











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
Las regiones montañosas tropicales se caracterizan por su elevada biodiversidad (Rahbek et al., 2019), que incluye una riqueza importante de tipos de vegetación arbórea y no arbórea distribuidos a lo largo de su gradiente altitudinal. De éstos, destaca un conjunto de comunidades arbóreas muy heterogéneas que se agrupan bajo el nombre de bosque de niebla (bosque mesófilo de montaña, bosque de montaña baja y alta, con su equivalente en inglés tropical montane cloud forests). Estas comunidades se distribuyen en un amplio cinturón altitudinal, desde los 1 600 (-800) a los 3 000 m.s.n.m. (Bruijnzeel et al., 2010; Villaseñor, 2010), en donde las condiciones ambientales se caracterizan porque la humedad y precipitación son altas y la niebla es frecuente durante la mayor parte del año (Eller et al., 2020; Scatena et al., 2010). Estas condiciones ambientales favorecen la presencia de una elevada riqueza de plantas vasculares y no vasculares que puede ser comparable a la detectada en comunidades vegetales de tierras bajas, como los bosques tropicales lluviosos (Scatena et al., 2010). Pero se diferencia de éstas por una mayor presencia de plantas con hojas esclerófilas y la abundancia de epífitas que muy frecuentemente llegan a tapizar completamente los árboles (Eller et al., 2020).
Las comunidades de bosque de niebla son muy heterogéneas cuando los factores climáticos y topográficos cambian gradualmente y, en mayor medida cuanto éstos se presentan de manera abrupta. Estos cambios se observan principalmente con la temperatura y precipitación, incluido el gradiente altitudinal, con el que presentan una fuerte correlación, que favorecen diferencias en los atributos de la comunidad de plantas del bosque de niebla (Ramírez-Marcial et al., 2001; Williams-Linera et al., 2013). Por ejemplo, la diversidad alfa se incrementa hacia las partes intermedias del gradiente altitudinal (Martínez-Camilo et al., 2018; Williams-Linera et al., 2013) y la composición de especies cambia entre los sitios separados altitudinalmente (Williams-Linera et al., 2013).
La mayoría de los estudios en bosques de niebla en el neotrópico se han enfocado en evaluaciones a lo largo de gradientes altitudinales que favorecen muchos cambios ambientales (Domínguez-Yescas et al., 2020; Jiménez-Paz et al., 2021; Toledo-Garibaldi & Williams-Linera, 2014; Williams-Linera et al., 2013; Worthy et al., 2019). O en la influencia de ciertos rasgos topográficos, como la orientación de la ladera y la consecuente exposición a distintos niveles de radiación o periodicidad solares, que determinan cambios en la diversidad y composición florística (Yirdaw et al., 2015). Sin embargo, no se ha realizado hasta el momento una evaluación que considere un mismo piso altitudinal en una amplia extensión geográfica.
La ubicación de sitios de muestreo distribuidos en una extensa área con escaso contraste altitudinal permitiría captar la variación climática a nivel regional y no tanto por las diferencias altitudinales en un mismo transecto. Las diferencias altitudinales determinan cambios ambientales conforme se asciende en las montañas, por ejemplo, sobre la temperatura, radiación atmosférica y la precipitación (Körner, 2007). Estos cambios hacen que las especies presenten nichos ambientales muy específicos, en donde la capacidad de dispersión de las plantas se limita ya que pocas especies pueden tolerar amplios rangos ambientales. Al establecer sitios de estudio distribuidos a lo largo de un sistema montañoso, en una misma amplitud altitudinal, las variables ambientales serían más o menos homogéneas. Se esperaría entonces que las comunidades arbóreas también lo fueran, debido a que las especies tendrían mayor capacidad de desplazamiento o dispersión. Esta idea en parte es propuesta por Janzen (1967) que sugiere que la ausencia de una estacionalidad térmica en las regiones tropicales hace que los nichos climáticos de las especies sean más restringidos y limiten su movilidad a lo largo de gradientes térmicos.
La Sierra Madre de Chiapas es un sistema montañoso que se ubica en el sureste de México. Forma parte de la Cordillera Central, un extenso sistema montañoso centroamericano que inicia entre los límites de los estados mexicanos de Chiapas y Oaxaca, recorre Chiapas de manera paralela al Océano Pacífico, y luego atraviesa Guatemala, El Salvador y termina en Honduras. La Sierra Madre de Chiapas se extiende en casi 2.3 grados y aquí se encuentra uno de los bosques mejor conservados y continuos del sureste de México. Sin embargo, los escenarios para su conservación son inciertos. Navarrete et al. (2010) estimaron una pérdida de 50.5 % de estos bosques en un periodo de 26 años (1974 a 2000). Sin embargo, los procesos de deforestación y fragmentación son distintos, dependiendo del contexto social y económico a lo largo de la Sierra Madre de Chiapas, del gradiente altitudinal y de la gestión y conservación mediante las áreas naturales protegidas. Por ejemplo, Cortina-Villar et al. (2012) estimó una pérdida de cobertura boscosa del 12 % del área total de tres áreas naturales protegidas (Reservas de la Biosfera El Triunfo y La Sepultura, Área de Protección de Flora y Fauna La Frailescana).
Considerando la cobertura y conservación del bosque de niebla en la Sierra Madre de Chiapas, es posible realizar distintos puntos de muestreo para determinar el nivel de heterogeneidad de los bosques de niebla y evaluar el efecto de variables ambientales sobre sus atributos. Por lo que en este estudio se evaluó la variación del bosque de niebla en cinco sitios de la Sierra Madre de Chiapas. Los objetivos de este estudio fueron: (1) analizar los patrones de diversidad y estructura del bosque de niebla en la Sierra Madre de Chiapas y (2) determinar su diferenciación florística a lo largo de su área de distribución natural. Para determinar el nivel de influencia de variables climáticas sobre los atributos evaluados del bosque de niebla, se consideraron dos variables: la precipitación y la temperatura. Debido a que el muestreo se realizó en un cinturón altitudinal más o menos estrecho, la variación de la temperatura es menor en comparación a un gradiente de precipitación, con un incremento de la precipitación hacia el sureste.
Materiales y métodos
Área de estudio: La Sierra Madre de Chiapas, ubicada al sureste de México, es un sistema montañoso que transcurre paralelo al Océano Pacífico. Su longitud es de aproximadamente 280 km y tiene una anchura que varía de unos 50 km en el noroeste y de 65 km en el sureste (Müllerried, 1957). Su origen se debe a un surgimiento producto de la interacción de tres placas tectónicas (Cocos, norteamericana y la del Caribe) hace unos 16-10 Ma (Witt et al., 2012). El perfil geológico de la Sierra Madre de Chiapas es variado e incluye principalmente la presencia de roca metamórfica del Precámbrico, roca volcánica del Terciario Superior y Cuaternario, roca intrusiva del Mioceno Superior y roca intrusiva del Precámbrico y Paleozoico (Müllerried, 1957). La actividad volcánica es distinta en la región y únicamente una porción ubicada en el sureste (cerro Boquerón y el volcán Tacaná) ha tenido actividad en los últimos miles de años. El único volcán activo es el Tacaná. Las interacciones de las placas continentales, la actividad volcánica y sísmica han moldeado una configuración topográfica compleja y abrupta (Andreani & Gloaguen, 2016; Witt et al., 2012). El clima es variable y depende de la ubicación en el gradiente altitudinal: cálido subhúmedo y cálido húmedo en las porciones bajas e intermedias (0-1 500 m) y semicálido húmedo y templado húmedo en las partes medias (> 1 500) y altas (> 3 000), respectivamente. Se han descrito varios tipos de vegetación que incluyen principalmente el bosque tropical perennifolio, bosque tropical caducifolio, bosque de coníferas, bosque de encinos, bosque de niebla y matorral perennifolio. De estos tipos de vegetación, el bosque de niebla de la Sierra Madre de Chiapas es uno de los remanentes más importantes de Mesoamérica y México (González-Espinosa et al., 2011; Navarrete et al., 2010). En la región se ubica entre los 1 500 (-1 300) m y hasta los 3 000 m. Limita en su parte baja con comunidades de bosque tropical perennifolio, bosque tropical caducifolio o bosques de ecotono (zona mixta con elementos florísticos de las porciones altas y bajas).
Sitios de estudio: Se determinaron cinco sitios con bosque de niebla en la Sierra Madre de Chiapas (Fig. 1), de noroeste a sureste: (1) La Sepultura, las parcelas se ubicaron entre los 1 700-1 900 m.s.n.m., en los límites del ejido Sierra Morena y la zona núcleo de la Reserva de la Biosfera La Sepultura; (2) Pijijiapan, entre los 1 700 - 1 900 m.s.n.m., en la zona de amortiguamiento y zona núcleo V de la Reserva de la Biosfera El Triunfo; (3) El Triunfo, entre los 1 800 - 2 000 m.s.n.m., dentro de la zona núcleo 1 de la Reserva de la Biosfera El Triunfo; (4) Pico de Loro, entre los 1 800 - 2 000 m.s.n.m., en áreas que son del ejido San Juan Panamá y cercanas a la reserva estatal denominada Área de Protección Ecológica de Flora y Fauna Pico de Loro-Paxtal; (5) Tacaná: las parcelas se realizaron entre los 1 800 - 2 100 m.s.n.m., en los alrededores del ejido Chiquihuite y dentro de la zona de amortiguamiento de la Reserva de la Biosfera Volcán Tacaná. Las parcelas se ubicaron en un cinturón altitudinal más o menos constante entre sitios (~ 400 m). En la Tabla 1 se muestra un resumen de los parámetros climáticos y de altitud de cada uno los sitios. Aunque se privilegió la búsqueda de rodales forestales en buenas condiciones y sin aparente perturbación humana, el único sitio en donde no se pudo encontrar un bosque en tal condición fue en Pico de Loro, en donde se observaron evidencias de la frecuente extracción de leña y madera. En este caso, se anotaron y midieron todos los tocones en cada parcela.
Tabla 1 Atributos ambientales del bosque de niebla en cinco sitios de la Sierra Madre de Chiapas. / Table 1. Environmental attributes of the cloud forest at five sites of Chiapas' Sierra Madre.
| Sitio/variables | Altitud (m.s.n.m.) | TPA (°C) | PTA (mm) |
| La Sepultura | 1 772 ± 11.88 | 20.13 ± 0.21 | 1 848.63 ± 66.13 |
| Pijijiapan | 1 739.38 ± 58.27 | 19.71 ± 0.13 | 2 164.97 ± 8.0 |
| El Triunfo | 2 029.38 ± 61.52 | 17.21 ± 0.29 | 2 434.91 ± 109.38 |
| Pico de Loro | 1 892 ± 75.91 | 18.06 ± 0.53 | 2 173.59 ± 54.84 |
| Tacaná | 1 927.38 ± 93.3 | 17.29 ± 0.25 | 3 298.25 ± 43.48 |
Se muestran los promedios y la desviación estándar. Leyendas: TPA=Temperatura promedio anual, PTA=Precipitación total anual. / Averages and standard deviation are shown. Legend: TPA=Average annual temperature, PTA=Total annual precipitation.
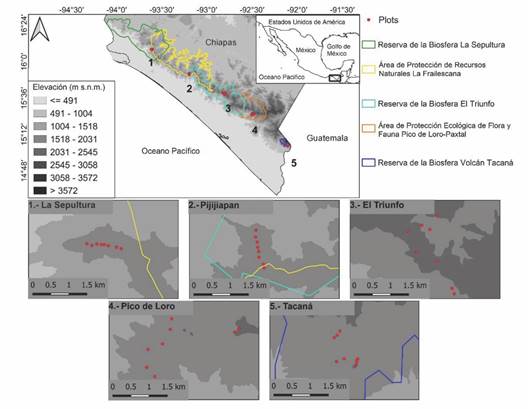
Fig. 1 Sitios y parcelas de estudio en el bosque de niebla en la Sierra Madre de Chiapas, México. / Fig. 1. Study sites and plots in Chiapas´ Sierra Madre cloud forest, Mexico.
Muestreo: En cada sitio se establecieron ocho parcelas circulares de 0.1 ha (40 parcelas en total). Se censaron y midieron todas las plantas leñosas con un diámetro a la altura del pecho (DAP, 1.3 m) ≥ 5 cm. En el centro de cada parcela se obtuvo una coordenada geográfica con un GPS marca Garmin Etrex Vista HCx. Los muestreos se realizaron del 2013 al 2016. Los ejemplares fueron determinados a nivel de especie en el campo, cuando no fue posible, se recolectaron muestras para su determinación con ayuda de claves taxonómicas y por comparación en el herbario Eizi Matuda de la Universidad de Ciencias y Artes de Chiapas.
Variables ambientales: Para cada parcela se obtuvieron los valores de temperatura promedio anual y de precipitación total anual. Estas se generaron con la base de datos Chelsa que comprende un periodo de 1980 a 2013 y una resolución espacial de ~ 1 km (Karger et al., 2017). Se utilizó esta base de datos ante la imposibilidad de obtener datos empíricos. Esta base de datos ha mostrado tener buen desempeño en sistemas montañosos en comparación a otras (Karger et al., 2017).
Análisis: Se calculó la diversidad alfa usando el enfoque de diversidad verdadera o los números de Hill (Chao et al., 2014). El cual considera la contribución relativa de las abundancias de las especies de acuerdo con las siguientes métricas (Chao et al., 2014; Jost, 2006; Jost, 2007): 0D, el número efectivo de especies o la riqueza observada de especies; 1D, las especies comunes, se pondera las especies en proporción a sus abundancias observadas y se obtiene con el exponencial del índice de Shannon; 2D, especies dominantes, asigna un mayor peso a las especies abundantes y se obtiene con el inverso del índice de Simpson.
Las variables estructurales de la comunidad que se evaluaron fueron: la densidad de individuos por unidad de área y el área basal obtenida para cada parcela. Los valores de ambas variables fueron extrapolados para su análisis por hectárea. Para analizar si hay diferencias estadísticas entre sitios en los parámetros de diversidad verdadera y de la estructura, se realizaron análisis de varianza (ANDEVA). Si las diferencias entre sitios fueron significativas se hicieron análisis post hoc con la prueba de Tukey. De manera complementaria, y a pesar de que las parcelas estuvieron ubicadas en un cinturón altitudinal más o menos estrecho, se realizaron modelos de regresión simple para determinar el nivel de relación entre las variables de respuesta (diversidad verdadera, área basal y densidad) con las variables climáticas (precipitación y temperatura) y el gradiente altitudinal. Para determinar los niveles de colinealidad entre las variables ambientales se realizaron análisis de correlación con la prueba de Pearson. En todos los modelos se verificó la correcta normalidad de los residuales.
Para evaluar los patrones de diversidad beta se utilizó una medida de disimilitud (índice de Bray-Curtis). Con este índice se obtuvo información que representan las diferencias en la composición de especies dentro y entre los sitios de estudio. Para analizar el efecto de las variables climáticas se utilizó un análisis de varianza permutacional utilizando matrices de distancia (PERMANOVA), por medio del uso de la función adonis del paquete vegan (Oksanen et al., 2019) en el programa R (R Core Team, 2021). En el PERMANOVA se usó el índice de Bray-Curtis como medida de distancia y 999 permutaciones. Con este análisis se incorporaron covariables o factores predictores, a partir del cual se parte la disimilitud entre las posibles fuentes de variación y usa una prueba de permutaciones para inspeccionar las fuentes de variación y ajustar un modelo lineal (Oksanen et al., 2019). Debido a la elevada colinealidad detectada entre la altitud y la temperatura promedio anual (r = -0.84, P < 0.001), y la temperatura promedio anual y precipitación total anual (r = -0.73, P < 0.001), y a la menor colinealidad entre altitud y precipitación total anual (r = 0.46, P < 0.05), en el modelo final del PERMANOVA las variables predictoras utilizadas fueron altitud, precipitación y la interacción entre éstas dos variables.
Para visualizar los patrones de composición y abundancia de la comunidad se realizó un análisis de Escalamiento Multidimensional no Métrico (NMDS), una técnica multivariante que representa en un espacio geométrico las afinidades entre las especies y las parcelas evaluadas (Oksanen et al., 2019). En el NMDS se utilizó la matriz de abundancia de las especies y el índice de Bray-Curtis como medida de distancia. Adicionalmente, en el gráfico se sobrepusieron los vectores calculados de las variables ambientales (altitud, precipitación y temperatura) para observar su correlación con la composición y abundancia, con la función envfit del paquete vegan (Oksanen et al., 2019) utilizando 1 000 permutaciones. En todos los análisis se usó un valor de significancia donde a = 0.05. Todos los análisis se realizaron con el programa R (R Core Team, 2021).
Resultados
Composición florística: Se censaron 4 021 individuos de árboles que corresponden a 220 especies, 114 géneros y 60 familias (Apéndice digital 1). Del total de especies, tres quedaron asignadas como morfoespecies por su dificultad para determinar la especie. Las familias con mayor riqueza de especies fueron: Rubiaceae con 22, Lauraceae (20) y Fagaceae (16). A nivel de género, destacan: Quercus con 16 especies, Ocotea (8) y Arachnothryx y Prunus (con siete especies cada una).
Diversidad alfa: El promedio de la riqueza de especies (0D) fue de 25.5 ± 6.9 (desviación estándar). Hay diferencias significativas en los promedios de riqueza entre los sitios (F4, 35 = 3.219, P < 0.05), aunque únicamente entre El Triunfo y Pico de Loro (P < 0.05; Fig. 2A). El promedio para 1D (exponencial del índice de Shannon) fue de 15.8 ± 5.0 y para 2D (inverso del índice de Simpson) de 11.0 ± 3.8 especies. En ambos parámetros no se encontraron diferencias estadísticas entre los sitios (1D: F4, 35 = 1.036, P = 0.403, Fig. 2B; 2D: F4, 35 = 0.483, P = 0.507, Fig. 2C). Los análisis con modelos de regresión simple indican una ligera relación entre 0D y el gradiente altitudinal, con una reducción de la riqueza hacia las partes más elevadas (Tabla 2; R2 = 0.12, P = 0.03). No se detectó relación de 0D, 1D y 2D con la temperatura y precipitación, y entre 1D y 2D con la altitud (P > 0.05, en todos los casos; Tabla 2).
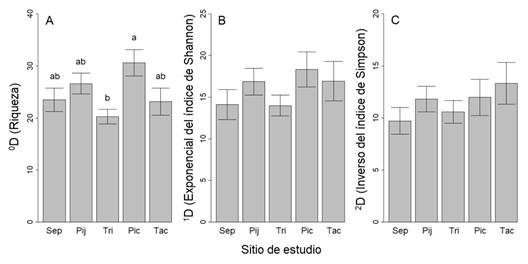
Fig. 2 Promedios y error estándar de tres parámetros de diversidad verdadera o números de Hill en cinco sitios de bosque de niebla en la Sierra Madre de Chiapas, México. La diversidad está representada por: A. 0D = riqueza observada, B. 1D = especies comunes, C. 2D = especies dominantes. Letras diferentes indican diferencias significativas de acuerdo con una prueba pareada de Tukey (P < 0.05). Leyendas: Sep = La Sepultura, Pij = Pijijiapan, Tri = El Triunfo, Pic = Pico de Loro, Tac = Tacaná. / Fig. 2. Averages and standard error of three parameters of true diversity or Hill numbers, at five sites of the Sierra Madre de Chiapas cloud forest, Mexico. Diversity is represented by A. 0D = observed richness, B. 1D = common species, C. 2D = dominant species. Different letters indicate significant differences, according to the Tukey pairwise test (P < 0.05). Legend: Sep = La Sepultura, Pij = Pijijiapan, Tri = El Triunfo, Pic = Pico de Loro, Tac = Tacaná.
Tabla 2 Resultados de los modelos de regresión simple para determinar si hay relación entre las variables ambientales y gradiente altitudinal sobre los parámetros de diversidad verdadera o números de Hill, área basal (m2 ha-1) y densidad de árboles (individuos ha-1). / Table 2. Results of the simple regression models to determine whether there is a relationship between environmental variables and elevational gradient on true diversity parameters or Hill numbers, basal area (m2 ha-1) and tree density (individuals ha-1).
| Variable de respuesta | Intercepto | Pendiente | R2 | P | |
| 0D | Altitud | 60.156647 | -0.018873 | 0.12 | 0.03* |
| Temperatura | 6.8561 | 0.972 | 0.03 | 0.27 | |
| Precipitación | 30.662980 | -0.002449 | 0.03 | 0.27 | |
| 1D | Altitud | 32.61628 | -0.00885 | 0.04 | 0.20 |
| Temperatura | 16.81773 | -0.04159 | 0.001 | 0.95 | |
| Precipitación | 14.404153 | 0.000690 | 0.004 | 0.68 | |
| 2D | Altitud | 19.798398 | -0.004439 | 0.02 | 0.43 |
| Temperatura | 18.2508 | -0.3660 | 0.01 | 0.5 | |
| Precipitación | 7.122529 | 0.001831 | 0.05 | 0.18 | |
| Densidad | Altitud | 2591.1060 | -0.8471 | 0.10 | 0.04* |
| Temperatura$ | 4.80513 | 0.11126 | 0.18 | 0.006* | |
| Precipitación$ | 7.738e+00 | -3.680e-04 | 0.32 | 0.0001* | |
| Área basal | Altitud | -65.10915 | 0.06438 | 0.15 | 0.01* |
| Temperatura | 126.805 | -3.864 | 0.06 | 0.14 | |
| Precipitación | 80.303863 | -0.010444 | 0.07 | 0.11 |
$Se aplicó un logaritmo natural a la variable de respuesta para normalizar los residuales. * indica diferencias significativas (P < 0.05). / $A natural logarithm was applied to the response variable to normalize the residuals. * indicates significant difference (P < 0.05).
Estructura: La densidad promedio fue de 1 005.25 individuos ha-1 (DE = 327.41). La parcela con la mayor densidad fue La Sepultura (1 770 individuos ha-1) y la de menor densidad El Triunfo (520 individuos ha-1). La densidad promedio de individuos fue diferente estadísticamente entre los sitios (F4, 35 = 5.009, P < 0.01, Fig. 3A). Las diferencias se detectaron entre los sitios Pijijiapan vs. Tacaná y Pico de Loro vs. Tacaná (P < 0.05, en ambos casos). Los modelos de regresión simple indican que la densidad de árboles se incrementa hacia las partes altas (R2 = 0.10, P = 0.04, Tabla 2) y con una reducción en la temperatura (R2 = 0.18, P < 0.01), y se incrementa también con una mayor precipitación (R2 = 0.32, P < 0.001).
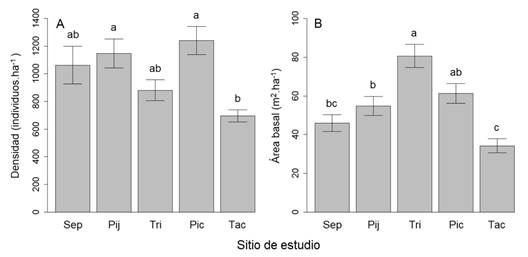
Fig. 3 Promedios y error estándar de las variables estructurales en cinco sitios de bosque de niebla en la Sierra Madre de Chiapas, México. A. Densidad de individuos. B. Área basal. Letras diferentes indican diferencias significativas de acuerdo con una prueba pareada de Tukey (P < 0.05). Leyendas: Sep = La Sepultura, Pij = Pijijiapan, Tri = El Triunfo, Pic = Pico de Loro, Tac = Tacaná. / Fig. 3. Averages and standard error of the structural variables at five sites of Chiapas´ Sierra Madre cloud forest, Mexico. A. Density of individuals. B. Basal area. Different letters indicate significant differences, according to the Tukey pairwise test (P < 0.05). Legend: Sep = La Sepultura, Pij = Pijijiapan, Tri = El Triunfo, Pic = Pico de Loro, Tac = Tacaná.
El sitio con la mayor cobertura de área basal fue El Triunfo, disminuyendo los valores hacia los sitios de ambos extremos geográficos (noroeste y sureste). El promedio de área basal fue de 55.4 m2 ha-1 (DE = 20.45), la parcela con el valor más elevado se localizó en El Triunfo (107.24 m2 ha-1) y la de menor área basal fue una de las de Tacaná con 52 m2 ha-1. El ANDEVA indica que hay diferencias en los promedios de área basal (F4, 35 = 12.76, P < 0.001). Las diferencias se detectaron entre los sitios La Sepultura vs. El Triunfo, Pijijiapan vs. El Triunfo, El Triunfo vs Tacaná, Pico de Loro vs. Tacaná (P < 0.05, en todos los casos, Fig. 3B). En los análisis de regresión, únicamente se detectó una leve relación positiva, pero significativa, entre la altitud y el área basal (R2 = 0.15, P = 0.01, Tabla 2).
Disimilitud y análisis multivariado: Los valores pareados de disimilitud indican una alta diferenciación en la composición de especies dentro y entre los sitios (Tabla 3). Las parcelas del Tacaná fueron las que tuvieron una mayor diferenciación en su composición. En el gráfico biplot de ordenación (NMDS) se observa que las parcelas se distinguen claramente por su composición entre los sitios, formando grupos bien definidos (Fig. 4). El análisis de PERMANOVA indica una relación entre la composición de especies con la altitud -temperatura-, precipitación y la interacción entre éstas (P < 0.01, en todos los casos, Tabla 4), estas en conjunto explican 24 % de la variación total. La precipitación presenta el valor de varianza ligeramente más alto (10 %) que la altitud.
Tabla 3 Comparación pareada de la diversidad beta (índice de Bray-Curtis) dentro y entre los sitios evaluados. Se muestran los valores promedios y la desviación estándar. / Table 3. Pairwise comparison of beta diversity (Bray-Curtis index) within and between evaluated sites. Average values and standard deviation are shown.
| Sitio | La Sepultura | Pijijiapan | El Triunfo | Pico de Loro | Tacaná |
| La Sepultura | 0.71 ± 0.13 | ||||
| Pijijiapan | 0.87 ± 0.07 | 0.68 ± 0.09 | |||
| El Triunfo | 0.92 ± 0.04 | 0.87 ± 0.05 | 0.69 ± 0.10 | ||
| Pico de Loro | 0.89 ± 0.07 | 0.84 ± 0.06 | 0.89 ± 0.05 | 0.71 ± 0.12 | |
| Tacaná | 0.96 ± 0.03 | 0.92 ± 0.05 | 0.94 ± 0.05 | 0.94 ± 0.05 | 0.78 ± 0.12 |
Los valores de referencia son: 0 = similitud total, 1 = diferencia total. / Reference values: 0 = total similarity, 1 = totally different.
Tabla 4 Análisis de varianza permutacional usando matrices de distancia (PERMANOVA) para evaluar el efecto de variables ambientales y de la altitud sobre la composición de especies de árboles en cinco sitios de la Sierra Madre de Chiapas. / Table 4. Permutation analysis of variance using distance matrixes (PERMANOVA) to evaluate the effect of environmental and elevation variables on the composition of tree species at five sites of the Sierra Madre de Chiapas.
| Variables | R2 | F | P |
| Precipitación | 0.10 | 4.8990 | 0.001*** |
| Altitud | 0.092 | 4.3782 | 0.001*** |
| Altitud * Precipitación | 0.05 | 2.3757 | 0.001*** |
| Total | 0.24 |
*Indica diferencias significativas: ***P < 0.001. / *Indicates significant differences: ***P < 0.001.
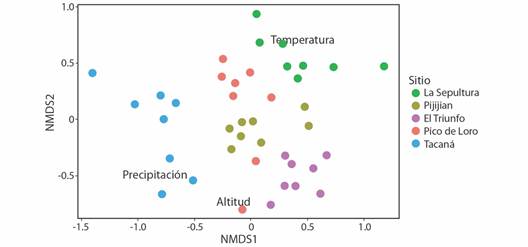
Fig. 4 Biplot de un Escalamiento Multidimensional no Métrico (NMDS) para observar las proximidades entre los sitios y parcelas, y sus relaciones con las variables ambientales evaluadas para la Sierra Madre de Chiapas, México. / Fig. 4. Biplot of Non-metric Multidimensional Scaling (NMDS) to observe proximities between sites and plots and their relationship to the evaluated environmental variables in the Sierra Madre de Chiapas, México.
Discusión
Patrones de diversidad alfa: En este estudio se evaluaron los patrones de variación en diversidad y estructura de especies arbóreas del bosque de niebla en aproximadamente 200 km en la Sierra Madre de Chiapas. Los valores de los parámetros de diversidad alfa son relativamente altos. Sin embargo, no es posible hacer una comparación detallada con otras regiones montañosas, principalmente de México y Centroamérica, por las discrepancias en los métodos de muestreo y procedimientos analíticos. Con esta premisa, la diversidad alfa a nivel de parcela en este estudio es similar a los valores detectados (0D = 20-30) en los bosques de la parte central de Veracruz, México (Toledo-Garibaldi & Williams-Linera, 2014; Williams-Linera et al., 2013), superior a los detectados en la Altiplanicie de Chiapas (con un promedio de 13.3 especies; Cayuela et al., 2006) y en el occidente de Jalisco (0D = 10-20 especies; Guerrero-Hernández et al., 2019), ambos en México; y menor a los encontrados en los Andes ecuatorianos, con una riqueza de 58 a 120 especies por 0.1 ha (considerando tallos > 2.5 cm DAP; Wilson & Rhemtulla, 2018).
Respecto a la variación en la diversidad alfa entre los sitios de estudio, con excepción del parámetro 0D (riqueza de especies), en 1D y 2D los valores fueron homogéneos, lo cual ha sido poco reportado en otras regiones montañosas con bosque de niebla, incluyendo estudios sin una variación altitudinal tan amplia (< 1 000 m) como en Brasil (Moreira et al., 2021) y Ecuador (Myster, 2017), aunque se ha encontrado esta tendencia similar en un estudio realizado en la región centro del estado de Veracruz, México, que incluye una variación altitudinal mayor a 1 000 m (Williams-Linera et al., 2013).
Sin embargo, cuando las comunidades de bosque de niebla se distribuyen en condiciones de poca variación ambiental, principalmente de factores asociados a la topografía y de variables ambientales como la temperatura local, parece haber poca variación en la diversidad alfa de árboles. Esto contrasta con la tendencia generalizada de cambios en la diversidad con relación al gradiente altitudinal y las variables climáticas (Körner, 2007; Rahbek, 1995, Rahbek, 2005), que promueven una mayor heterogeneidad del bosque de niebla (Moreira et al., 2021; Williams-Linera, 2002).
Patrones de estructura: Aún persisten controversias sobre el efecto de factores ambientales en los patrones estructurales del bosque de niebla. Los resultados de este estudio coinciden con otros en que la densidad de tallos disminuye con la altitud (Guerrero-Hernández et al., 2019), mientras que el área basal se incrementa (Alrutz et al., 2022; Guerrero-Hernández et al., 2019; Lieberman et al., 1996). Aunque también se encontrado otros patrones, como una tendencia unimodal de la densidad (Guerrero-Hernández et al., 2019) y la ausencia de cambios en los parámetros estructurales (Schwarzkopf et al., 2011). Esto sugiere respuestas particulares a factores ambientales, topográficos y, principalmente, de alteración natural y antrópico de cada sitio.
Las diferencias en la densidad entre los sitios, y en particular de aquellas que presentaron una mayor densidad, principalmente Pico de Loro, pueden deberse a los posibles efectos de disturbios naturales y humanos. Las parcelas en Pico de Loro fueron las únicas en donde se observó la presencia de extracción de madera (tocones) y de leña (ramas cortadas): en cinco parcelas se contabilizaron en total 25 tocones: promedio del diámetro = 27.6 cm, desviación estándar = 29.8. En este sitio, los remanentes de bosque son escasos y están próximos a áreas de cultivo de café y del poblado principal (ejido San Juan Panamá, municipio de Escuintla). Estas actividades de disturbio antropogénico son frecuentes y de magnitud moderada, que modifican la estructura de los bosques (Ramírez-Marcial et al., 2001). Principalmente, la reducción de árboles de diámetros grandes y la apertura de claros en el dosel que facilita la germinación y el crecimiento de especies arbóreas de rápido crecimiento, lo que a su vez incentiva la presencia de árboles de tallas medianas y pequeñas (Alrutz et al., 2022; Hölscher et al., 2010).
En Pijijiapan y La Sepultura también se encontró una alta densidad de árboles, aunque en estas zonas no se detectó evidencia de disturbio humano. Sin embargo, aquí predominan frecuentes e intensos vientos, principalmente entre los meses de noviembre a febrero (Briones-Salas et al., 2017; Velázquez-Muñoz et al., 2011), por lo que en estos bosques es frecuente la caída de árboles lo que conduce a una menor densidad de árboles gruesos y altos, y una mayor cantidad de árboles con diámetros medianos. En el estudio realizado por Myster (2021) en la Reserva Biológica San Francisco (Ecuador), aunque no precisa si éste fue en condiciones totalmente naturales, revela un incremento en la densidad de árboles del bosque de niebla de su distribución inferior altitudinal hacia la parte más elevada (de 1 900 a 2 800 m.s.n.m.), aunque tampoco se plantea si estos cambios están correlacionados con variables ambientales. Por lo tanto, no hay todavía una explicación clara de cómo la densidad se ve afectada por factores ambientales, pero los aspectos locales de alteración tanto natural y humana sí parecen ser importantes.
Las tendencias de área basal muestran un incremento hacia la parte central de la Sierra Madre de Chiapas, en El Triunfo. En este sitio se encontraron los árboles con los DAPs más grandes (> 140 cm); siete de los diez árboles más gruesos se encontraron en este sitio. Los demás sitios presentaron una reducción del área basal. Son pocos los estudios que han evaluado el área basal entre diferentes sitios con bosque de niebla. Los realizados en bosques conservados han encontrado en general patrones homogéneos, incluso en diferentes pisos altitudinales (Myster, 2021). Cuando los bosques están sujetos a perturbaciones se aprecia una reducción general en las coberturas de área basal (Alrutz et al., 2022; Hölscher et al., 2010; Ramírez-Marcial et al., 2001). Lo que sugiere que los rodales forestales evaluados en El Triunfo es uno de los sitios mejor conservados de la Sierra Madre de Chiapas.
Disimilitud y análisis multivariado: La diversidad beta (disimilitud) es marcadamente elevada dentro y entre los sitios a lo largo de la Sierra Madre de Chiapas. Se encontró que una separación de 40 km, considerando un mismo cinturón altitudinal, es suficiente para detectar un cambio drástico en la composición. Con una diferenciación en los ensambles de especies, casi de manera completa entre los sitios más alejados entre sí. Esta tendencia es similar a otros estudios realizados en otras regiones con bosques de niebla en el Neotropico (por ejemplo, Moreira et al., 2021; Williams-Linera, 2002) y particularmente similar a los resultados de Wilson & Rhemtulla (2018), quienes encontraron una alta diferenciación en la composición de especies considerando sitios de estudios ubicados en mismo cinturón altitudinal. Esta es una peculiaridad importante del bosque de niebla, que sugiere que la diversidad beta contribuye de manera importante a la diversidad regional (Jankowski et al., 2009). Cuando se incorpora un gradiente ambiental y climático, las diferencias suelen ser más fuertes en los cambios en la diversidad beta (Chain-Guadarrama et al., 2012).
La diferenciación florística observada en la Sierra Madre de Chiapas puede ser multicausal, incluyendo factores históricos (geológicos y evolutivos) y recientes (ecológicos). Sobre todo, al tratar de explicar la mayor separación entre las parcelas del Tacaná con las de los otros sitios, como se observa en la ordenación NMDS. Aunque la Sierra Madre de Chiapas tiene un mismo origen geológico, se pueden apreciar ciertos rasgos o características que pueden definir unidades geológicas más o menos diferentes: el sitio Tacaná es la única sub-región que ha tenido actividad volcánica reciente, lo que puede modificar las propiedades de los suelos, por ejemplo, las cenizas pueden enriquecer los suelos (Graham, 2010; Müllerried, 1957). Lo que promueve cambios sobre los conjuntos florísticos como se ha sugerido en otros estudios que describen la relevancia de la actividad volcánica y su influencia en la biodiversidad (Dossa et al., 2013; Montoya et al., 2021). La diferenciación florística también se debe a factores topográficos y climáticos (Hu et al., 2020; Rahbek et al., 2019). En los topográficos, principalmente el aislamiento ''insular'' de las montañas (''sky islands'') o efecto del aislamiento geográfico que puede dificultar la movilización de las especies, a pesar de encontrarse en una misma amplitud altitudinal (Ghalambor et al., 2006; Janzen, 1967). En los factores climáticos, se ha documentado un patrón muy notorio en la Sierra Madre de Chiapas, que implica la (1) disminución de la precipitación total anual de sureste a noroeste, y de (2) un incremento de la estacionalidad (mayor duración de la época seca y menor precipitación en ésta) de sureste a noroeste (Martínez-Camilo et al., 2018), lo que también explica la relevancia de las variables evaluadas en el análisis de PERMANOVA.
Conservación: Los bosques de niebla de la Sierra Madre de Chiapas son ecosistemas frágiles para su conservación debido a las amenazas por cambio de uso de suelo y otros disturbios humanos relacionados con la fragmentación de los bosques y los incendios forestales (Toledo-Aceves et al., 2011). Por lo que es importante establecer mecanismos que coadyuven a mantener la representatividad de esta comunidad vegetal a lo largo de la Sierra Madre de Chiapas. Esto es importante en términos de conservación, ya que, con un área total de muestreo en este estudio, de apenas 4 ha, se documentó 11 % de la riqueza arbórea estimada para el estado de Chiapas (González-Espinosa et al., 2004). Ningún sitio en particular puede servir para identificar sitios de mayor o menor diversidad arbórea en la Sierra Madre de Chiapas, ya que todos presentaron más o menos los mismos niveles. Así mismo, ningún sitio puede ser representativo del bosque de niebla, debido a su importante diferenciación en su composición florística. Por lo tanto, los esfuerzos de conservación deben ser fortalecidos y se debe de considerar en la medida de lo posible, la variación florística del bosque de niebla en esta región fisiográfica, incluyendo áreas que no están sujetas en algún esquema de conservación (por ejemplo, área natural protegida). Aunque buena parte de esta comunidad se encuentra en condiciones relativamente bien conservadas en la región, hay partes que están sujetas a escenarios críticos de amenaza debido a la deforestación, fragmentación y cambio de uso de suelo (Toledo-Aceves et al., 2011), como se ha observado en Pico de Loro.
Declaración de ética: los autores declaran que todos están de acuerdo con esta publicación y que han hecho aportes que justifican su autoría; que no hay conflicto de interés de ningún tipo; y que han cumplido con todos los requisitos y procedimientos éticos y legales pertinentes. Todas las fuentes de financiamiento se detallan plena y claramente en la sección de agradecimientos. El respectivo documento legal firmado se encuentra en los archivos de la revista.

