











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introduction
In recent decades, anthropogenic pressures (i.e., habitat destruction, climate change, chemical pollution, overharvesting, and land use change) on the natural environment and biodiversity have increased worldwide (Butchart et al., 2010). This has left a legacy of species range contractions and extinctions (Tilman et al., 2017; Torres-Romero et al., 2020). The world's mammalian large carnivores (Order: Carnivora) are disproportionately impacted by these pressures, and their remaining range is increasingly overlapping with human activities (López-Bao et al., 2017; Torres-Romero et al., 2020). Most large carnivore species have undergone extensive range contractions in recent human history and, are now listed as threatened with extinction (Ripple et al., 2014; Torres-Romero et al., 2020).
Variation in the ability of large carnivores to share the landscape with humans is important to both global and regional landscape conservation planning. Previous studies have demonstrated different evidence that illustrates coexistence scenarios between large carnivores and humans (Boron et al., 2016; Chapron et al., 2014; López-Bao et al., 2017). Land abandonment and/or conservation policies have facilitated coexistence between large carnivores and humans in different regions, such as Europe: i.e., the Eurasian lynx (Lynx lynx), brown bear (Ursus arctos) and wolf (Canis lupus) (Chapron et al., 2014; Cimatti et al., 2021). Sharing the landscapes with these carnivore species across many extensive human-dominated areas remains, however, one of the major barriers to large mammal's conservation (López-Bao et al., 2017; Lute et al., 2018).
Different conservation approaches have been proposed to protect large carnivores (Ripple et al., 2014). Some strategies have relied on the use of physical boundaries around protected areas, such as fences, to prevent conflicts and reduce threats (Packer et al., 2013); or have been focused on connecting core protected areas for these species using habitat corridors (Saura et al., 2019). Alternative strategies increasingly seek to incorporate the persistence of large carnivores into human-dominated landscapes (Chapron et al., 2014; Llaneza et al., 2018; Lute et al., 2018). This approach can sometimes lead to more frequent conflicts between people on how to conserve large carnivores, and consequently, greater local pressure threats on carnivores (i.e., retaliatory killings as retribution for livestock depredations, increased vulnerability to opportunistic but illegal hunting) (López-Bao et al., 2017). Regardless of the conservation strategy adopted, a lack of understanding regarding how large carnivores respond to these different conservation approaches (i.e., spatial use in relation to landscape protection) limits our ability to prioritize conservation planning efforts.
The jaguar (Panthera onca) is the largest felid in the Western hemisphere. It was historically distributed from the Southern United States, southward into Northern Argentine Patagonia (Sanderson et al., 2002). Over the past century, habitat destruction, poaching for illegal trade in body parts (i.e., skins, claws), depletion of the predator's prey base, and retaliatory killings following livestock depredations (Quigley et al., 2017), have contributed to the range contraction of the species, disappearing across more than half of its original geographic range (Sanderson et al., 2002). As land use intensifies and expands across Americas, this threat to jaguars will be compounded further by human population growth, illegal hunting, medicine markets, livestock production, fragmentation, road and other anthropogenic factors, all of which will only accelerate extinction risk (Cullen et al. 2016; Torres-Romero et al., 2020), and thus, effective policies are desperately needed to curb these threats. A review of suitable landscapes across Central and South America, highlighted that protected areas are too small and few to effectively protect jaguar populations (Rabinowitz & Zeller, 2010). Because jaguars range over relatively large territories (i.e., average home ranges of 128.6 ± 49.5 km2; González-Borrajo et al., 2017), they are increasingly overlapping with human activities in human biomes (Figel et al., 2019; Payan et al., 2013), which are biomes predominantly anthropogenic, the product of human population growth, land use and land cover change, croplands and other human influence on ecosystems (Ellis et al., 2010).
One conservation approach to ensure viable populations of this large felid conservation is land-sharing (Johansson et al., 2016). Although, protected areas can be critical to the viability of low density, wide-ranging species such as jaguars, private and communal lands with high-quality habitat and sustainable land use practices can be important to ensuring long-term population viability and connectivity (Sanderson et al., 2002). In Colombia, for example, recent studies have highlighted the importance of unprotected areas across human-modified landscapes to the conservation of jaguars and their prey (Boron et al., 2016; Payan et al., 2013). Similarly, private lands appear to be playing a significant role in the persistence of jaguars in the Gran Chaco near the Paraguay-Argentina border (Giordano et al., 2014; McBride & Thompson, 2018), south of which jaguars are considered ''functionally extinct'' (Quiroga et al., 2014). Understanding how jaguars use core habitat areas and mixed-use landscapes across the range of the species could be used to improve our understanding of which landscapes might benefit from more sustainable practices, as well as to identify key areas essential to jaguar persistence in human-dominated landscapes.
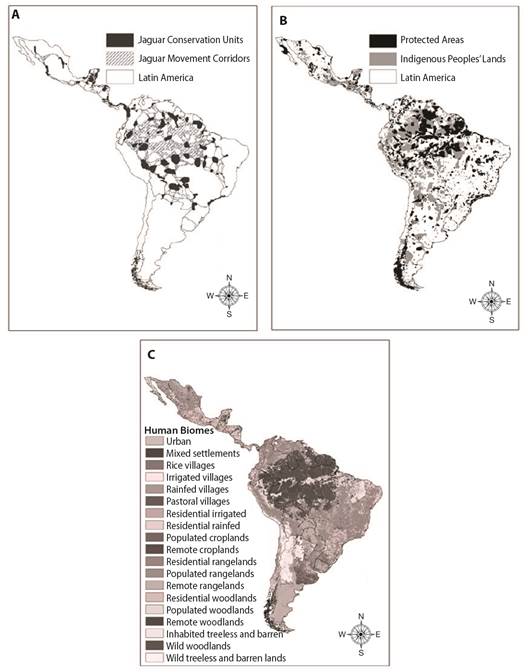
In this study, we evaluated the relative importance of different landscapes towards the extent of jaguar home-ranges across the Americas, including human biomes (Fig. 1). We did this by examining different spatial layers of land use relevant to jaguar conservation, including: (a) Jaguar Conservation Units; (b) Jaguar Movement Corridors; (c) Protected Areas; (d) Indigenous People's Lands, (e) a layer combination of potential core habitats as Jaguar Conservation Units, Protected Areas and Indigenous People's Lands, all layers pooled together, and (f) a layer related to landscape transformations caused by direct human interaction classified as anthropogenic biomes, also known as ''anthromes'' or ''human biomes'' (henceforth, human biomes; see., Ellis et al., 2010). Here, we analyze the home range of 117 jaguars to address the following questions: (1) What is the role of Protected Areas, Jaguar Conservation Units, Jaguar Movement Corridors, and Indigenous People's Lands in facilitating the movement of jaguars?; (2) What role do human biomes play in facilitating movement by jaguars?; (3) Can strategies that encompass land-sharing across human biomes also play an important role and if so, to what degree?
Materials and methods
Spatial and Landscape Data: We acquired the most recent database on jaguar movement from a database compiled by Morato et al. (2018). This database contains 134 690 spatial locations from 117 jaguars (54 males and 63 females) tracked using GPS technology. These individuals were monitored in five different countries: Brazil (N = 82), Paraguay (N = 23), Mexico (N = 8), Argentina (N = 3) and Costa Rica (N = 1), representing diverse parts of the jaguar's range. The majority of jaguars (N = 111) were adults (> 2 years old), four jaguars were juvenile, and two jaguars had undetermined age (see., Morato et al., 2018).
We used information on six layers conservation land protection categories considered important for jaguar conservation, including broad categories of human biomes. The six major spatial layers we considered included: (1) Jaguar Conservation Units (JCUs), defined by Sanderson et al. (2002) as ''core'' habitat areas of across the jaguar's range, representing ~2 million km2 and believed to have stable prey and capable of supporting at least 50 breeding jaguars; (2) Jaguar Movement Corridors (JMCs), which delineate potential routes of dispersal, immigration, and emigration among jaguar conservation units as described by Rabinowitz and Zeller (2010) (Fig. 1); (3) Protected Areas (PAs), which constitute ~7 million km2 of lands in Americas formally protected for their natural, ecological or cultural values as defined by the International Union for Conservation of Nature (UNEP-WCMC, 2020), and (4) Indigenous People's Lands (IPLs), which include terrestrial lands that are managed or co-managed by Indigenous Peoples (Garnett et al., 2018), and represented over ~4 million km2 across Latin America. Together, PAs and IPLs constitute most of the recognized terrestrial conservation land, habitat protection, and ecologically intact landscapes on Earth (Garnett et al., 2018) (Fig. 1). We used seven distinct PAs sub-categories, which differ in number and category of protection depending on the enabling laws of each country (see., International Union for Conservation of Nature, 2020 for further details of levels and regulatory protection), including: Ia-Strict Nature Reserves, Ib-Wilderness Areas, II-National Parks, III-Natural Monument or Features, IV-Habitat/Species Management Areas, V-Protected Landscape/Seascapes, and VI-Multiple Use Management & Protected Areas, whereby the sustainable use of natural resources is permitted.
We also examined (5) the combination of the three largest aforementioned conservation land layers together (i.e., JCUs, PAs and IPLs pooled; henceforth, ''Lyr-COMB''), as this overlap may indicate areas particularly important to regional biodiversity, or larger core habitat. Finally, we evaluated the effectiveness of (6) human biomes (HBs) layer, which constitute at least some measure of human modification, pressure, or footprint on the land via development. We examined this last category, which included different level and classes of development activities such as dense settlements, villages, croplands, rangeland, seminatural lands, and various other land uses (Fig. 1) (see., Ellis et al., 2010 for further details of classification and description).
Spatial data analyses and home range estimate: Because temporal autocorrelation or serial correlation in the radiotelemetry can underestimate the true home range size for each animal (Swihart & Slade, 1985), as well as bias home range size estimates due to different collection schedules, monitoring periods, and sampling frequencies among individuals and years, we randomly sampled one spatial location/day per individual (Börger et al., 2006). This helped to mitigate against spatio-temporal autocorrelation, resulting in a total dataset of 19 622 independent spatial locations for subsequent analyses. We then calculated annual home ranges for each jaguar, incorporating probabilistic estimators using kernel density estimation (KDE, via 95 % isopleth as a representative area; we refer hereafter to those results obtained using this metric) using the Home Range Tools module for ArcGIS (ESRI, 2012) a commonly used method to estimate an animal's home range (Laver & Kelly, 2008; Worton, 1989).
Next, we evaluated how each jaguar's home range overlapped with the different habitat protection categories and land uses, previously described. For each jaguar home-ranges (JHR) in our dataset, we calculate the percentage of the home range overlap where it occurred for each individual with every selected land category. In those cases where > 90 % of the KDE home range was occupied by a single landscape category (i.e., JCUs, JMCs, PAs, IPLs or Lyr-COMB) only that spatial category was considered. Then, when KDE jaguar home-ranges did not overlap with any formal conservation land or habitat protection categories, we identified the main human biomes (i.e., croplands, rangeland or seminatural lands) overlapping totally within each home range.
We then tested for differences in jaguar home-ranges between those located inside conservation lands, and the ones being outside or called ''unprotected'' areas. In order to determine if a significant difference in KDE home-range size exists between jaguars within any land categories, we built a generalized linear mixed model (GLMM) with gaussian distribution error and identity link to test differences in home range size in relation to the spatial overlap of jaguars with the different land conservation categories considered in this study. We also included the interaction terms between sex and land conservation categories. The number of locations was included as a covariate in the model to control for potential bias associated to different sampling efforts. The country was included as a random factor in the model (grouping factor, several jaguars sampled by country and mean home range size). We used the ''glmmAMDM'' package to run the model (Skaug et al., 2013).
Finally, we tested spatial variation in those cases where KDE home ranges overlapped in human-modified landscapes or human biomes (i.e., croplands, rangeland and seminatural lands); we compared differences of home-range size between females and males in relation to each human biome using linear mixed model (GLMM). All statistical analyses were performed in program R 3.4.0 (R Core Team, 2014) while spatial analyses, land use mapping and spatial comparative assessments were performed using ArcGIS (ESRI, 2012).
Results
Our results identified that 89 of 117 individuals (76 %; 49 females: 40 males) had a home range size ranging from 155 ± 19 km2 and, overlapped with the combination of land protection areas together (i.e., Lyr-COMB). Among these 89 jaguars, the size of home range of females and males ranged from 99 ± 14 to 224 ± 38 km2, respectively (Table 1). However, 28 of 117 jaguars (24 %; 13 females: 15 males) utilized ''unprotected'' lands, i.e., outside the boundaries of all land with some protected status combined. Of these jaguars (N = 28), the size of home ranges was generally larger though still varied considerably, with females and males ranging from 211 ± 59 to 762 ± 219 km2, respectively (Table 1). The size of home ranges varied depending on whether the home ranges were inside or not of the combination conservation lands (P < 0.05) (Table 1).
Table 1 Individual jaguars that occurred inside (in) and outside (out) of each land protection category
| JCUs | P | IPLs | P | PAs | P | JMCs | P | Lyr-COMB | P | |||||||||
| in | out | in | out | in | out | in | out | in | out | |||||||||
| females | females | females | females | females | ||||||||||||||
| JHR | 101 ± 18 | 157 ± 35 | > 0.05 | 157 ± 31 | 118 ± 19 | > 0.05 | 105±15 | 127 ± 22 | > 0.05 | 83 | 122±18 | 99±14 | 211±59 | <0.05 | ||||
| n | 38 (32.48 %) | 24 (20.51 %) | 7 (5.98 %) | 55 (47.01 %) | 13 (11.11 %) | 49 (41.88 %) | 1 (0.85 %) | 60 (51.28 %) | 49 (41.88 %) | 13 (11.11 %) | ||||||||
| males | males | males | males | males | ||||||||||||||
| JHR | 232 ± 41 | 614 ± 163 | > 0.05 | 384 ± 45 | 370 ± 76 | > 0.05 | 258 ± 57 | 403 ± 86 | > 0.05 | 473 ± 202 | 373 ± 70 | >0.05 | 224±38 | 762±219 | <0.05 | |||
| n | 35 (29.91 %) | 20 (17.09 %) | 5 (4.27 %) | 50 (42.73 %) | 12 (10.25 %) | 43 (36.75 %) | 2 (1.71 %) | 54 (46.15 %) | 40 (34.18) | 15 (12.82 %) | ||||||||
| both sexes | both sexes | both sexes | both sexes | both sexes | ||||||||||||||
| JHR | 164 ± 23 | ± 83 | > 0.05 | 251 ± 42 | 238 ± 39 | > 0.05 | 179 ± 32 | 256 ± 44 | > 0.05 | 343 ± 174 | 241 ± 36 | >0.05 | 155±19 | 507±123 | <0.05 | |||
| n | 73 (62.39 %) | 44 (37.60 %) | 12 (10.21 %) | 105 (89.74 %) | 25 (21.37 %) | 92 (78.63 %) | 3 (2.56 %) | 114 (97.43 %) | 89 (76.07 %) | 28 (23.93 %) | ||||||||
The number of individuals is indicated as n, and jaguar home-ranges are indicated as JHR. Mean home range sizes (km 2) for females, males and both sexes, and the results as mean ± standard error (SE) are shown. In each case (P) in bold represent the level statistically significant. Abbreviations are: Jaguar Conservation Units (JCUs), Indigenous People's Lands (IPLs), Protected Areas (PAs), Jaguar Movement Corridors (JMCs), and (Lyr-COMB) represent three layers categories combined (JCUs, IPLs and PAs). See main text for details on how jaguar home-ranges occurred inside and outside were depicted.
Seventy-three of 117 (62.4 %; 38 females: 35 males) JHR occurred in some part of JCUs, where females and males JHR ranged from 101 ± 18 to 232 ± 41 km2, respectively. While 44 of 117 (37.6 %; 24 females: 20 males) individual home ranges occurred outside the designated boundaries of JCUs, where the sizes of JHR ranging from 157 ± 35 to 614 ± 163 km2 for males and females, respectively, but jaguar home-ranges do not show significant differences (P > 0.05) between inside and outside of JCUs (Table 1).
We found fewer individuals (10 %; 7 females: 5 males) that overlapped with the IPLs, which presented home range areas ranging from 157 ± 31 to 384 ± 45 km2, respectively. While more individuals (N = 105) occurred outside of IPLs, we did not find significant differences (P > 0.05) in range size both inside and outside of IPLs (Table 1).
Twenty-five of 117 jaguar home-ranges (21 %; 13 females:12 males) occurred within PAs, where ranged from 105 ± 15 to 258 ± 57 km2, respectively. While 92 individuals (~79 %; 49 females: 43 males) were outside of PAs. The JHR both females and males increased their home ranges outside the boundaries of PAs, but we did not find significant differences (P > 0.05) both inside and outside of PAs (Table 1). On the other hand, only three individual jaguars (2 males: 1 female) (2.56 %) overlapped with potential JMCs; stated differently, the overwhelming majority of individual jaguar movements did not overlap at all with any part of a JMCs (97.43 %; 54 males: 60 females) and, in both overall, we did not find significant differences comparisons (Table 1).
Our results show that male and female jaguars in protected landscapes averaged smaller home ranges than male and female jaguars outside these conservation lands, respectively (Table 1). Thus, some individuals did not overlap their home ranges at all with some categories of conservation lands considered in this study. For example, we found that 28 jaguars (24 %; 15 males and 13 females) occurred in some human biomes lands such as rangelands (i.e., lands used mainly for livestock grazing and pasture), followed by croplands (i.e., lands used mainly for annual crops) and seminatural lands (i.e., inhabited rural lands, with minor use for permanent agriculture and settlements) (Table 2). Based on our dataset, male jaguars that occurred in human biomes lands showed the largest home ranges compared with females with significant differences in rangelands (females 249 km2 and males 685 km2; P = 0.015) and croplands (females 8 km2 and males 28 km2; P = 0.049), but non-significant was found in seminatural lands (females 7 km2 and males 21 km2; P = 0.699).
Table 2 Percent of contribution of each human biomes vs. 28 individual home range of jaguars
| Percent of contribution | |||||||||||
| Croplands | Rangeland | Seminatural lands | |||||||||
| Lands used mainly for annual crops | Lands used mainly for livestock grazing and pasture | Inhabited lands with minor use for permanent agriculture and settlements | |||||||||
| Sex | Country | # ind. | Residential rainfed croplands | Populated rainfed cropland | Remote croplands | Residential rangelands | Populated rangelands | Remote rangelands | Populated woodlands | Remote woodlands | |
| Female | Paraguay | 1 | 9.73 | 90.27 | |||||||
| Male | Paraguay | 2 | 0.5 | 99.5 | |||||||
| Female | Paraguay | 3 | 5.15 | 94.85 | |||||||
| Male | Paraguay | 4 | 100 | ||||||||
| Male | Paraguay | 5 | 12.36 | 87.64 | |||||||
| Female | Paraguay | 6 | 33.61 | 66.39 | |||||||
| Female | Paraguay | 7 | 100 | ||||||||
| Female | Paraguay | 8 | 32.07 | 67.93 | |||||||
| Male | Brazil | 9 | 77.54 | 14.04 | 8.42 | ||||||
| Male | Paraguay | 10 | 100 | ||||||||
| Male | Brazil | 11 | 79.29 | 20.71 | |||||||
| Female | Brazil | 12 | 39.22 | 60.78 | |||||||
| Female | Brazil | 13 | 21.63 | 19.23 | 59.13 | ||||||
| Female | Mexico | 14 | 4 | 96 | |||||||
| Male | Paraguay | 15 | 100 | ||||||||
| Male | Brazil | 16 | 6.8 | 48.54 | 44.66 | ||||||
| Male | Brazil | 17 | 10.37 | 23.89 | 5.55 | 15 | 33.34 | 11.85 | |||
| Male | Mexico | 18 | 44 | 56 | |||||||
| Female | Brazil | 19 | 81.25 | 18.75 | |||||||
| Female | Brazil | 20 | 8.24 | 88.84 | 2.92 | ||||||
| Male | Brazil | 21 | 77.78 | 22.22 | |||||||
| Male | Paraguay | 22 | 100 | ||||||||
| Female | Paraguay | 23 | 100 | ||||||||
| Female | Paraguay | 24 | 100 | ||||||||
| Male | Brazil | 25 | 77.25 | 22.75 | |||||||
| Female | Brazil | 26 | 100 | ||||||||
| Male | Brazil | 27 | 6.12 | 89.69 | 4.19 | ||||||
| Male | Brazil | 28 | 41.72 | 5 828 | |||||||
These 28 jaguars were outside the combination of the three lands with protected status (i.e., Jaguar Conservation Units, Protected Areas and Indigenous People's Lands pooled) can occur in more than one biome. Note: Human biomes were organized into groups according to Ellis et al. 2010 (see Ellis et al., 2010 for further details of classification).
Discussion
The results presented here correspond to the first continental-scale evaluation of how jaguar home ranges overlap with different land protection status, and anthropized systems, an important consideration given that previous jaguar studies have underscored the importance of protected and unprotected areas for the species (Boron et al., 2016; McBride & Thompson 2019; Payan et al., 2013; Thompson et al., 2021). Our results enlarge information in the sense of landscape conservation planning for species with large spatial requirements, which requires the political and financial commitment needed to implement ambitious local, regional and continental conservation and connectivity strategies (Keeley et al., 2019; McBride & Thompson 2019; Saura et al., 2019). Indeed, lands with some ''protected'' or conservation status have been shown to be important to jaguar populations and other large mammals (Pringle, 2017; Torres-Romero et al., 2020). Thus, PAs are arguably regarded as the most critical aspects of world conservation planning efforts, and they can be particularly critical to carnivore populations and other vertebrates (Di Minin & Toivonen, 2015; Wegmann et al., 2014).
On the other hand, indigenous lands are also crucial to the sustainability and conservation of wild animal and plant communities on Earth, occupying an even greater total area than existing protected areas (Fa et al., 2020; Schuster et al., 2019). Indigenous People's Lands are of course inclusive of land tenure rights for semi-autonomous and autonomous indigenous nations across ~38 million km2 in 87 countries; they further intersect with and/or are adjacent to about 40 % of all terrestrial protected land area, enhancing the ecological functioning and landscapes intactness (Garnett et al., 2018). The IPLs, despite representing less important areas to space use and movement for jaguars, still represents approximately 10.50 % of ecologically intact landscapes. Over the past several decades, however, anthropogenic pressure on PAs, IPLs and, otherwise, intact forest habitats have been accelerating (Qin et al., 2019). In 2019, for instance, bushfires devastated approximately 308 048 km2 of the Amazon Basin, nearly 70 % more than during the period of 2018; large portions of Brazil, Venezuela, Bolivia, and Colombia were affected (Lizundia-Loiola et al., 2020). Other ecoregions in South America, including the Gran Chaco and Pantanal, which include important JCUs for jaguars, similarly suffered devastating losses resulting from intentional but uncontrolled fires.
Rabinowitz and Zeller (2010) originally described 90 JCUs encompassing 1.9 million km2, and ~98 % of JCUs overlaps with other conservation lands. In this sense, our results show that JCUs play a leading role in the spatial patterns of space and movement of jaguars in a human-dominated landscape. This conservation lands have been proposed as critical jaguar conservation landscapes because of their perceived importance to long-term jaguar population viability and connectivity (Rabinowitz & Zeller, 2010; Zeller, 2007; Zeller et al., 2011). We should reiterate here that JCUs are not formal designations by any country and, while many encompass national protected and management areas, they may contain a mix of habitat quality and land use categories.
Similarly, Rabinowitz and Zeller (2010) identified 182 least-cost path corridors JMCs connecting the 90 JCUs across the jaguar's range. We do note, however, that based on our results, there is little evidence to suggest that JMCs are effectively being used by jaguars. This sort of evidence, however, requires special attention, because JMCs are not resident range, and it is therefore not surprising to found that jaguars do not show a space use and movement on these landscapes. As such, we recommend that future field investigations turn their attention to this critical knowledge gap, and to evaluate their use by jaguars. Otherwise, we face the risk of a proliferation of -paper corridors- that are never validated in the field.
On the other hand, our study highlights the relative importance of human biomes to jaguars. Our analyses show that throughout their geographic distribution, jaguars include within their home range human biomes such as rangeland, cropland, semi-natural lands and even villages' lands that do not necessarily have recognized conservation programs, but that could be important to spatial movements of endangered species. This shows that in addition to the decree of protected areas, it is essential to recognize and promote the proper management that various societies have over their territory, a strategy to do this is by promoting the establishment of voluntary conservation areas, even in partially anthropized systems (for example: areas voluntarily designated for conservation in Mexico, Gutiérrez-Hernández et al., 2021).
In the Paraguayan Chaco, for instance, evidence of the presence of jaguars was encountered regularly in unprotected and/or privately-owned mixed forest, semi-natural, and rangeland landscapes; however, at greater distances from protected areas, their presence was detected less frequently than pumas (Puma concolor), a consideration that has different conservation planning implications for each species (Giordano, 2015). In parts of Colombia, where hunting of primary prey species and retaliation killings were limited, jaguars were able to use human-dominated agricultural landscapes (Boron et al., 2016; Payan et al., 2013). A previous study of female cheetahs (Acinonyx jubatus) showed they tended to prefer denser vegetation types than males; they thus seek to mitigate exposure to human pressure through greater protective cover (Broomhall et al., 2003). Consistent with our results, we found that jaguars might be present in different human-dominated land-use types more broadly such as croplands, rangeland and seminatural-lands (Table 2). This finding is congruent with growing evidence that jaguars occur in sites with higher prey biomass because there would be sufficient food, which would increase the human-carnivore conflict (McBride & Thompson, 2018) due to depredations on livestock and pets, especially when natural prey and habitat is lacking (Athreya et al., 2016; Majgaonkar et al., 2019).
Therefore, effective continental, regional and local jaguar conservation planning may integrate the different types of human biomes used by the species (either to favor dispersal and connectivity between protected areas, or to support the permanent presence of jaguars) in the human-dominated matrix (Llaneza et al., 2018), together with conservation lands, in order to achieve an effective landscape-scale conservation approach for the species.
Because of the origin of the dataset, it is not surprising to find that the ranges of the collared animals overlap with the categories of protected lands, but the use of anthropized systems by the jaguar highlights the opportunity to promote the participation of different types of land management at the local, regional, and continental level for the conservation of this species. It is also important to recognize the different conservation schemes in the territory where landowners carry out surveillance activities, even in places with anthropized environments; cooperation between different levels of land management and conservation is essential to facilitate jaguar mobility through the JMCs maximizing the effective protective potential of the JCUs.
Therefore, the dataset from which we carry out the analyses presented in this work come from research works that did not follow the same protocols, or they did not have the same telemetry equipment; it also evidently comes from places where the population characteristics of the jaguar allowed its capture and subsequent monitoring, often close to areas expressly designated for the conservation of biodiversity (Giordano, 2015; Thompson et al., 2021). In this sense, we recognize that the data cannot be used to make comparisons among sites or countries, which is not our aim, and that it would be necessary to carry out a monitoring effort on spatial analysis of jaguar home-ranges in areas where there are no previous efforts to monitor or conserve the species.
Finally, this continental network of PAs, IPLs, and JCUs, as well as the incorporation of HBs connected by JMCs, might theoretically facilitate the movement of jaguars, while simultaneously protecting other endangered species, and enhancing the functioning of extensive ecological communities or entire ecosystems. We further note that some core areas with respect to JCUs and JMCs may have changed with respect to land use change and human impact since Sanderson et al. (2002) and, Rabinowitz and Zeller (2010). Furthermore, long-term viability of jaguars across their range necessitates a strategy of developing functional connectivity among key areas for the species, on one hand, and reassesses JCUs and JMCs on the other. Our work here therefore must be continued by others and improved upon, so that we might better understand how jaguars move across fragmented and human-dominated landscapes at regional and continental scales.
Ethical statement: the authors declare that they all agree with this publication and made significant contributions; that there is no conflict of interest of any kind; and that we followed all pertinent ethical and legal procedures and requirements. All financial sources are fully and clearly stated in the acknowledgements section. A signed document has been filed in the journal archives.

