Martínez-Castillo, 2020; Martínez-Castillo, Rodríguez-Troncoso, Mayfield et al., 2022; Rodríguez-Troncoso et al., 2016; Tortolero-Langarica et al., 2017).
Coral growth is influenced by abiotic factors from which sea surface temperature is considered the most important, and the threshold is determined by the local and regional acclimatization capacity of these organisms (Manzello et al., 2010; Martínez-Castillo et al., 2020). More important high temperatures also trigger bleaching response and may cause mortalities as observed previously in the region (Carriquiry et al., 2001); however, there was no significant relationship between coral growth or survival and sea temperature even under the influence of the most severe ENSO event recorded to date. After almost 60 weeks from coral re-attachment, survival rates in both restoration periods were above ~ 60 %, which is considered as successful as most restoration projects report between 60-70 % of survival (Boström-Einarsson et al., 2020), and it was even higher than other remote locations of the CMP with low influence of human activity (Tortolero-Langarica et al., 2020). Unlike those areas, Punta de Mita coral community is a coastal site 200 m distant from the shore, where there are luxury touristic complexes and gulf camps, with intense marine traffic and no regulation of touristic activities. The high mortality observed at the end of the first restoration period then may be due to two interacting factors: first, even though coral growth was not compromised by the 2015-2016 El Niño warm temperatures, the coral community must have suffered individual negative effects during its first stage (which coincides with the end of the first restoration period); second anthropogenic activity in Punta de Mita may also have played a role in coral mortality as this site lacks an official protection status. In spite of this, the growth rates of the surviving coral colonies were sufficient enough to prevent the loss of coral cover during this period, reinforcing the idea that the CMP harbor resilient coral communities (Cruz-García et al., 2020; Martínez-Castillo et al., 2020; Martínez-Castillo, Rodríguez-Troncoso, Tortolero-Langarica et al., 2022; Rodríguez-Troncoso et al., 2016; Tortolero-Langarica et al., 2017; Tortolero-Langarica et al., 2022). In this sense, the coral community at Punta de Mita may represent an important coral refuge in the near future as corals'in the region exhibit a high acclimatization capacity, a key feature of corals in the region that promotes the recovery of CMP coral ecosystems (Martínez-Castillo et al., 2020; Martínez-Castillo, Rodríguez-Troncoso, Mayfield et al., 2022; Martínez-Castillo, Rodríguez-Troncoso, Tortolero-Langarica et al., 2022; Rodríguez-Troncoso et al., 2016; Tortolero-Langarica et al., 2017; this study).
While restoration efforts at Punta de Mita have been successful so far, there is an urgent need for management policies to ensure that full ecosystem recovery can be achieved. Punta de Mita is located within Bandera's Bay, in an area that is planned to continue high-end urban development (Merchand-Rojas, 2012). A major threat to coral reefs are changes in land use caused by urban growth (Burke et al., 2011; Carpenter et al., 2008). Therefore, if there is no adequate management and regulation of recreative aquatic activities in Punta de Mita, corals will be resisting more stressors than they can withstand, and therefore, local or official protection of this resilient site is urgent. The present study gives insight into the efforts of coral restoration activities in the Central Mexican Pacific. It highlights the importance of considering ecological, biological, and physiological processes when developing restoration strategies and emphasizes the importance of identifying coral communities with evidence of both resistance and resilience capacity. In this sense, Punta de Mita represents an important site, as both natural and restored fragments have contributed to increase and maintain coral cover in a region unlikely to recover from thermal anomalies (Carriquiry et al., 2001). Moreover, recent ENSO effects have not negatively affected fragment growth rates, suggesting that this location is essential for coral recovery and a conservation priority as corals are more likely to withstand future environmental perturbations than other Eastern Pacific locations due to their biological and physiological traits.
Ethical statement: the authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

